6染色体与连锁群
- 格式:ppt
- 大小:1.45 MB
- 文档页数:87

第五章染色体和连锁群本章重点一、连锁遗传:二对性状杂交有四种表现型,亲型多、重组型少;杂种产生配子数不等,亲型相等、重组型相等。
二、连锁和交换机理:粗线期交换、双线期交叉,非姐妹染色体交换。
三、交换值及其测定:重组配子数/总配子数;测交法测定,也可用F2 材料进行估计。
四、基因定位和连锁遗传图:确定位置、距离,基因位于染色体上;二点测验、三点测验;连锁群、连锁遗传图。
学时:91900年孟德尔遗传规律重新发现以后,生物界广泛重视,进行了大量试验。
其中有些属于两对性状的遗传结果不符合独立分配规律→摩尔根以果蝇为材料进行深入细致研究→提出连锁遗传规律→创立基因论→认为基因成直线排列在染色体上,进一步发展为细胞遗传学。
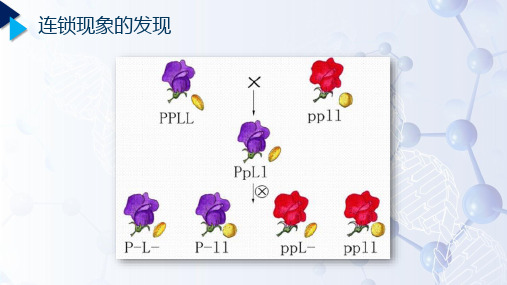
第一节连锁和交换一、连锁(一)连锁现象的发现1906年,贝特生(Bateson W.)和贝拉特(Punnett R. C.)在香豌豆的二对性状杂交试验中→首先发现性状连锁遗传现象。
第一个试验:P 紫花、长花粉粒(PPLL)×红花、圆花粉粒(ppll)↓F1紫、长PpLl↓⊗F2紫、长紫、圆红、长红、圆P_L_P_ll ppL_ppll总数实际个体数4831390393 13386952按9:3:3:1推算3910.51303.5 1303.5434.56952上述结果进行X2检验时,X2=3371.58,说明实计数与预计数差异极其显著,不可能由随机误差造成,应作重复实验。
以上结果表明F2 :①. 同样出现四种表现型;②. 不符合9:3:3:1;③. 亲本组合数偏多,重新组合数偏少(与理论数相比)。
第二个试验:P紫花、圆花粉粒(PPll)×红花、长花粉粒(ppLL)↓F1 紫、长PpLl↓⊗F2 紫、长紫、圆红、长红、圆P_L_P_ll ppL_ppll总数实际个体数22695971419按9:3:3:1推算235.878.578.526.2419结果与第一个试验情况相同。
连锁现象的发现一、孟德尔定律的实质图6-1 分离规律的实质图6-2 自由组合规律的实质二、人类和黑腹果蝇的染色体数与基因数人类(Homo sapiens )染色体(男性)(2n=46,约3.5万个基因)黑腹果蝇(Drosophila melanogaster )(2n=8,约1.36万个基因)三、连锁基因与连锁群位于同一条染色体上的基因,称为连锁基因(linkage gene )同一染色体上的所有基因,构成了1个连锁群(linkage group)1连锁现象的发现3基因在染色体上排列2连锁基因的遗传规律连锁基因连锁群4基因定位与染色体作图四、连锁现象的发现1906年彼特逊(Bateson )和庞尼特(Punnett)最先发现的。
香豌豆(Lathyrus odoratus )紫花长花粉粒红花圆花粉粒花色花粉粒形状◆F2出现4种表型,单个性状的遗传均符合3:1的分离比;◆两个性状的遗传不符合自由组合9:3:3:1的比例。
亲组合与两亲本性状组合(紫圆和红长)相同的亲组合类型实际观测数高于理论数;重组合两种性状重新组合出现的(紫长和红圆)重组合类型的观测数少于理论数;◆两性状的遗传不符合9:3:3:1的自由组合比例;◆单个性状的遗传均符合3:1的分离比;◆重组合(紫花圆花粉和红花长花粉粒)实际观测数少于理论数;重组合◆亲组合(紫花长花粉和红花圆花粉)的实际观测数高于理论数;亲组合七、两性状遗传是否符合自由组合规律的检验⚫卡方适合度检验统计推断:df=k-1=3时,χ2 0.01=11.35,χ2>>χ2 0.01,p < < 0.01说明两性状的遗传不符合9:3:3:1的自由组合(p < 0.01)。
∑−==EE O 221-k df )(χ=3371.58八、彼特逊和庞尼特的解释两对基因在杂交子代中并不是随机的自由组合。
☐相引(Coupling):原来属于同一亲本的两个基因更倾向于进入同一配子中;☐互斥(repulsion):原来属于不同亲本的两个基因之间在形成配子时相互排斥;相引相互斥相。


第六章染色体与连锁群我们已知基因在染色体上,它的分离,重组与染色体的行为相平行。
但是一个生物有很多基因,常以千万计,而染色体数目比较少。
如;玉米:n=10,400多个基因;果蝇:n=4,500多个基因;人类:n=23,5(10)万左右基因。
基因的数目远远的超过了染色体的数目。
这又怎么来说明呢?在一条染色体上一定有许多基因,它们又是怎样遗传的呢?这些基因叫连锁基因,连锁基因往往伴同而遗传。
这种遗传现象叫连锁遗传。
但由于形成配子时,同源染色体在某些区段可以发生交换,而使原来在同一条染色体上的基因不再伴同遗传,此现象称为交换。
第一节连锁与交换一、连锁遗传的发现1906年英国学者贝特森(W. Bateson)和潘耐特(R.C. Punnett)研究香豌豆两对性状的遗传。
紫花长花粉(PPLL)×红花圆花粉(ppll)和紫花圆花粉(PPll)×红花长花粉(ppLL)。
F1自交的结果图示。
发现:(1) F2不符合孟德尔9∶3∶3∶1之比。
而是7∶1∶1∶7(测交)。
(2) 原来同一亲本的两个性状,在F2中常常有连系有一起的倾向。
贝特森(W. Bateson)和潘耐特(R.C. Punnett)同时又将性状重新组合:紫花圆花粉(PPll)×红花长花粉(ppLL)↓紫花长花粉↓(自交)紫花长花粉(226)紫花圆花粉(95)红花长花粉(97)红花圆花粉(1)按9:3:3:1理论值 235.8 78.5 78.5 26.2进行卡平方测验=32.40 n=3 P< 0.01 差异极其显著也发现:(1) F2不符合孟德尔9∶3∶3∶1之比。
(2) 原来同一亲本的两个性状,在F2中常常有连系有一起的倾向。
对于上述实验结果,贝特森,潘耐特并没有提出科学的解释。
1910,美国学者Morgan, T.H. Bridges C.B.以果蝇为材料(白眼性状等基因的伴性遗传)。
同时研究了两对伴性性状的连锁遗传,指出两个基因存在于同一条染色体上,因而在形成配子时,这两个基因常常连在一起。

第六章染色体和连锁群遗传的染色体学说建立以后,进一步就要了解染色体与基因的关系。
但一个生物有很多基因,而染色体数目比较少,这又怎样来说明呢?正象Bridges研究果蝇眼色的遗传,发现了例外个体,使他注意到了新的现象一样,两对基因的杂交试验中,子二代分离比数与预期的9∶3∶3∶1有非常显著的差异,使遗传学工作者注意到了连锁现象。
这样不仅证明了染色体带有很多基因,而且证明了这些基因在染色体上是以直线方式排列的。
第一节连锁与交换一、连锁(linkage)连锁现象是Bateson和Punnett(1906年)最初发现的。
他们研究香豌豆的两对性状的遗传,发现同一亲体来的基因较多地联在一起,这就是所谓基因的连锁(linkage),但是他们未能提出正确的解释。
摩尔根(1910年)发现白眼性状的伴性遗传后,同年又发现几个伴性遗传的性状;他同时研究了两对伴性性状的遗传,知道凡是伴性遗传的基因,相互之间是连锁的。
这就证实了同一染色体上的基因有连锁现象。
1.香豌豆(Lathyrus odoratus)两对相对性状杂交试验现在请同学们一起来看一看香豌豆的这两个试验:花色:紫花(P)对红花(p)为显性;花粉粒形状:长花粉粒(L)对圆花粉粒(l)为显性。
①紫花、长花粉粒×红花、圆花粉粒(图6-1)图6-1 香豌豆紫花、长花粉粒×红花、圆花粉粒遗传分析A.F1两对相对性状均表现为显性,F2出现四种表现型;B.F2四种表现型个体数的比例与9:3:3:1相差很大,并且两亲本性状组合类型(紫长和红圆)的实际数高于理论数,而两种新性状组合类型(紫圆和红长)的实际数少于理论数。
②紫花、圆花粉粒×红花、长花粉粒(图6-2)图6-2 香豌豆紫花、圆花粉粒×红花、长花粉粒遗传分析A.F1两对相对性状均表现为显性,F2出现四种表现型;B.F2四种表现型个体数的比例与9:3:3:1相差很大,并且两亲本性状组合类型(紫圆和红长)的实际数高于理论数,而两种新性状组合类型(紫长和红圆)的实际数少于理论数。
