【遗传学】第五章 染色体和连锁群
- 格式:doc
- 大小:476.50 KB
- 文档页数:18


第五章连锁遗传和性连锁(一) 名词解释:1.交换:指同源染色体的非姊妹染色单体之间的对应片段的交换,从而引起相应基因间的交换与重组。
2.交换值(重组率):指同源染色体的非姊妹染色单体间有关基因的染色体片段发生交换的频率。
3.基因定位:确定基因在染色体上的位置。
主要是确定基因之间的距离和顺序。
4.符合系数:指理论交换值与实际交换值的比值,符合系数经常变动于0—1之间。
5.干扰(interference):一个单交换发生后,在它邻近再发生第二个单交换的机会就会减少的现象。
6.连锁遗传图(遗传图谱):将一对同源染色体上的各个基因的位置确定下来,并绘制成图的叫做连锁遗传图。
7.连锁群(linkage group):存在于同一染色体上的基因群。
8.性连锁(sex linkage):指性染色体上的基因所控制的某些性状总是伴随性别而遗传的现象,又称伴性遗传(sex-linked inheritance)。
9.性染色体(sex-chromosome):与性别决定有直接关系的染色体叫做性染色体。
10.常染色体(autosome):性染色体以外其他的染色体称为常染色体。
同配性别11.限性遗传(sex-limited inheritance):是指位于Y染色体(XY型)或W染色体(ZW型)上的基因所控制的遗传性状只限于雄性或雌性上表现的现象。
12.从性遗传(sex-influenced inheritance):常染色体上基因所控制的性状,在表现型上受个体性别的影响,只出现于雌方或雄方;或在一方为显性,另一方为隐性的现象。
13.交叉遗传:父亲的性状随着X染色体传给女儿的现象。
14.连锁遗传:指在同一同源染色体上的非等位基因连在一起而遗传的现象。
(二) 是非题:1.雄果蝇完全连锁是生物界少见的遗传现象。
这仅指X染色体上的连锁群而言。
因为它的X染色体只有一条,所以,不会发生交换。
(-)2.基因连锁强度与重组率成反比。
(+)3.基因型+ C/Sh +的个体在减数分裂中有6%的花粉母细胞在Sh和C之间形成一个交叉,那么,所产生的重组型配子++和 Sh C 将各占3%。

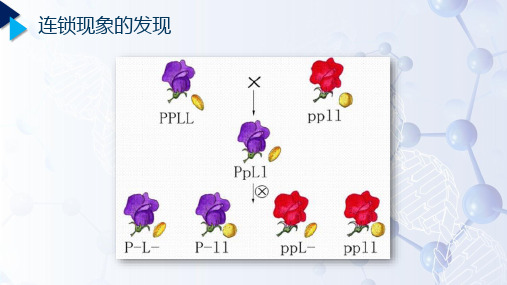
连锁现象的发现一、孟德尔定律的实质图6-1 分离规律的实质图6-2 自由组合规律的实质二、人类和黑腹果蝇的染色体数与基因数人类(Homo sapiens )染色体(男性)(2n=46,约3.5万个基因)黑腹果蝇(Drosophila melanogaster )(2n=8,约1.36万个基因)三、连锁基因与连锁群位于同一条染色体上的基因,称为连锁基因(linkage gene )同一染色体上的所有基因,构成了1个连锁群(linkage group)1连锁现象的发现3基因在染色体上排列2连锁基因的遗传规律连锁基因连锁群4基因定位与染色体作图四、连锁现象的发现1906年彼特逊(Bateson )和庞尼特(Punnett)最先发现的。
香豌豆(Lathyrus odoratus )紫花长花粉粒红花圆花粉粒花色花粉粒形状◆F2出现4种表型,单个性状的遗传均符合3:1的分离比;◆两个性状的遗传不符合自由组合9:3:3:1的比例。
亲组合与两亲本性状组合(紫圆和红长)相同的亲组合类型实际观测数高于理论数;重组合两种性状重新组合出现的(紫长和红圆)重组合类型的观测数少于理论数;◆两性状的遗传不符合9:3:3:1的自由组合比例;◆单个性状的遗传均符合3:1的分离比;◆重组合(紫花圆花粉和红花长花粉粒)实际观测数少于理论数;重组合◆亲组合(紫花长花粉和红花圆花粉)的实际观测数高于理论数;亲组合七、两性状遗传是否符合自由组合规律的检验⚫卡方适合度检验统计推断:df=k-1=3时,χ2 0.01=11.35,χ2>>χ2 0.01,p < < 0.01说明两性状的遗传不符合9:3:3:1的自由组合(p < 0.01)。
∑−==EE O 221-k df )(χ=3371.58八、彼特逊和庞尼特的解释两对基因在杂交子代中并不是随机的自由组合。
☐相引(Coupling):原来属于同一亲本的两个基因更倾向于进入同一配子中;☐互斥(repulsion):原来属于不同亲本的两个基因之间在形成配子时相互排斥;相引相互斥相。


第五章连锁遗传和性连锁(一) 名词解释:1.交换:指同源染色体的非姊妹染色单体之间的对应片段的交换,从而引起相应基因间的交换与重组。
2.交换值(重组率):指同源染色体的非姊妹染色单体间有关基因的染色体片段发生交换的频率。
3.基因定位:确定基因在染色体上的位置。
主要是确定基因之间的距离和顺序。
4.符合系数:指理论交换值与实际交换值的比值,符合系数经常变动于0—1之间。
5.干扰(interference):一个单交换发生后,在它邻近再发生第二个单交换的机会就会减少的现象。
6.连锁遗传图(遗传图谱):将一对同源染色体上的各个基因的位置确定下来,并绘制成图的叫做连锁遗传图。
7.连锁群(linkage group):存在于同一染色体上的基因群。
8.性连锁(sex linkage):指性染色体上的基因所控制的某些性状总是伴随性别而遗传的现象,又称伴性遗传(sex-linkedinheritance)。
9.性染色体(sex-chromosome):与性别决定有直接关系的染色体叫做性染色体。
10.常染色体(autosome):性染色体以外其他的染色体称为常染色体。
同配性别11.限性遗传(sex-limited inheritance):是指位于Y染色体(XY型)或W染色体(ZW型)上的基因所控制的遗传性状只限于雄性或雌性上表现的现象。
12.从性遗传(sex-influenced inheritance):常染色体上基因所控制的性状,在表现型上受个体性别的影响,只出现于雌方或雄方;或在一方为显性,另一方为隐性的现象。
13.交叉遗传:父亲的性状随着X染色体传给女儿的现象。
14.连锁遗传:指在同一同源染色体上的非等位基因连在一起而遗传的现象。
(二) 是非题:1.雄果蝇完全连锁是生物界少见的遗传现象。
这仅指X染色体上的连锁群而言。
因为它的X染色体只有一条,所以,不会发生交换。
(-)2.基因连锁强度与重组率成反比。
(+)3.基因型+ C/Sh +的个体在减数分裂中有6%的花粉母细胞在Sh和C之间形成一个交叉,那么,所产生的重组型配子++和 Sh C 将各占3%。
第五章连锁遗传分析5.1 性染色体与性别决定位于一对同源染色体上的非等位基因间的遗传关系以及性染色体上基因的遗传一、性染色体的发现1性染色体(sex chromosome)成对染色体中直接与性别决定有关的一个或一对染色体。
成对性染色体往往是异型的:形态、结构、大小、功能上都有所不同。
[1Y染色体;2XO与子;3染色体;1其中:X:A=1→雌性X:A=0.5→雄性X:A大于1的个体将发育成超雌性,小于0.5时发育成超雄性,介于两者则为间性(inter sex);并伴随着生活力、育性下降。
2 人类性别决定畸变.人类也存在由于性染色体组成异常而产生的性别畸变现象,对这些畸变现象的研究表明:与果蝇不同,人类的性别主要取决于是否存在Y染色体。
几种常见的人类性别畸变与症状表现:XO型(2n=45): 表现为女性,但出现唐纳氏(Turner’s)综合症;性别为女性,身材矮小(120-140cm),蹼颈、肘外翻和幼稚型生殖器官;部分表现为智力低下;卵巢发育不全、无生育能力。
XXY型(2n=47): 表现为男性,但出现克氏(Klinefelter’s)综合症;性别为男性,身材高大,第二性征类似女性,一般智力低下,睾丸发育不全、无生育能力。
XYY型(2n=47):性别为男性,智力稍差(也有智力高于一般人的)、较粗野、进攻性强,有生育能力。
四、其它类型的性别决定1 染色体倍数性决定蜜蜂等膜翅目的昆虫:性别取决于染色体的倍数性,并受到环境影响。
雄蜂为单倍体,孤雌生殖产生,形成配子时不进行减数分裂;雌蜂(蜂王)为二倍体,受精卵发育而来,并在幼虫期得到足够的蜂王浆(5天);如果幼虫期仅得到2-3天蜂王浆则发育为工蜂。
2 植物性别决定BaTs30℃条件下5.2 性连锁遗传(sex linkage)性连锁:也称为伴性遗传 (sex-linked inheritance),指位于性染色体上的基因所控制的某些性状总是伴随性别而遗传的现象;特指X 或Z 染色体上基因的遗传。
第五章染色体和连锁群本章重点一、连锁遗传:二对性状杂交有四种表现型,亲型多、重组型少;杂种产生配子数不等,亲型相等、重组型相等。
二、连锁和交换机理:粗线期交换、双线期交叉,非姐妹染色体交换。
三、交换值及其测定:重组配子数/总配子数;测交法测定,也可用F2 材料进行估计。
四、基因定位和连锁遗传图:确定位置、距离,基因位于染色体上;二点测验、三点测验;连锁群、连锁遗传图。
学时:91900年孟德尔遗传规律重新发现以后,生物界广泛重视,进行了大量试验。
其中有些属于两对性状的遗传结果不符合独立分配规律→摩尔根以果蝇为材料进行深入细致研究→提出连锁遗传规律→创立基因论→认为基因成直线排列在染色体上,进一步发展为细胞遗传学。
第一节连锁和交换一、连锁(一)连锁现象的发现1906年,贝特生(Bateson W.)和贝拉特(Punnett R. C.)在香豌豆的二对性状杂交试验中→首先发现性状连锁遗传现象。
第一个试验:P 紫花、长花粉粒(PPLL)×红花、圆花粉粒(ppll)↓F1紫、长PpLl↓⊗F2紫、长紫、圆红、长红、圆P_L_P_ll ppL_ppll总数实际个体数4831390393 13386952按9:3:3:1推算3910.51303.5 1303.5434.56952上述结果进行X2检验时,X2=3371.58,说明实计数与预计数差异极其显著,不可能由随机误差造成,应作重复实验。
以上结果表明F2 :①. 同样出现四种表现型;②. 不符合9:3:3:1;③. 亲本组合数偏多,重新组合数偏少(与理论数相比)。
第二个试验:P紫花、圆花粉粒(PPll)×红花、长花粉粒(ppLL)↓F1 紫、长PpLl↓⊗F2 紫、长紫、圆红、长红、圆P_L_P_ll ppL_ppll总数实际个体数22695971419按9:3:3:1推算235.878.578.526.2419结果与第一个试验情况相同。
X2=32.40,证明它仍然显著不符合孟德尔规律。
从这两个实验的数据可以看出一种共同的倾向:即与自由组合定律所预期结果相比较,F2中性状的亲本组合类型远远多于重组组合的类型,这等于说,在F1杂种形成配子时两对基因可能发生的4种类型中,有更多保持亲代原来组合的倾向,而且这种倾向与显隐性无关。
连锁遗传:原来亲本所具有的两个性状,在F2连系在一起遗传的现象。
相引组:甲乙两个显性性状,连系在一起遗传、而甲乙两个隐性性状连系在一起的杂交组合。
如:PL/pl。
相斥组:甲显性性状和乙隐性性状连系在一起遗传,而乙显性性状和甲隐性性状连系在一起的杂交组合。
如:Pl/pL。
(二)连锁遗传的解释试验结果是否受分离规律支配?第一个试验:紫花:红花(4831+390):(1338+393)=5221:1731 ≈3:1长花粉:短花粉(4831+393):(1338+390)=5224:1728 ≈3:1第二个试验:紫花:红花(226+95):(97+1)=321:98 ≈3:1长花粉:短花粉(226+97):(95+1)=323:96 ≈3:1*以上结果都受分离规律支配,但不符合独立分配规律。
*F2不符合9:3:3:1,则说明F1产生的四种配子不等。
可用测交法加以验证,∵测交后代的表现型种类以及比例可反映出F1配子的种类和比例。
利用测交法验证连锁遗传现象:特点:连锁遗传的表现为:♥两个亲本型配子数是相等,>50%;♥两个重组型配子数相等,<50%。
1.相引组:玉米(种子性状当代即可观察):有色、饱满CCShSh×无色、凹陷ccshsh↓F1有色饱满×无色凹陷CcShsh ccshsh配子CSh Csh cSh csh cshFt CcShsh Ccshsh ccShsh ccshsh有色饱满有色凹陷无色饱满无色凹陷总数粒数403214915240358368%48.2 1.8 1.848.2上述结果证实F1产生的四种配子不等:不是1:1:1:1 (25:25:25:25%)。
其中(Ft)亲本组合= (4032+4035)/8368×100% = 96.4%重新组合= (149+152)/8368×100% = 3.6%摩尔根(Morgan T. H.)等以果蝇为材料进行测交的结果:红眼长翅pr+pr+vg+vg+×紫眼正常翅prprvgvg↓F1 红眼长翅×紫眼正常翅pr+prvg+vg prprvgvg配子pr+vg+ prvg+ pr+vg prvg prvgFt pr+prvg+vg prprvg+vg pr+prvgvg prprvgvg红眼长翅紫眼长翅红眼正常翅紫眼正常翅总数个数133915415111952839结果:亲本组合=((1339+1195)/2839)×100%=89.26%重新组合=((154+151)/2839)×100%=10.74%∴证实F1产生的四种配子数不等,两种亲型配子多,两种重组型少,分别接近1:1。
亲本具有的两对非等位基因(Cc和Shsh 或pr+pr和vg+vg)不是独立分配,而是连系在一起遗传,如C-Sh、c-sh或pr+vg+和prvg常常连系在一起。
∴F1配子中总是亲本型配子(CSh和csh或pr+vg+和prvg)数偏多,重新组合配子(Csh、cSh或pr+vg和prvg+)数偏少。
2.相斥组:玉米:有色、凹陷CCshsh×无色、饱满ccShSh↓F1有色饱满×无色凹陷CcShsh ccshsh配子CSh Csh cSh csh cshFt CcShsh Ccshsh ccShsh ccshsh有色饱满有色凹陷无色饱满无色凹陷总数粒数638 21379 21096 67243785% 1.5 48.5 48.5 1.5结果:亲本组合=(21379+21096)/43785×100%=97.01%重新组合=(638+672)/843785×100%=2.99%∴相斥组的结果与相引组结果一致,同样证实F1所成的四种配子数不等,C-sh、c-Sh连系在一起的配子为多。
果蝇(相斥组):红眼正常翅pr+pr+vgvg×紫眼长翅prprvg+vg+↓F1 红眼长翅×紫眼正常翅pr+prvg+vg prprvgvg配子pr+vg+ prvg+ pr+vg prvg prvgFt pr+prvg+vg prprvg+vg pr+prvgvg prprvgvg红眼长翅紫眼长翅红眼正常翅紫眼正常翅总数个数157 1067 965 1462335结果:亲本组合=((1067+965)/2335)×100%=87.02%重新组合=((157+146)/2335)×100%=12.98%∴证实F1所产生的四种配子数不等。
♥上述结果均说明重组型配子数占总配子数的百分率< 50%。
重组率(交换值):重组型的配子百分数称为重组率。
♥当两对基因为连锁遗传时,其重组率总是<50。
∵相引组或相斥组中一对同源染色体的四条非姐妹染色单体,两个基因之间的染色体区段内仅有两条非姐妹染色单体发生交换(crossing over)。
∴重组型配子数目只是少数。
(三)完全连锁与不完全连锁:生物性状很多,控制这些性状的基因自然也多,而生物的染色体数目有限→必有许多基因位于同一染色体上→引起连锁遗传。
♣连锁:若干非等位基因位于同一染色体而发生连系遗传的现象。
♣完全连锁:同源染色体上非等位基因间不能发生非姐妹染色单体之间的交换→F1只产生两种亲型配子、其自交或测交后代个体的表现型均为亲本组合。
非等位基因完全连锁的情形很少(雄果蝇和雌家蚕),一般是不完全连锁。
不完全连锁(部分连锁):F1可产生多种配子,后代出现新性状的组合,但新组合较理论数为少。
如玉米颜色基因Cc和籽粒饱满度基因Shsh是位于玉米第9对染色体上的两对不完全连锁的非等位基因。
由连锁遗传例证中可见两个问题:1.相引组和相斥组都表现为不完全连锁,后代中均出现重组类型,且重组率很接近→其重组型配子是如何出现的?2.为何重组型配子数<亲型配子数,其重组率<50%?二、交换:1.交换:成对染色体非姐妹染色单体间基因的互换。
2.交换的过程:杂种减数分裂时期(前期I的粗线期)。
3.根据染色体细胞学行为和基因位置上的变化关系→可以说明连锁和交换的实质。
例如玉米有色饱满基因:(1)基因在染色体上呈直线排列;(2)等位基因位于一对同源染色体的两个不同成员上;(3)同源染色体上有两对处于不同位置上的非等位基因;(4)减数分裂前期I的偶线期中各对同源染色体配对(联会)→粗线期已形成四合体→双线期同源染色体出现交叉→非姐妹染色单体粗线期时发生交换→随机分配到子细胞内→发育成配子。
在全部孢母细胞中,各联会的同源染色体在C与Sh基因间不可能全部都发生交换,故重组率<50%;例如玉米F1的100个孢母细胞中,交换发生在Cc和Shsh相连区段之内的有7个,则重组率为3.5 %。
亲本组合=((193+193)/400)×100%=96.5%重新组合=((7+7)/400)×100%=3.5%∴两对连锁基因间发生交换的孢母细胞的百分率,恰是交换配子(重组型配子)百分率的一倍。
带有亲本原有基因组合的配子比带有新的基因组合的配子多,而且每一对连锁基因的重组频率是固定的,或者说连锁强度是固定的,但不同的连锁基因间却是不同的。
交叉型假设(chiasmatype hypothesis):(1)在减数分裂前期,尤其是双线期,配对中的染色体不是简单地平行,而是在某些点上出现交叉。
(2)相互连锁的两个基因位于同一染色体的不同位置,如果这两个位置之间发生交换,就导致着两个连锁基因的重组(recombination)这个学说的核心是:交叉是交换的结果,而不是交换的原因,也就是说,遗传学上的交换发生在细胞学上的交叉出现之前。
交换的解释:(1)交换是相等的,一般不多一个基因,不少一个基因。
(2)交换发生在减数分裂的双线期,即发生在非姊妹染色单体之间。
(3)在连锁的基因间每发生一个交叉,只有一半的重组类型。
交换值愈小时,基因间的连锁愈易察觉;交换值愈大时,基因间的连锁愈难察觉。
第二节交换值及其测定一、交换值(重组率)交换值(重组率):指同源染色体非姐妹染色单体间有关基因的染色体片段发生交换的频率,一般利用重新组合配子数占总配子数的百分率进行估算。
交换值(%)=(重新组合配子数/总配子数)×100二、交换值的测定(一)测交法:上例玉米测交:相引组交换值为3.6%,两种重组配子各1.8 %;相斥组交换值为2.99%,两种重组配子各1.5 %。