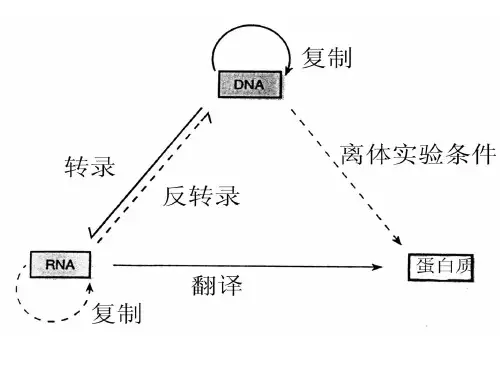
RNA转录后的加工与修饰
- 格式:docx
- 大小:214.47 KB
- 文档页数:8


简述rna转录后加工过程摘要:1.RNA转录后加工过程的概述2.RNA转录后加工的主要步骤a.剪接b.剪切c.RNA编辑d.RNA降解3.各步骤的功能和意义4.实例分析5.RNA转录后加工在生物体中的作用6.研究RNA转录后加工的意义和前景正文:在我们生物体内,基因通过转录过程将DNA信息转化为RNA,但这只是RNA生命历程中的第一步。
接下来,RNA要经历一系列复杂的加工过程,才能最终发挥其生物学功能。
这个过程被称为RNA转录后加工。
RNA转录后加工的主要步骤包括剪接、剪切、RNA编辑和RNA降解。
剪接是指将RNA前体分子中的内含子去除,并将外显子连接成成熟的RNA分子。
这一过程通过特定的酶家族,如剪接酶,来实现。
剪切是指在RNA分子的3"端添加poly(A)尾巴,这是几乎所有真核生物RNA的共同特征。
RNA编辑则是指在RNA分子上发生碱基改变,这一过程依赖于特定的编辑酶和相应的底物。
最后,RNA降解是指RNA分子在细胞内的分解过程,这对于调控RNA水平和维持细胞内稳态至关重要。
这些加工过程对于RNA最终的生物学功能具有重要意义。
以剪接为例,它能消除RNA前体中无功能的RNA片段,使成熟的RNA更具特异性和高效性。
同时,RNA编辑能够改变RNA的序列,从而影响其翻译效率和稳定性。
在生物体中,RNA转录后加工涉及多种生物过程,如基因表达调控、病毒复制和免疫反应等。
对RNA转录后加工的研究,有助于我们深入了解生命过程中的基因表达调控机制,为治疗疾病和开发新型药物提供理论依据。
随着生物科学技术的不断发展,对RNA转录后加工的研究将越来越深入。

rna转录后加工方式
RNA转录后加工(RNA post-transcriptional processing)是指在RNA分子合成之后,在细胞中对其进行修饰和修剪的过程。
这些加工方式可以使原始RNA分子成熟,并使其具有功能性。
以下是几种常见的RNA转录后加工方式:
剪接(Splicing):在真核生物中,基因的转录产物(前体mRNA)经过剪接过程,去除其中的内含子(intron),保留外显子(exon),从而形成成熟的mRNA分子。
剪接是通过剪接体(spliceosome)来完成的,其中包括snRNPs等辅助因子。
5'端修饰:RNA的5'端通常经过加上7-甲基鸟苷(7-methylguanosine)和三磷酸核苷酸链(PPP 链)的修饰,形成5'甲基鸟苷帽(5' cap)。
这个帽子在RNA稳定性、转运和翻译起重要作用。
3'端修饰:RNA的3'端通常经过加上聚腺苷酸(polyadenylation)的修饰。
这个poly(A)尾巴有助于RNA的稳定性、转运和翻译,并参与转录终止的过程。
RNA编辑:在一些生物体中,RNA的序列可以通过RNA编辑(RNA editing)进行改变。
这种编辑通常涉及碱基的替换、插入或删除,从而改变RNA的编码能力和功能。
RNA修饰:RNA分子可能会经历各种修饰,如甲基化、脱氨基、糖基化等。
这些修饰可以增强RNA的稳定性、调节翻译和识别,以及影响RNA的功能。
RNA转录后加工是一个复杂而精确的过程,它可以使原始的转录产物转化为功能性的RNA 分子。
这些加工方式对于基因表达调控和细胞功能起着重要的作用。

转录后RNA加工在基因表达调控中的作用在人类细胞中,转录后RNA加工是一项非常重要的过程,它决定了RNA是否能最终被翻译成蛋白质。
RNA加工包括了剪接、剪切、去除内含子和poly(A)尾处理等多个过程。
这些过程可以去除不必要的RNA,修饰RNA分子的稳定性、定位和功能等,从而在基因表达调控中起到极为关键的作用。
RNA剪接RNA剪接是RNA加工的一个重要分支,通过剪接可以将RNA前体分子中的内含子去除掉,从而促进mRNA的形成。
在剪接过程中,先由snRNP等辅助RNA 蛋白辅助组装剪接酶,然后在RNA前体分子的精确识别和定位下进行剪接。
在正常情况下,RNA前体分子的剪接是高度精确的,但在某些情况下,错误的剪接可能会导致某些疾病和癌症的发生。
RNA剪接通常分为五类,其中包括了acceptor位点、donor位点、branch位点、poly-pyrimidine三元序列和剪接增强子等多个元素。
这些元素都相互作用,形成一个复杂的调控网络,从而实现了RNA剪接的高度精确和复杂性。
RNA剪切RNA剪切是另外一个RNA加工的分支,它包括了一系列的过程,可以影响RNA的稳定性、生命周期和功能等。
相比于RNA剪接,RNA剪切更加复杂多变,因为它可能会发生在RNA分子的各种位置。
在RNA剪切过程中,主要存在三种不同类型的剪切,分别是特异剪切、选择性剪切和组合剪切。
特异剪切通常会有精确的位点被剪切,包括了接头剪切和凝集素剪切等多种类型。
选择性剪切则会受到外部调节因子的影响,从而在特定条件下发生剪切。
组合剪切则是特异剪切和选择性剪切的结合,通常会在多种条件下发生剪切。
RNA去除内含子内含子是RNA前体分子中不需要的RNA片段,通过内含子去除过程,这些RNA也可以被去除并最终形成mRNA。
内含子去除过程通常与RNA剪接紧密相连,同时又包括了许多独立的调控元素。
RNA去除内含子同样也会被一系列RNA蛋白调控,从而形成一个复杂的调控框架。




第二节RNA转录后的加工与修饰不论原核或真核生物的rRNAs都是以更为复杂的初级转录本形式被合成的,然后再加工成为成熟的RNA 分子。
然而绝大多数原核生物转录和翻译是同时进行的,随着mRNA开始的DNA上合成,核蛋白体即附着在mRNA上并以其为模板进行蛋白质的合成,因此原核细胞的mRNA并无特殊的转录后加工过程,相反,真核生物转录和翻译在时间和空间上是分天的,刚转录出来的mRNA是分子很大的前体,即核内不均一RNA.hnRNA 分子中大约只有10%的部分转变成成熟的mRNA,其余部分将在转录后的加工过程中被降解掉。
(一)mRNA的加工修饰原核生物中转录生成的mRNA为多顺反子,即几个结构基因,利用共同的启动子和共同终止信号经转录生成一条mRNA,所以此mRNA分子编码几种不同的蛋白质.例如乳糖操纵子上的Z、Y及A基因,转录生成的mRNA可翻译生成三种酶,即半乳糖苷酶,透过酶和乙酰基转移酶。
原核生物中没有核模,所以转录与翻译是连续进行的,往往转录还未完成,翻译已经开始了,因此原核生物中转录生成的mRNA没有特殊的转录后加工修饰过程。
真核生物转录生成的mRNA为单顺反子,即一个mRNA分子只为一种蛋白质分子编码。
真核生物mRNA的加工修饰,主要包括对5’端和3’端的修饰以及对中间部分进行剪接。
1.在5’端加帽成熟的真核生物mRNA,其结构的5’端都有一个m7G—PPNmN结构,该结构被称为甲基鸟苷的帽子。
如图17-9所示.鸟苷通过5’—5’焦磷酸键与初级转录物的5’端相连。
当鸟苷上第7位碳原子被甲基化形成m7G-PPNmN时,此时形成的帽子被称为“帽0”,如果附m7G-PPNmN外,这个核糖的第“2”号碳上也甲基化,形成m7G-PPNm,称为“帽1”,如果5'末端N1和N2中的两个核糖均甲基化,成为m7G—PPNmPNm2,称为“帽2”.从真核生物帽子结构形成的复杂可以看出,生物进化程度越高,其帽子结构越复杂。

rna转录后加工名词解释
RNA转录后加工是指对在转录过程新合成的RNA的前体分子,进行进一步的加工修饰,从而使其成为具有生物学活性的、成熟的RNA 分子的过程,主要包括剪接、化学修饰等方式。
1.可变剪切:通过不同的剪切方式使得同一个基因可以产生多个不同的成熟mRNA,最终产生不同的蛋白质,从而使转录本和蛋白质结构与功能具有多样性。
2.RNA编辑:属于修饰的一种,是指转录后的RNA在编码区发生碱基的加入、丢失或转换等现象,可以在RNA水平上增加一些原来DNA模板上没有编码的碱基,从而扩充遗传信息。
因此,经过剪切或者修饰等加工,遗传信息的含量以及多样性大大增加。
RNA后转录修饰及其生物学意义RNA后转录修饰指的是在RNA转录之后,通过各种化学反应对RNA分子进行化学修饰。
这些修饰可以改变RNA分子的结构和功能,在RNA转录后的生物学过程中起到至关重要的作用。
1. RNA后转录修饰的种类RNA后转录修饰的种类繁多,其中最常见的包括:甲基化、腺苷酸转换、亚硫酸化、伸长和端修饰等。
1.1 甲基化甲基化是RNA后转录修饰中最常见的一种。
RNA甲基化是指在RNA分子上添加甲基基团。
甲基化的位置和数量可能不同,从而影响RNA分子的稳定性、可行性、转录激活和基因表达等方面。
RNA 甲基化的存在对于体内细胞的抗衰老、肿瘤等方面的研究有着非常大的潜力。
1.2 腺苷酸转换RNA腺苷酸转换是RNA后转录修饰中最具代表性的一种。
通过改变RNA中腺苷酸的结构,进而影响RNA的功能和稳定性。
如RNA编辑便是腺苷酸转换的一种典型形式。
RNA编辑是指在RNA中单核苷酸的改变,从而影响RNA的功能。
RNA编辑是人类基因组学研究的一个重要热点。
1.3 亚硫酸化亚硫酸化是一种较少见的RNA后转录修饰。
该修饰是在RNA上加入亚硫酸基团,从而影响RNA的功能和稳定性。
这种修饰常常涉及到RNA分子的3′末端,对于RNA分子的转移特别重要。
1.4 伸长和端修饰RNA伸长和端修饰指的是在RNA分子两端进行修饰。
在RNA的5端,伸长和端修饰可以提高RNA稳定性和转录激活,同时可以加强RNA的抗衰老和诱导基因表达的效果。
而在RNA的3端,伸长和端修饰则有助于增加RNA的可行性,并可以抑制某些RNA的稳定性降低。
2. RNA后转录修饰的生物学意义RNA后转录修饰对于生物学过程中的一些基本功能起到了非常重要的作用。
2.1 RNA后转录修饰与细胞周期RNA后转录修饰的存在对于细胞周期的正常运作有着至关重要的作用。
例如,RNA甲基化可以影响许多细胞周期关键基因的表达,从而影响细胞的生长和增殖。
2.2 RNA后转录修饰与免疫反应RNA后转录修饰对于免疫反应的调节有着非常重要的作用。
第二节 RNA转录后得加工与修饰不论原核或真核生物得rRNAs都就是以更为复杂得初级转录本形式被合成得,然后再加工成为成熟得RNA分子。
然而绝大多数原核生物转录与翻译就是同时进行得,随着mRNA开始得DNA上合成,核蛋白体即附着在mRNA上并以其为模板进行蛋白质得合成,因此原核细胞得mRNA并无特殊得转录后加工过程,相反,真核生物转录与翻译在时间与空间上就是分天得,刚转录出来得mRNA就是分子很大得前体,即核内不均一RNA。
hnRNA分子中大约只有10%得部分转变成成熟得mRNA,其余部分将在转录后得加工过程中被降解掉。
(一)mRNA得加工修饰原核生物中转录生成得mRNA为多顺反子,即几个结构基因,利用共同得启动子与共同终止信号经转录生成一条mRNA,所以此mRNA分子编码几种不同得蛋白质。
例如乳糖操纵子上得Z、Y及A基因,转录生成得mRNA可翻译生成三种酶,即半乳糖苷酶,透过酶与乙酰基转移酶。
原核生物中没有核模,所以转录与翻译就是连续进行得,往往转录还未完成,翻译已经开始了,因此原核生物中转录生成得mRNA没有特殊得转录后加工修饰过程。
真核生物转录生成得mRNA为单顺反子,即一个mRNA分子只为一种蛋白质分子编码。
真核生物mRNA得加工修饰,主要包括对5’端与3’端得修饰以及对中间部分进行剪接。
1.在5’端加帽成熟得真核生物mRNA,其结构得5’端都有一个m7G-PPNmN结构,该结构被称为甲基鸟苷得帽子。
如图17-9所示。
鸟苷通过5’-5’焦磷酸键与初级转录物得5’端相连。
当鸟苷上第7位碳原子被甲基化形成m7G-PPNmN时,此时形成得帽子被称为“帽0”,如果附m7G-PPNmN外,这个核糖得第“2”号碳上也甲基化,形成m7G-PPNm,称为“帽1”,如果5’末端N1与N2中得两个核糖均甲基化,成为m7G-PPNmPNm 2,称为“帽2”。
从真核生物帽子结构形成得复杂可以瞧出,生物进化程度越高,其帽子结构越复杂。
RNA加工和调节基因表达随着生物学领域的不断深入,越来越多的科学家开始注重研究RNA分子在生物体内的作用。
RNA作为一种基因表达的重要调控元件,在转录和翻译过程中扮演着至关重要的角色。
RNA分子需要经过加工和修饰,才能正常发挥其功能。
本文将从RNA加工和调节基因表达的角度进行探讨。
RNA加工的过程RNA加工是指在转录过程中,通过剪接、修饰、剪切等步骤,将原始的RNA转录出来的前体RNA(pre-mRNA)分解成成熟的RNA(mRNA)。
这个过程中涉及到了许多RNA加工的蛋白质因子和RNA酶。
下面我们详细介绍一下RNA加工的过程。
一、剪接剪接是RNA加工的重要环节之一,即对pre-mRNA的剪接边界进行切割,把内含子切下并形成外显子。
这个过程中需要一些特殊的RNA酶和snRNP(小核仁核糖核蛋白复合体)。
snRNP可以组成一个复合物,叫做小核仁体(spliceosome),它能识别pre-mRNA上的内含子-外显子边界。
在识别完这个边界后,就会按照边界的位置把内含子切下,形成一个含有外显子的成熟RNA分子。
二、修饰修饰过程是RNA加工中另一个非常重要的环节。
修饰可以分为两类:核苷酸修饰和结构修饰。
核苷酸修饰是指在RNA链上,将某些核苷酸进行化学修饰,如在腺嘌呤核苷酸上添加m6A(N6-甲基腺嘌呤)。
这些化学修饰能够影响RNA分子的结构,从而改变它们的稳定性和功能。
结构修饰是指通过添加一些化合物或类似物来修饰RNA链的二级结构,如RNA分子上的N6-methyladenosine(m6A)修饰物,它能影响RNA分子的相互结合和调节基因表达。
三、剪切剪切是RNA加工过程的一种不同于剪接的加工方式,它主要发生在小RNA和pre-mRNA上。
过程中需要一个特殊的RNA酶叫做Dicer,它能识别和剪切一些形态不一的RNA前体分子。
剪切后的小RNA可以形成类似于microRNA或siRNA的分子,通过介导基因表达来发挥调控作用。
RNA修饰在RNA后转录修饰中的研究RNA能够传递基因信息,从而控制蛋白质的表达。
然而,随着对RNA的研究越来越深入,研究人员发现了许多在RNA级别上的后转录修饰,这些修饰是在RNA转录之后进行的。
其中最重要的是RNA修饰。
RNA修饰是一种在转录后修饰RNA的高度保守的生物化学过程,它能够改变RNA的结构和功能。
RNA修饰早在上世纪70年代就已经被发现。
到目前为止,已经确定了超过170种RNA修饰,其中最为重要的是N6-甲基腺苷酸(m6A)修饰、5-羟甲基脲嘧啶(hm5C)修饰和2’-O-甲基化修饰。
这些修饰在RNA的运输、翻译、分解和RNA的空间结构中都扮演着重要的角色。
m6A修饰是RNA中最常见的修饰,在mRNA、tRNA和rRNA中都能够发现它。
m6A修饰与gene regulation、转录、RNA降解等多种生物过程相关联。
近年来,研究者对m6A修饰的机制和作用进行了深入的研究,m6A修饰的作用越来越受到人们的重视。
当前,研究者已经通过多种技术手段对m6A修饰进行了定量和分析。
其中最常用的是抗体法。
抗体法利用与m6A特异性结合的抗体来检测和定量m6A修饰。
与m6A修饰不同,hm5C和2’-O-甲基化修饰对RNA的影响尚未完全被理解。
但是在最近的研究中,研究者发现hm5C修饰在RNA的结构和稳定性中扮演重要的角色。
2’-O-甲基化修饰则被发现与RNA的翻译和降解有关。
RNA修饰的发现和研究极大地促进了对RNA的研究,同时也为许多疾病的研究提供了新的思路。
例如,m6A修饰在某些肿瘤中有明显的异常表达,研究m6A修饰的异常表达和机制有助于深入了解肿瘤的发生机制。
总之,RNA修饰是RNA后转录修饰中非常重要的一部分。
研究者对RNA修饰的研究使我们更深入地了解RNA的功能和机制。
早期的研究主要集中在m6A修饰上,但现在研究者已经开始探索其他类型的RNA修饰,并通过研究RNA修饰来促进对各种生物过程的深入了解。
第二节RNA转录后的加工与修饰不论原核或真核生物的rRNAs都是以更为复杂的初级转录本形式被合成的,然后再加工成为成熟的RNA 分子。
然而绝大多数原核生物转录和翻译是同时进行的,随着mRNA开始的DNA上合成,核蛋白体即附着在mRNA上并以其为模板进行蛋白质的合成,因此原核细胞的mRNA并无特殊的转录后加工过程,相反,真核生物转录和翻译在时间和空间上是分天的,刚转录出来的mRNA是分子很大的前体,即核内不均一RNA。
hnRNA 分子中大约只有10%的部分转变成成熟的mRNA,其余部分将在转录后的加工过程中被降解掉。
(一)mRNA的加工修饰原核生物中转录生成的mRNA为多顺反子,即几个结构基因,利用共同的启动子和共同终止信号经转录生成一条mRNA,所以此mRNA分子编码几种不同的蛋白质。
例如乳糖操纵子上的Z、Y及A基因,转录生成的mRNA可翻译生成三种酶,即半乳糖苷酶,透过酶和乙酰基转移酶。
原核生物中没有核模,所以转录与翻译是连续进行的,往往转录还未完成,翻译已经开始了,因此原核生物中转录生成的mRNA没有特殊的转录后加工修饰过程。
真核生物转录生成的mRNA为单顺反子,即一个mRNA分子只为一种蛋白质分子编码。
真核生物mRNA的加工修饰,主要包括对5’端和3’端的修饰以及对中间部分进行剪接。
1.在5’端加帽成熟的真核生物mRNA,其结构的5’端都有一个m7G-PPNmN结构,该结构被称为甲基鸟苷的帽子。
如图17-9所示。
鸟苷通过5’-5’焦磷酸键与初级转录物的5’端相连。
当鸟苷上第7位碳原子被甲基化形成m7G-PPNmN时,此时形成的帽子被称为“帽0”,如果附m7G-PPNmN外,这个核糖的第“2”号碳上也甲基化,形成m7G-PPNm,称为“帽1”,如果5’末端N1和N2中的两个核糖均甲基化,成为m7G-PPNmPNm2,称为“帽2”。
从真核生物帽子结构形成的复杂可以看出,生物进化程度越高,其帽子结构越复杂。
图17-9 Post-transcriptional modification of mRNa showing the 7-methylguanosine cap and poly-Atail.真核生物mRNA 5’端帽子结构的重要性在于它是mRNa 做为翻译起始的必要的结构,对核糖体对mRNA 的识别提供了信号,这种帽子结构还可能增加mRNA的稳定性,保护mRNa 免遭5’外切核酸酶的攻击。
2.在3’端加尾大多数的真核mRNA 都有3’端的多聚尾巴(A),多聚(A)尾巴大约为200bp。
多聚(A)屠巴不是由DNA编码的,而是转录后在核内加上去的。
受polyA聚合酶催化,该酶能识别,mRNa 的游离3’-OH端,并加上约200个A残基。
近年来已知,在大多数真核基因的3’一端有一个AATAA序列,这个序列是mRNa 3’-端加polyA尾的信号。
靠核酸酶在此信号下游10-15碱基外切断磷酸二酯键,在polyA聚合酶催化下,在3’-OH上逐一引入100-200个A碱基。
关于polyA尾巴的功能问题尽管经过极其广泛的探索,但还不完全清楚。
有人推测polyA可能与mRNA从细胞核转送到细胞质有关,但是相当数量,的没有polyA屠巴的mRNA如组蛋白mRNA,也照样通过核膜进入细胞质。
还有人认为这种结构对真核mRNA的翻译效率具有某种作用,并能稳定mRNA 结构,保持一定的生物半衰期。
3.mRNA前体(hnRNA)的拼接原核生物的结构基因是连续编码序列,而真核生物基因往往是断裂基因,即编码一个蛋白质分子的核苷酸序列被多个插入片断所隔开,一个真核生物结构基因中内含子的数量,往往与这个基因的大小有关,例如胰岛素是一个很小的蛋白质,它结构基因只有两个内含子,而有些很大的蛋白质,它的结构基因中可以有几十个内含子。
经过复杂的过程后,切去内元,将有编码意义的核苷酸片段(Extron外元也叫外显子)连接起来(图17-10)。
图17-10 Primary polymerase 11transcript of a eukaryote gene showing (a)introns after capping and addition of polyA tail.(b)Excision of introns to form the mature mRNA is called splicing.真核生物的结构的基因中具有可表达活性的外显子,也含有无表达活性的内含子,但内含子序列下是无意义的,越来越多的实验证明有许多基因中的内含子参与基因表达调控,在转录时,外显子及内含子均转录到hnRNA中。
在细胞核中hnRNA进行剪接作用,首先在核酸内切酶作用下剪切掉内含子;然后在连接酶作用下,将外显子各部分连接起来,而变为成熟的mRNA,这就是剪接作用,也有少数基因的hnRNA不需进行剪接作用,例如α-干扰素基因,图17-11以卵清蛋白基因为例,介绍一个典型的转录及加工过程。
图17-11 卵清蛋白基因转录及加工过程图中外显示以1、2、3、4……表示,内含子以A、B、C、D…表示mRNA的拼接,需要在拼接部位有供拼接识别的保守性强的一致顺序,通过对100多种真核细胞基因的分析,发现外元和内元拼接部位部分碱基顺序有一定的规律(见表17-4)。
表17-4 含有内元的转录产物其拼接处的碱基顺序表中划线的碱基对拼接识别有重要作用,如将兔的β-珠蛋白的拼接部位的GT改为AT后,拼接反应即受到影响。
mRNA前体拼机制图17-12 The RNA splicing mechanism.RNA splicing is catalyzed by a spliceosome formed from the assembly of U1,U2,U5,and sn RNPs(shown as green circles )plus other components (not shown).After assembly of the spliceosome ,the reaction occures in two speps:in step 1the branch-point A nucleotide in the intron sequence,which is located colse to the 3'splice site ,attacks the 5'splice site and cleaves it;the cut 5'end of the intron sequence thereby becomes covalently linked to this A nucleotide,forming the branched nucleotide shown in Figure 8-55.In step 2 the 3'-OH end of the first exon sequence,which was created in the first step,adds to the beginning of the second exon sequence,cleqving the RNA molecule at the 3'splice site;the two exon sequences are thereby joined to each other and the intron sequence is released ad a ribosone.These splicing reactions occur im the nucleus and gengerate mRNa molecules from primary RNA transcripts (mRNA precursormolecules).mRNA拼接反应需要有核内小分子RNA参与它们与蛋白质形成的复合物称为小核糖核蛋白颗粒,SnRNA 分别被命名为U1,U2,U3,U4,U5,和U6RNA。
SnRNA中的U2RNA由与内元右端拼接部位附近的UACUAA顺序高度互补,形成一个环状结构,由特定的酶来识别切除该环状结构,完成拼接过程,如图17-12所示。
图17-13 Mechanim of mRNa splicing.Note that,for clarity,the process is shown in two stages;energy is not required for the process since transesterification reactions are involved.真核生物 mRNA前体在剪接过程中,还可以形成套索样的结构,在内含子序列中常有一个分支部位的腺苷酸残基,它的2’-OH可以自动攻击内含子5’端与外显子1连接的磷酸二酯键,切开了外噗子1,而腺苷酸原来已有3’,5’--磷酸二酯键相连的两个相邻的核苷酸残基,加上此3’,5’-磷酸二酯键连接后,在腺苷酸处出现了一个套索,已被切下的外显子1的3’-OH攻击内含子3’末端与外显子2之间的3’,5’-磷酸二酯键,键断裂后,内含子以套索的形式被节下来,此时外显子1和外显子2可以连接起来(图17-13)。
不论拼接过程如何,拼接必须极为精确,否则会导致遗传信息传递障碍,合成的蛋白质可能丧失其正常的功能。
我国南方广大地区是β-地中海贫血的高发区,这是由于β-珠蛋白链的合成受到部分或完全抑制所引起的一种血红蛋白病。
实验表明β-珠蛋白基因元1中核苷酸的点突变改变了正常拼接部位的碱基顺序,结果造成错误部位的拼接。
加工成熟的mRNA虽能翻译,但产物不是正常的β-珠蛋白,结果引起血红蛋白级结构和功能的改变。
(二)rRNA转录后加工原核生物rRNA转录后加工,包括以下几方面:①rRNA前体被大肠杆菌RNaseⅢ,RNaseE等剪切成一定链长的rRNA分子;②rRNA在修饰酶催化下进行碱基修饰;③rRNA与蛋白质结合形成核糖体的大、小亚基(见图17-14)图17-14 大肠杆菌rRNA前体的加工真核生物rRNA前体比原核生物大,哺乳动物的初级转录产物为45s,低等真核生物的rRNA前体为38s,真核生物5sRNA前体独立于其他三种rRNA的基因转录(图17-15)。
图17-15 真核生物rRNA前体的加工真核生物rRNA前体中含有插入顺序,rRNA前体要形成成熟的rRNA,需要经过拼接反应。
例如,四膜虫的rRNA前体的拼接是一种无酶催化的自动拼接过程。
四膜虫基因组内,26srRNA编码的区域内有413bp 的插入顺序。
该插放序列可以不消耗能量从rRNA前体中被除掉。