第七章 转录产物的加工修饰及转运降解
- 格式:ppt
- 大小:2.25 MB
- 文档页数:61

转录后加工名词解释
转录后加工是指在基因组中进行转录的过程后,对转录产物(RNA分子)进行进一步的修饰和加工的过程。
转录是指在DNA模板上合成RNA分子的过程,而转录后加工则是在RNA分子合成完成后对其进行一系列的修饰和处理。
转录后加工的目的是为了产生成熟的RNA分子,使其能够发挥特定的功能。
在转录后加工过程中,RNA分子经历剪接、修饰和运输等多个步骤,以形成成熟的RNA分子。
剪接是转录后加工中最重要的步骤之一。
在剪接过程中,RNA 分子的内含子(非编码区域)会被剪除,而外显子(编码区域)则会被保留下来。
这样一来,通过剪接,一个基因可以产生多个不同的成熟RNA分子,从而扩大了基因的功能和多样性。
除了剪接,转录后加工还包括其他的修饰过程。
例如,RNA分子可能会经历5'端帽子的添加和3'端的聚腺苷酸尾巴的加入,这些修饰可以保护RNA分子免受降解,并有助于其在细胞内的稳定性和转运过程中的识别。
此外,转录后加工还可以包括RNA编辑、互补RNA合成和核糖体扫描等过程。
RNA编辑是指在转录后,RNA分子中的碱基序列可以发生改变,从而导致RNA分子的信息内容发生变化。
互补RNA合成是指利用RNA分子作为模板合成互补的DNA分子。
核糖体扫描是指RNA分子被核糖体识别并翻译成蛋白质的过程。
总的来说,转录后加工是一系列对转录产物进行修饰和加工的过程,通过这些过程,RNA分子可以获得特定的功能和稳定性,从而发挥其在细胞中的重要作用。

第七讲原核生物的基因调控科学家把这个从DNA到蛋白质的过程称为基因表达(gene expression),对这个过程的调节就称为基因表达调控(gene regulation或gene control)。
要了解动、植物发展发育的规律、形态布局特征和生物学功能,就必需弄清楚基因表达调控的时间和空间概念,掌握了基因表达调控的奥秘,我们手中就有了一把揭示生物学微妙的金钥匙。
基因表达调控主要暗示在以下几个方面:①转录程度上的调控(transcriptional regulation);②mRNA加工成熟程度上的调控(differential processing of RNAtranscript);③翻译程度上的调控(differential translation of mRNA).原核生物中,营养状况(nutritionalstatus)和环境因素(environmental factor)对基因表达起着举足轻重的影响。
在真核生物尤其是高等真核生物中,激素程度(hormone level)和发育阶段(developmental stage)是基因表达调控的最主要手段,营养和环境因素的影响力大为下降。
二、基因表达调控的底子道理〔一〕基因表达的多级调控基因的布局活化、转录起始、转录后加工及转运、mRNA降解、翻译及翻译后加工及蛋白质降解等均为基因表达调控的控制点。
可见,基因表达调控是在多级程度长进行的复杂事件。
此中转录起始是基因表达的底子控制点。
四个底子的调控点:〔1〕基因布局的活化。
DNA表露碱基后RNA聚合酶才能有效结合。
活化状态的基因暗示为:1.对核酸酶敏感;2.结合有非组蛋白及修饰的组蛋白;3.低甲基化。
〔2〕转录起始。
最有效的调节环节,通过DNA元件与调控蛋白彼此作用来调控基因表达。
〔3〕转录后加工及转运。
RNA编纂、剪接、转运。
〔4〕翻译及翻译后加工。
翻译程度可通过特异的蛋白因子阻断mRNA 翻译翻译后对蛋白的加工、修饰也是底子调控环节。

基因的转录、转录后加工及逆转录转录(transcription) 是以DNA单链为模板,NTP为原料,在DNA依赖的RNA 聚合酶催化下合成RNA链的过程。
与DNA勺复制相比,有很多相同或相似之处,亦有其特点,它们之间的异同可简要示于表13-1转录的模板是单链DNA与复制的模板有较多的不同特点,引出了下列相关概念。
转录过程只以基因组DNA中编码RNA(mRNAtRNA rRNA及小RNA 的区段为模板。
把DNA分子中能转录出RNA的区段,称为结构基因(structure gene)。
结构基因的双链中,仅有一股链作为模板转录成RNA称为模板链(template strand),也称作Watson(W链(Watson strand)、负(-)链(minus strand) 或反意义链(antisense strand) 。
与模板链相对应的互补链,其编码区的碱基序列与mRN的密码序列相同(仅T、U互换),称为编码链(coding strand),也称作Crick (0链(Crick strand )、正(+)链(plus strand),或有意义链(sense strand)。
不同基因的模板链与编码链,在DNA分子上并不是固定在某一股链,这种现象称为不对称转录(asymmetric transcription) 。
模板链在相同双链的不同单股时,由于转录方向都从5'f 3',表观上转录方向相反,如图13-1 o与DNA复制类似,转录过程在原核生物和真核生物中所需的酶和相关因子有所不同,转录过程及转录后的加工修饰亦有差异。
下面的讨论中将分别叙述。
? 参与转录的酶转录酶(transcriptase )是依赖DNA的RNA聚合酶(DNA dependent RNA polymerase,DDRP,亦称为DNA指导的RNA聚合酶(DNA directed RNA polymerase ),简称为RNA聚合酶(RNA pol)。


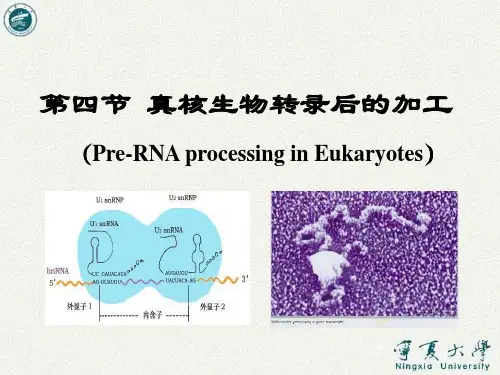
第二节RNA转录后的加工与修饰不论原核或真核生物的rRNAs都是以更为复杂的初级转录本形式被合成的,然后再加工成为成熟的RNA 分子。
然而绝大多数原核生物转录和翻译是同时进行的,随着mRNA开始的DNA上合成,核蛋白体即附着在mRNA上并以其为模板进行蛋白质的合成,因此原核细胞的mRNA并无特殊的转录后加工过程,相反,真核生物转录和翻译在时间和空间上是分天的,刚转录出来的mRNA是分子很大的前体,即核内不均一RNA。
hnRNA 分子中大约只有10%的部分转变成成熟的mRNA,其余部分将在转录后的加工过程中被降解掉。
(一)mRNA的加工修饰原核生物中转录生成的mRNA为多顺反子,即几个结构基因,利用共同的启动子和共同终止信号经转录生成一条mRNA,所以此mRNA分子编码几种不同的蛋白质。
例如乳糖操纵子上的Z、Y及A基因,转录生成的mRNA可翻译生成三种酶,即半乳糖苷酶,透过酶和乙酰基转移酶。
原核生物中没有核模,所以转录与翻译是连续进行的,往往转录还未完成,翻译已经开始了,因此原核生物中转录生成的mRNA没有特殊的转录后加工修饰过程。
真核生物转录生成的mRNA为单顺反子,即一个mRNA分子只为一种蛋白质分子编码。
真核生物mRNA的加工修饰,主要包括对5’端和3’端的修饰以及对中间部分进行剪接。
1.在5’端加帽成熟的真核生物mRNA,其结构的5’端都有一个m7G-PPNmN结构,该结构被称为甲基鸟苷的帽子。
如图17-9所示。
鸟苷通过5’-5’焦磷酸键与初级转录物的5’端相连。
当鸟苷上第7位碳原子被甲基化形成m7G-PPNmN时,此时形成的帽子被称为“帽0”,如果附m7G-PPNmN外,这个核糖的第“2”号碳上也甲基化,形成m7G-PPNm,称为“帽1”,如果5’末端N1和N2中的两个核糖均甲基化,成为m7G-PPNmPNm2,称为“帽2”。
从真核生物帽子结构形成的复杂可以看出,生物进化程度越高,其帽子结构越复杂。

★复习1.什么是转录?2.转录的模板、酶、原料、合成部位及方向、转录特点。
★新授:三、转录过程(一)转录起始阶段1.概况:RNA聚合酶结合到DNA模板;第一个NTP加入形成转录起始物2.相关概念*操纵子:由若干结构基因及其上游的调控序列组成的转录单位。
*启动子:RNA聚合酶全酶与DNA模板结合并启动转录的部位。
3.具体过程⑴根据模板链上核苷酸序列,NTP进入生成与DNA互补的RNA第一、第二位三磷酸核苷⑵RNA聚合酶催化第一、二位三磷酸核苷形成第一个3',5'-磷酸二酯键。
述:通常RNA链起始5'端总是三磷酸嘌呤核苷GTP或ATP,又以GTP更为常见。
4.举例――原核生物的转录起始●全酶(α2 ββ‘σ)结合启动子,σ70 识别-35区●RNA5'第一个核苷酸GTP(ATP)●第一个磷酸二酯键形成:pppGpN-3'OH●转录起始复合物:RNA pol(α2ββ'σ)-DNA- pppGpN-3'OH(二)链的延长1.概况:在起始阶段形成第一个磷酸二酯键后,σ亚基从转录起始物上脱落。
RNA聚合酶核心酶沿模板向下游(3'→5')移动,新生的RNA链按碱基互补配对原则(T→U),以5'→3' 的方向进行延伸。
述:合成的RNA自3'末端逐步延长。
合成出的RNA与DNA 形成杂交分子,约12bp,因结合不紧很易脱离。
随转录的向前进行,RNA链的5'端不断脱离模板链,然后模板链与编码链又恢复双螺旋结构。
2.转录泡:是由DNA双链,RNA聚合酶与新合成的转录本RNA局部形成的结构,它贯穿于延长过程的始终。
(三)转录终止1.概况:RNA聚合酶合成移到终止信号时停止,转录产物RNA从转录复合物中脱落。
2.原核生物转录终止的类型⑴ρ因子参与的转录终止述:r因子与RNA产物中富含C的部位结合,并诱使RNA聚合酶构象改变停止滑动;ρ因子的解螺旋酶活性,利于RNA 产物的释放。

简答2010-01-16 06:220、何为色氨酸操纵子弱化子与转录弱化作用?答:色氨酸操纵子位于转录起始部位的终止子结构称弱化子,色氨酸的浓度决定转录到达弱化子能否形成终止子发夹结构导致转录终止或转录继续。
1.简述乳糖操纵子的正负调控机制答案要点:包括正负调控两种(—)阻遏蛋白的负调控①当细胞内有诱导物时,诱导物结合阻遏蛋白,此刻聚合酶与启动子形成开放式启动子复合物转录乳糖操纵子结构基因。
②当无诱导物时,阻遏蛋白结合与启动子与蛋白质部分重叠不转录。
(=)CAP正调控①当细胞内缺少葡萄糖时ATP→CAMP结合,CRP生成CAP与CAP位点结合,增前RNA聚合酶转录活性。
②当有葡萄糖存在时CAMP分解多合成少,CAP不与启动子上的CAP位点结合RNA聚合酶不与操纵区结合无法起始转录结构基因表达下降。
2.试比较原核和真核细胞的mRNA的异同答案要点:①真核生物5…端有帽子结构大部分成熟没mRNA 还同时具有3‟多聚A尾巴,原核一般没有;②原核的没mRNA 可以编码几个多肽真核只能编码一个。
③原核生物以AUG作为起始密码有时以GUG,UUG作为起始密码,真核几乎永远以AUG作为起始密码。
④原核生物mRNA半衰期短,真核长。
⑤原核生物以多顺反子的形式存在,真核以单顺反子形式存在3基因敲除的基本程序?答:通过DNA同源重组,使得胚胎干细胞特定的内源基因被破坏而造成功能丧失,然后通过胚胎干细胞介导得到该基因丧失的小鼠模型的过程称为基因敲除。
1、打靶载体的构建:同源序列要足够长,要含有筛选用的标志基因。
2、胚胎干细胞的体外培养3、打靶载体导入胚胎干细胞4、同源重组胚胎干细胞的筛选5、基因敲除胚胎干细胞注射入胚泡6、胚泡植入假孕小鼠的子宫中7、杂交育种获得纯合的基因敲除动物、4简述真核生物mRNA的帽子结构及其功能。
当RNA polII 聚合的转录产物达到约25nt长时,在其5‟端加上了一个7-甲基鸟苷(m7G),该7-甲基鸟苷称为帽子结构,是以5‟- 5‟方向相连的,用来防止5‟-外切酶的攻击,但却有利于剪接、转运和翻译的进行。

第二节 RNA转录后得加工与修饰不论原核或真核生物得rRNAs都就是以更为复杂得初级转录本形式被合成得,然后再加工成为成熟得RNA分子。
然而绝大多数原核生物转录与翻译就是同时进行得,随着mRNA开始得DNA上合成,核蛋白体即附着在mRNA上并以其为模板进行蛋白质得合成,因此原核细胞得mRNA并无特殊得转录后加工过程,相反,真核生物转录与翻译在时间与空间上就是分天得,刚转录出来得mRNA就是分子很大得前体,即核内不均一RNA。
hnRNA分子中大约只有10%得部分转变成成熟得mRNA,其余部分将在转录后得加工过程中被降解掉。
(一)mRNA得加工修饰原核生物中转录生成得mRNA为多顺反子,即几个结构基因,利用共同得启动子与共同终止信号经转录生成一条mRNA,所以此mRNA分子编码几种不同得蛋白质。
例如乳糖操纵子上得Z、Y及A基因,转录生成得mRNA可翻译生成三种酶,即半乳糖苷酶,透过酶与乙酰基转移酶。
原核生物中没有核模,所以转录与翻译就是连续进行得,往往转录还未完成,翻译已经开始了,因此原核生物中转录生成得mRNA没有特殊得转录后加工修饰过程。
真核生物转录生成得mRNA为单顺反子,即一个mRNA分子只为一种蛋白质分子编码。
真核生物mRNA得加工修饰,主要包括对5’端与3’端得修饰以及对中间部分进行剪接。
1.在5’端加帽成熟得真核生物mRNA,其结构得5’端都有一个m7G-PPNmN结构,该结构被称为甲基鸟苷得帽子。
如图17-9所示。
鸟苷通过5’-5’焦磷酸键与初级转录物得5’端相连。
当鸟苷上第7位碳原子被甲基化形成m7G-PPNmN时,此时形成得帽子被称为“帽0”,如果附m7G-PPNmN外,这个核糖得第“2”号碳上也甲基化,形成m7G-PPNm,称为“帽1”,如果5’末端N1与N2中得两个核糖均甲基化,成为m7G-PPNmPNm 2,称为“帽2”。
从真核生物帽子结构形成得复杂可以瞧出,生物进化程度越高,其帽子结构越复杂。

第二节RNA转录后的加工取建饰之阳早格格创做不管本核或者真核死物的rRNAs皆是以更为搀纯的初级转录本形式被合成的,而后再加工成为老练的RNA分子.然而绝大普遍本核死物转录战翻译是共时举止的,随着mRNA开初的DNA上合成,核蛋黑体即附着正在mRNA 上并以其为模板举止蛋黑量的合成,果此本核细胞的mRNA并不特殊的转录后加工历程,差同,真核死物转录战翻译正在时间战空间上是分天的,刚刚转录出去的mRNA是分子很大的前体,即核内不均一RNA.hnRNA分子中约莫惟有10%的部分转形成老练的mRNA,其余部分将正在转录后的加工历程中被落解掉.(一)mRNA的加工建饰本核死物中转录死成的mRNA为多逆反子,即几个结构基果,利用共共的开用子战共共终止旗号经转录死成一条mRNA,所以此mRNA分子编码几种分歧的蛋黑量.比圆乳糖把持子上的Z、Y及A基果,转录死成的mRNA可翻译死成三种酶,即半乳糖苷酶,透过酶战乙酰基变化酶.本核死物中不核模,所以转录取翻译是连绝举止的,往往转录还已完毕,翻译已经开初了,果此本核死物中转录死成的mRNA不特殊的转录后加工建饰历程.真核死物转录死成的mRNA为单逆反子,即一个mRNA分子只为一种蛋黑量分子编码.真核死物mRNA的加工建饰,主要包罗对付5’端战3’端的建饰以及对付中间部分举止剪交.1.正在5’端加帽老练的真核死物mRNA,其结构的5’端皆有一个m7GPPNmN结构,该结构被称为甲基鸟苷的帽子.如图179所示.鸟苷通过5’5’焦磷酸键取初级转录物的5’端贯串.当鸟苷上第7位碳本子被甲基化产死m7GPPNmN时,此时产死的帽子被称为“帽0”,如果附m7GPPNmN中,那个核糖的第“2”号碳上也甲基化,产死m7GPPNm,称为“帽1”,如果5’终端N1战N2中的二个核糖均甲基化,成为m7GPPNmPNm2,称为“帽2”.从真核死物帽子结构产死的搀纯不妨瞅出,死物进化程度越下,其帽子结构越搀纯.图179Posttranscriptional modification of mRNa showing the 7methylguanosine cap and polyA tail.真核死物mRNA 5’端帽子结构的要害性正在于它是mRNa 搞为翻译起初的需要的结构,对付核糖体对付mRNA的辨别提供了旗号,那种帽子结构还大概减少mRNA的宁静性,呵护mRNa 免遭5’中切核酸酶的攻打.2.正在3’端加尾大普遍的真核mRNA 皆有3’端的多散尾巴(A),多散(A)尾巴约莫为200bp.多散(A)屠巴不是由DNA编码的,而是转录后正在核内加上去的.受polyA散合酶催化,该酶能辨别,mRNa 的游离3’OH端,并加上约200个A残基.连年去已知,正在大普遍真核基果的3’一端有一个AATAA序列,那个序列是mRNa 3’端加polyA尾的旗号.靠核酸酶正在此旗号下游1015碱基中切断磷酸二酯键,正在polyA散合酶催化下,正在3’OH上逐一引进100200个A 碱基.闭于polyA尾巴的功能问题纵然通过极其广大的探索,然而还不真足收会.有人推测polyA大概取mRNA从细胞核转收到细胞量有闭,然而是相称数量,的不polyA屠巴的mRNA如组蛋黑mRNA,也照样通过核膜加进细胞量.另有人认为那种结构对付真核mRNA的翻译效用具备某种效用,并能宁静mRNA结构,脆持一定的死物半衰期.3.mRNA前体(hnRNA)的拼交本核死物的结构基果是连绝编码序列,而真核死物基果往往是断裂基果,即编码一个蛋黑量分子的核苷酸序列被多个拔出片断所隔开,一个真核死物结构基果中内含子的数量,往往取那个基果的大小有闭,比圆胰岛素是一个很小的蛋黑量,它结构基果惟有二个内含子,而有些很大的蛋黑量,它的结构基果中不妨有几十个内含子.通过搀纯的历程后,切去内元,将有编码意思的核苷酸片段(Extron 中元也喊中隐子)连交起去(图1710).图1710Primary polymerase 11transcript of a eukaryote gene showing (a)introns after capping and addition of polyA tail.(b)Excision of introns to form the mature mRNA is calledsplicing.真核死物的结构的基果中具备可表黑活性的中隐子,也含有无表黑活性的内含子,然而内含子序列下是偶尔思的,越去越多的真验道明有许多基果中的内含子介进基果表黑调控,正在转录时,中隐子及内含子均转录到hnRNA 中.正在细胞核中hnRNA举止剪交效用,最先正在核酸内切酶效用下剪切掉内含子;而后正在连交酶效用下,将中隐子各部分连交起去,而形成老练的mRNA,那便是剪交效用,也有少量基果的hnRNA不需举止剪交效用,比圆α搞扰素基果,图1711以卵浑蛋黑基果为例,介绍一个典型的转录及加工历程.图1711卵浑蛋黑基果转录及加工历程图中中隐现以1、2、3、4……表示,内含子以A、B、C、D…表示mRNA的拼交,需要正在拼交部位有供拼交识别的守旧性强的普遍程序,通过对付100多种真核细胞基果的分解,创制中元战内元拼交部位部分碱基程序有一定的程序(睹表174).表17-4含有内元的转录产品其拼交处的碱基程序表中划线的碱基对付拼交辨别有要害效用,如将兔的β珠蛋黑的拼交部位的GT改为AT后,拼交反应即受到效用.mRNA前体拼体制图1712The RNA splicing mechanism.RNA splicing iscatalyzed by a spliceosome formed from the assembly ofU1,U2,U5,and sn RNPs(shown as green circles )plus other components (not shown).After assembly of the spliceosome ,the reaction occures in two speps:in step 1the branchpoint Anucleotide in the intron sequence,which is located colse to the 3'splice site ,attacks the 5'splice site and cleaves it;the cut 5'end of the intron sequence thereby becomes covalently linked to this A nucleotide,forming the branched nucleotide shown in Figure 855.In step 2 the 3'OH end of the first exon sequence,which was created in the first step,adds to the beginning of the second exon sequence,cleqving the RNA molecule at the 3'splice site;the two exon sequences are thereby joined to each other and the intron sequence is released ad a ribosone.These splicing reactions occur im the nucleus and gengerate mRNa molecules fromprimary RNA transcripts (mRNA precursor molecules).mRNA拼交反应需要有核内小分子RNA介进它们取蛋黑量产死的复合物称为小核糖核蛋黑颗粒,SnRNA分别被命名为U1,U2,U3,U4,U5,战U6RNA.SnRNA中的U2RNA由取内元左端拼交部位附近的UACUAA程序下度互补,产死一个环状结构,由特定的酶去辨别切除该环状结构,完毕拼交历程,如图1712所示.图1713Mechanim of mRNa splicing.Note that,for clarity,the process is shown in two stages;energy is not required for theprocess since transesterification reactions are involved.真核死物 mRNA前体正在剪交历程中,还不妨产死套索样的结构,正在内含子序列中常有一个分收部位的腺苷酸残基,它的2’OH不妨自动攻打内含子5’端取中隐子1连交的磷酸二酯键,切开了中噗子1,而腺苷酸本去已有3’,5’磷酸二酯键贯串的二个相邻的核苷酸残基,加上此3’,5’磷酸二酯键连交后,正在腺苷酸处出现了一个套索,已被切下的中隐子1的3’OH攻打内含子3’终端取中隐子2之间的3’,5’磷酸二酯键,键断裂后,内含子以套索的形式被节下去,此时中隐子1战中隐子2不妨连交起去(图1713).不管拼交历程怎么样,拼交必须极为透彻,可则会引导遗传疑息传播障碍,合成的蛋黑量大概丧得其平常的功能.尔国北圆广天里区是β天中海贫血的下收区,那是由于β珠蛋黑链的合成受到部分或者真足压制所引起的一种血黑蛋黑病.真验标明β珠蛋黑基果元1中核苷酸的面突变改变了平常拼交部位的碱基程序,截止制成过得部位的拼交.加工老练的mRNA虽能翻译,然而产品不是平常的β珠蛋黑,截止引起血黑蛋黑级结媾战功能的改变.(二)rRNA转录后加工本核死物rRNA转录后加工,包罗以下几圆里:①rRNA前体被大肠杆菌RNaseⅢ,RNaseE等剪切成一定链少的rRNA分子;②rRNA正在建饰酶催化下举止碱基建饰;③rRNA取蛋黑量分离产死核糖体的大、小亚基(睹图1714)图1714大肠杆菌rRNA前体的加工真核死物rRNA前体比本核死物大,哺乳动物的初级转录产品为45s,矮等真核死物的rRNA前体为38s,真核死物5sRNA前体独力于其余三种rRNA的基果转录(图1715).图1715真核死物rRNA前体的加工真核死物rRNA前体中含有拔出程序,rRNA前体要产死老练的rRNA,需要通过拼交反应.比圆,四膜虫的rRNA 前体的拼交是一种无酶催化的自动拼交历程.四膜虫基果组内,26srRNA编码的天区内有413bp的拔出程序.该插搁序列不妨不必耗能量从rRNA前体中被撤除.用SDS煮沸战用蛋黑酶中理等损害酶活性办法,皆不克不迭损害拼交活性,然而反应中Mg2+战鸟嘌呤核苷酸是必正在的.用32PGTP举止逃踪真验标明,起初历程是GTP正在拔出程序5’端爆收亲核反应,共时GMP取5’端切面的切除段产死磷酸二酯键并使本RNA断开.第二步是5’切面的中元3’O H 取3’切面的中元5’P共价连交,赢得老练的rRNA,被切除部分终尾环化,产死一个环状结构,共时从5’端去掉一个15核苷酸啐片.结余部分连交成399核苷酸的环状产品,再通过几步,终尾切下一个19个核苷酸的线性内含子序列即L19,它具备催化活性,上头的剪交效用,是由内含子自己的催化本量决断的(图1716).图1716四膜虫rRNA前体的自尔剪交那种rRNA的自己剪交反应给人们一个提示:即RNA 分子也有酶的催化活性.那背酶的化教真量是蛋黑量那一保守观念提出了挑拨.那种有酶催化活性的RNA分子命名为Ribozyme.T.Cech战S.Altman各自分别创制RNA具备催化效用,他们的创制对付于相识死命举止历程有要害意思.很大概正在本初死掷中,RNA所催化的断裂一连交反应是最早出现的催化历程.为此,他们共共赢得了1989年Nobel化教奖.从大普遍Ribozymw的结构中创制一些特性,比圆:锤头状结构的RNA分子有13个守旧的核苷酸序列,如果它们中的碱基改变会使那种催化活性得去效用.根据那种特片,科教家们正在体中出计并人为合成那种RNA分子,用于抗肿瘤及抗病毒的真验中(图1717).图1717锤头结构模式图(三)tRNA转录后的加工建饰本核死物战真核死物刚刚转录死成的tRNA前体普遍无死物活性,需要举止①剪切战拼交②碱基建饰③3’OH连交ACC结构(图1718).图1718tRNA前体的加工①tRNA前体正在tRNA剪切酶的效用下,切成一定正在小的tRNA分子.大肠杆菌RNase P可特同剪切tRNA前体的5’旁程序,果此,该酶被称为tRNA5’老练酶.除了RNaseP 中,tRNA前体的剪切尚需要一个3’核酸内切酶,那可将tRNA前体3’端的一段核苷酸序列切下去.别的RNaseD是tRNA3’端老练酶.连年去的钻研标明大肠杆菌RNaseP是一种非常特殊的酶分子,它是由RNA战蛋黑量组成,迩去创制RNAaseP分子中的RNA部分正在某些条件下,不妨单独天催化tRNA前体的加工老练,那个创制战四膜虫tRNA 能自尔拼交被认为是近十年去死化范围内最令人饱舞的创制之一.道明RNA分子确具备酶的催化活性.通过剪切后的tRNA分子还要正在拼交酶效用下,将老练tRNA分子所需的片段拼起去.②老练的tRNA分子中有许多的密有碱基,果此tRNA正在甲基变化酶催化下,某些嘌呤死成甲基嘌呤如A→mA,G→mA.有些尿嘧啶还本为单氢尿嘧啶.尿嘧啶核苷转化不假尿嘧啶核苷.某些腺苷酸脱氨基为成为次黄嘌呤核苷酸(Ⅰ)③3’终端加上CCA:正在核苷酸变化酶效用下,3’终端取消各别碱基后,换上tRNA分子统一的CCAOH终端,完毕tRNA分子中的氨基酸臂结构.。

第一章绪论1.染色体具有哪些作为遗传物质的特征?答:①分子结构相对稳定;②能够自我复制,使亲子代之间保持连续性;③能够指导蛋白质的合成,从而控制整个生命过程;④能够产生可遗传的变异..2.什么是核小体?简述其形成过程。
答:由DNA和组蛋白组成的染色质纤维细丝是许多核小体连成的念珠状结构.核小体是由H2A,H2B,H3,H4各两个分子生成的八聚体和由大约200bp的DNA组成的。
八聚体在中间,DNA分子盘绕在外,而H1则在核小体外面核小体的形成是染色体中DNA压缩的第一阶段。
在核小体中DNA盘绕组蛋白八聚体核心,从而使分子收缩至原尺寸的1/7。
200bpDNA完全舒展时长约68nm,却被压缩在10nm的核小体中。
核小体只是DNA压缩的第一步。
核小体长链200bp→核酸酶初步处理→核小体单体200bp→核酸酶继续处理→核心颗粒146bp3简述真核生物染色体的组成及组装过程答:组成:蛋白质+核酸.组装过程:1,首先组蛋白组成盘装八聚体,DNA缠绕其上,成为核小体颗粒,两个颗粒之间经过DNA连接,形成外径10nm的纤维状串珠,称为核小体串珠纤维;2,核小体串珠纤维在酶的作用下形成每圈6个核小体,外径30nm的螺线管结构;3,螺线管结构再次螺旋化,形成超螺旋结构;4,超螺线管,形成绊环,即线性的螺线管形成的放射状环。
绊环在非组蛋白上缠绕即形成了显微镜下可见的染色体结构。
4. 简述DNA的一,二,三级结构的特征答:DNA一级结构:4种核苷酸的的连接及排列顺序,表示了该DNA分子的化学结构DNA二级结构:指两条多核苷酸链反向平行盘绕所生成的双螺旋结构DNA三级结构:指DNA双螺旋进一步扭曲盘绕所形成的特定空间结构6简述DNA双螺旋结构及其在现代分子生物学发展中的意义(1)DNA双螺旋是由两条互相平行的脱氧核苷酸长链盘绕而成的,多核苷酸的方向由核苷酸间的磷酸二酯键的走向决定,一条是5-——3,另一条是3---——5。
蛋白质合成中的转录后修饰及其生理学意义转录后修饰是指生物体在基因转录过程后,对所合成的RNA进行某些化学修饰。
这些修饰对蛋白质合成过程非常重要,也具有一定的生理学意义。
1. mRNA修饰转录后修饰的一种类型是对mRNA的修饰。
mRNA主要由四种核苷酸组成,即腺嘌呤(A)、胸腺嘧啶(U)、鸟嘌呤(G)和胞嘧啶(C)。
在mRNA的前体序列中,可能存在大量的嘌呤氧化酶、脱甲基化酶和甲基转移酶等酶类,对mRNA进行化学修饰。
这些RNA后转录修饰的过程,对mRNA的功能和生理学意义产生了很大的影响。
例如,mRNA上存在的特定修饰可能会影响翻译终止子的选择和启动子的效率,从而影响蛋白质的表达。
2. tRNA修饰另一个被广泛研究的RNA后转录修饰类型是对tRNA的修饰。
tRNA是带有氨基酸的RNA和蛋白质合成中必不可少的分子。
在tRNA修饰中,发现存在大量的tRNA甲基转移酶、tRNA二磷酸核糖化酶和tRNA修饰酶等酶类,对tRNA进行修饰。
这些修饰会直接影响tRNA在蛋白质合成中的地位。
例如,在tRNA的特异性子结合位点上的修饰可能会影响tRNA的识别、附着和释放,从而影响蛋白质的合成。
3. rRNA修饰除了mRNA和tRNA的修饰外,rRNA上的修饰也被广泛关注。
rRNA是与核糖体有关的RNA,对蛋白质合成起重要作用。
此外,rRNA结构复杂而独特,包括许多翻译相关基序,可能对转录后修饰有特殊的响应。
rRNA后转录修饰的类型包括脱甲基化、甲基化、2'-ribose甲基化、2'-ribose乙酰化和碳酸化等。
这些修饰会影响核糖体的结构、功能和招募,从而影响蛋白质的合成速率和选择性。
4. 修饰与代谢活跃度之间的关系需要注意的是,RNA后转录修饰和代谢活跃度之间存在巨大的相互影响。
在细胞代谢过程中,多种代谢反应正在进行。
这些反应可以提供ATP和其他核苷酸和代谢物作为RNA修饰的底物。
例如,在甲基化过程中,S-腺苷甲硫氨酸(SAM)是一个必须的因素,即SAM会转移甲基到RNA中,形成的还原型SAM在此过程中活化。
第二节RNA转录后的加工与修饰之青柳念文创作不管原核或真核生物的rRNAs都是以更为复杂的初级转录本形式被合成的,然后再加工成为成熟的RNA分子.然而绝大多数原核生物转录和翻译是同时停止的,随着mRNA 开端的DNA上合成,核蛋白体即附着在mRNA上并以其为模板停止蛋白质的合成,因此原核细胞的mRNA并没有特殊的转录后加工过程,相反,真核生物转录和翻译在时间和空间上是分天的,刚转录出来的mRNA是分子很大的前体,即核内不均一RNA.hnRNA分子中大约只有10%的部分转变成成熟的mRNA,其余部分将在转录后的加工过程中被降解掉.(一)mRNA的加工修饰原核生物中转录生成的mRNA为多顺反子,即几个布局基因,操纵共同的启动子和共同终止信号经转录生成一条mRNA,所以此mRNA分子编码几种分歧的蛋白质.例如乳糖把持子上的Z、Y及A基因,转录生成的mRNA可翻译生成三种酶,即半乳糖苷酶,透过酶和乙酰基转移酶.原核生物中没有核模,所以转录与翻译是持续停止的,往往转录还未完成,翻译已经开端了,因此原核生物中转录生成的mRNA没有特殊的转录后加工修饰过程.真核生物转录生成的mRNA为单顺反子,即一个mRNA分子只为一种蛋白质分子编码.真核生物mRNA的加工修饰,主要包含对5’端和3’端的修饰以及对中间部分停止剪接.1.在5’端加帽成熟的真核生物mRNA,其布局的5’端都有一个m7GPPNmN布局,该布局被称为甲基鸟苷的帽子.如图179所示.鸟苷通过5’5’焦磷酸键与初级转录物的5’端相连.当鸟苷上第7位碳原子被甲基化形成m7GPPNmN时,此时形成的帽子被称为“帽0”,如果附m7GPPNmN外,这个核糖的第“2”号碳上也甲基化,形成m7GPPNm,称为“帽1”,如果5’结尾N1和N2中的两个核糖均甲基化,成为m7GPPNmPNm2,称为“帽2”.从真核生物帽子布局形成的复杂可以看出,生物退化程度越高,其帽子布局越复杂.图179 Posttranscriptional modification of mRNa showing the 7methylguanosine cap and polyA tail.真核生物mRNA 5’端帽子布局的重要性在于它是mRNa 做为翻译起始的需要的布局,对核糖体对mRNA的识别提供了信号,这种帽子布局还能够增加mRNA的稳定性,呵护mRNa 免遭5’外切核酸酶的攻击.2.在3’端加尾大多数的真核mRNA 都有3’端的多聚尾巴(A),多聚(A)尾巴大约为200bp.多聚(A)屠巴不是由DNA编码的,而是转录后在核内加上去的.受polyA聚合酶催化,该酶能识别,mRNa 的游离3’OH端,并加上约200个A残基.近些年来已知,在大多数真核基因的3’一端有一个AATAA序列,这个序列是mRNa 3’端加polyA尾的信号.靠核酸酶在此信号下游1015碱基外切断磷酸二酯键,在polyA聚合酶催化下,在3’OH上逐一引入100200个A碱基.关于polyA尾巴的功能问题虽然颠末极其广泛的探索,但还不完全清楚.有人推测polyA能够与mRNA从细胞核转送到细胞质有关,但是相当数量,的没有polyA屠巴的mRNA如组蛋白mRNA,也照样通过核膜进入细胞质.还有人认为这种布局对真核mRNA的翻译效率具有某种作用,并能稳定mRNA布局,坚持一定的生物半衰期.3.mRNA前体(hnRNA)的拼接原核生物的布局基因是持续编码序列,而真核生物基因往往是断裂基因,即编码一个蛋白质分子的核苷酸序列被多个拔出片断所隔开,一个真核生物布局基因中内含子的数量,往往与这个基因的大小有关,例如胰岛素是一个很小的蛋白质,它布局基因只有两个内含子,而有些很大的蛋白质,它的布局基因中可以有几十个内含子.颠末复杂的过程后,切去内元,将有编码意义的核苷酸片段(Extron外元也叫外显子)毗连起来(图1710).图1710 Primary polymerase 11transcript of a eukaryote gene showing (a)introns after capping and addition of polyA tail.(b)Excision of introns to form the mature mRNA is called splicing.真核生物的布局的基因中具有可表达活性的外显子,也含有无表达活性的内含子,但内含子序列下是无意义的,越来越多的实验证明有许多基因中的内含子参与基因表达调控,在转录时,外显子及内含子均转录到hnRNA中.在细胞核中hnRNA停止剪接作用,首先在核酸内切酶作用下剪切掉内含子;然后在毗连酶作用下,将外显子各部分毗连起来,而变成成熟的mRNA,这就是剪接作用,也有少数基因的hnRNA不需停止剪接作用,例如α干扰素基因,图1711以卵清蛋白基因为例,先容一个典型的转录及加工过程.图1711 卵清蛋白基因转录及加工过程图中外显示以1、2、3、4……暗示,内含子以A、B、C、D…暗示mRNA的拼接,需要在拼接部位有供拼接识此外守旧性强的一致顺序,通过对100多种真核细胞基因的分析,发现外元和内元拼接部位部分碱基顺序有一定的规律(见表174).表17-4 含有内元的转录产品其拼接处的碱基顺序表中划线的碱基对拼接识别有重要作用,如将兔的β珠蛋白的拼接部位的GT改为AT后,拼接反应即受到影响.mRNA前体拼机制图1712 The RNA splicing mechanism.RNA splicing is catalyzed by a spliceosome formed from the assembly of U1,U2,U5,and sn RNPs(shown as greencircles )plus other components (not shown).After assembly of the spliceosome ,the reaction occuresin two speps:in step 1the branchpoint A nucleotide in the intron sequence,which is located colse to the 3'splice site ,attacks the 5'splice site and cleaves it;the cut 5'end of the intron sequence thereby becomes covalently linked to this A nucleotide,forming the branched nucleotide shown in Figure 855.In step 2 the 3'OH end of the first exon sequence,which was created in the first step,adds to the beginning of the second exon sequence,cleqving the RNA molecule at the 3'splice site;the two exon sequences are thereby joined to each other and the intron sequence is released ad a ribosone.These splicing reactions occur im the nucleus and gengerate mRNa molecules from primary RNA transcripts (mRNA precursor molecules).mRNA拼接反应需要有核内小分子RNA参与它们与蛋白质形成的复合物称为小核糖核蛋白颗粒,SnRNA分别被定名为U1,U2,U3,U4,U5,和U6RNA.SnRNA中的U2RNA由与内元右端拼接部位附近的UACUAA顺序高度互补,形成一个环状布局,由特定的酶来识别切除该环状布局,完成拼接过程,如图1712所示.图1713 Mechanim of mRNa splicing.Note that,for clarity,the process is shown in two stages;energy is not required for the process since transesterification reactions are involved.真核生物 mRNA前体在剪接过程中,还可以形成套索样的布局,在内含子序列中常有一个分支部位的腺苷酸残基,它的2’OH可以自动攻击内含子5’端与外显子1毗连的磷酸二酯键,切开了外噗子1,而腺苷酸原来已有3’,5’磷酸二酯键相连的两个相邻的核苷酸残基,加上此3’,5’磷酸二酯键毗连后,在腺苷酸处出现了一个套索,已被切下的外显子1的3’OH攻击内含子3’结尾与外显子2之间的3’,5’磷酸二酯键,键断裂后,内含子以套索的形式被节下来,此时外显子1和外显子2可以毗连起来(图1713).不管拼接过程如何,拼接必须极为切确,否则会导致遗传信息传递障碍,合成的蛋白质能够丧失其正常的功能.我国北方广大地区是β地中海贫血的高发区,这是由于β珠蛋白链的合成受到部分或完全抑制所引起的一种血红蛋白病.实验标明β珠蛋白基因元1中核苷酸的点突变改变了正常拼接部位的碱基顺序,成果造成错误部位的拼接.加工成熟的mRNA虽能翻译,但产品不是正常的β珠蛋白,成果引起血红蛋白级布局和功能的改变.(二)rRNA转录后加工原核生物rRNA转录后加工,包含以下几方面:①rRNA 前体被大肠杆菌R NaseⅢ,RNaseE等剪切成一定链长的rRNA 分子;②rRNA在修饰酶催化下停止碱基修饰;③rRNA与蛋白质连系形成核糖体的大、小亚基(见图1714)图1714 大肠杆菌rRNA前体的加工真核生物rRNA前体比原核生物大,哺乳动物的初级转录产品为45s,低等真核生物的rRNA前体为38s,真核生物5sRNA前体独立于其他三种rRNA的基因转录(图1715).图1715 真核生物rRNA前体的加工真核生物rRNA前体中含有拔出顺序,rRNA前体要形成成熟的rRNA,需要颠末拼接反应.例如,四膜虫的rRNA前体的拼接是一种无酶催化的自动拼接过程.四膜虫基因组内,26srRNA编码的区域内有413bp的拔出顺序.该插放序列可以不必耗能量从rRNA前体中被除掉.用SDS煮沸和用蛋白酶外理等破坏酶活性法子,都不克不及破坏拼接活性,但反应中Mg2+和鸟嘌呤核苷酸是必在的.用32PGTP停止追踪实验标明,起始过程是GTP在拔出顺序5’端发生亲核反应,同时GMP与5’端切点的切除段形成磷酸二酯键并使原RNA断开.第二步是5’切点的外元3’OH与3’切点的外元5’P共价毗连,获得成熟的rRNA,被切除部分最后环化,形成一个环状布局,同时从5’端去掉一个15核苷酸啐片.剩余部分毗连成399核苷酸的环状产品,再颠末几步,最后切下一个19个核苷酸的线性内含子序列即L19,它具有催化活性,上面的剪接作用,是由内含子自己的催化性质决议的(图1716).图1716 四膜虫rRNA前体的自我剪接这种rRNA的自身剪接反应给人们一个提示:即RNA分子也有酶的催化活性.这向酶的化学实质是蛋白质这一传统概念提出了挑战.这种有酶催化活性的RNA分子定名为Ribozyme.T.Cech和S.Altman各自分别发现RNA具有催化作用,他们的发现对于懂得生命停止过程有重要意义.很能够在原始生射中,RNA所催化的断裂一毗连反应是最早出现的催化过程.为此,他们共同获得了1989年Nobel化学奖.从大多数Ribozymw的布局中发现一些特征,例如:锤头状布局的RNA分子有13个守旧的核苷酸序列,如果它们中的碱基改变会使这种催化活性失去作用.根据这种特片,迷信家们在体外没计并人工合成这种RNA分子,用于抗肿瘤及抗病毒的实验中(图1717).图1717 锤头布局形式图(三)tRNA转录后的加工修饰原核生物和真核生物刚转录生成的tRNA前体一般无生物活性,需要停止①剪切和拼接②碱基修饰③3’OH毗连ACC布局(图1718).图1718 tRNA前体的加工①tRNA前体在tRNA剪切酶的作用下,切成一定在小的tRNA分子.大肠杆菌RNase P可特异剪切tRNA前体的5’旁顺序,因此,该酶被称为tRNA5’成熟酶.除了RNaseP 外,tRNA前体的剪切尚需要一个3’核酸内切酶,这可将tRNA前体3’端的一段核苷酸序列切下来.此外RNaseD是tRNA3’端成熟酶.近些年来的研究标明大肠杆菌RNaseP是一种非常特殊的酶分子,它是由RNA和蛋白质组成,最近发现RNAaseP分子中的RNA部分在某些条件下,可以单独地催化tRNA前体的加工成熟,这个发现和四膜虫tRNA能自我拼接被认为是近十年来生化范畴内最使人鼓舞的发现之一.说明RNA分子确具有酶的催化活性.颠末剪切后的tRNA分子还要在拼接酶作用下,将成熟tRNA分子所需的片段拼起来.②成熟的tRNA分子中有许多的稀有碱基,因此tRNA在甲基转移酶催化下,某些嘌呤生成甲基嘌呤如A→mA,G→mA.有些尿嘧啶还原为双氢尿嘧啶.尿嘧啶核苷转变不假尿嘧啶核苷.某些腺苷酸脱氨基为成为次黄嘌呤核苷酸(Ⅰ)③3’结尾加上CCA:在核苷酸转移酶作用下,3’结尾除去个别碱基后,换上tRNA分子统一的CCAOH结尾,完成tRNA 分子中的氨基酸臂布局.。