转录与后加工
- 格式:doc
- 大小:29.50 KB
- 文档页数:2


转录后加工名词解释
转录后加工是指在基因组中进行转录的过程后,对转录产物(RNA分子)进行进一步的修饰和加工的过程。
转录是指在DNA模板上合成RNA分子的过程,而转录后加工则是在RNA分子合成完成后对其进行一系列的修饰和处理。
转录后加工的目的是为了产生成熟的RNA分子,使其能够发挥特定的功能。
在转录后加工过程中,RNA分子经历剪接、修饰和运输等多个步骤,以形成成熟的RNA分子。
剪接是转录后加工中最重要的步骤之一。
在剪接过程中,RNA 分子的内含子(非编码区域)会被剪除,而外显子(编码区域)则会被保留下来。
这样一来,通过剪接,一个基因可以产生多个不同的成熟RNA分子,从而扩大了基因的功能和多样性。
除了剪接,转录后加工还包括其他的修饰过程。
例如,RNA分子可能会经历5'端帽子的添加和3'端的聚腺苷酸尾巴的加入,这些修饰可以保护RNA分子免受降解,并有助于其在细胞内的稳定性和转运过程中的识别。
此外,转录后加工还可以包括RNA编辑、互补RNA合成和核糖体扫描等过程。
RNA编辑是指在转录后,RNA分子中的碱基序列可以发生改变,从而导致RNA分子的信息内容发生变化。
互补RNA合成是指利用RNA分子作为模板合成互补的DNA分子。
核糖体扫描是指RNA分子被核糖体识别并翻译成蛋白质的过程。
总的来说,转录后加工是一系列对转录产物进行修饰和加工的过程,通过这些过程,RNA分子可以获得特定的功能和稳定性,从而发挥其在细胞中的重要作用。



rna转录后加工名词解释
RNA转录后加工是指对在转录过程新合成的RNA的前体分子,进行进一步的加工修饰,从而使其成为具有生物学活性的、成熟的RNA 分子的过程,主要包括剪接、化学修饰等方式。
1.可变剪切:通过不同的剪切方式使得同一个基因可以产生多个不同的成熟mRNA,最终产生不同的蛋白质,从而使转录本和蛋白质结构与功能具有多样性。
2.RNA编辑:属于修饰的一种,是指转录后的RNA在编码区发生碱基的加入、丢失或转换等现象,可以在RNA水平上增加一些原来DNA模板上没有编码的碱基,从而扩充遗传信息。
因此,经过剪切或者修饰等加工,遗传信息的含量以及多样性大大增加。
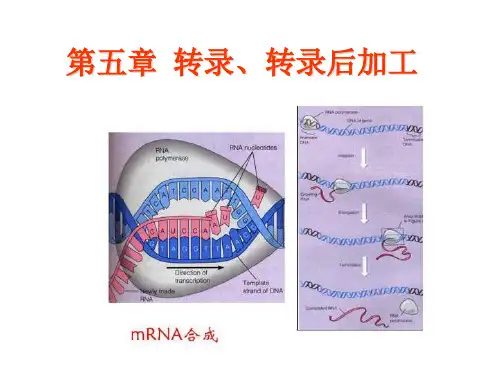


①中心法则:是指遗传信息从DNA传递给RNA,再从RNA传递给蛋白质,即完成遗传信息的转录和翻译的过程。
也可以从DNA传递给DNA,即完成DNA的复制过程。
这是所有有细胞结构的生物所遵循的法则。
在某些病毒中的RNA自我复制(如烟草花叶病毒等)和在某些病毒中能以RNA为模板逆转录成DNA的过程(某些致癌病毒)是对中心法则的补充。
②基因表达:指细胞在生命过程中,把储存在DNA顺序中遗传信息经过转录和翻译,转变成具有生物活性的蛋白质分子。
(差别基因表达(differential gene expression):指细胞分化过程中,奢侈基因按一定顺序表达,表达的基因数约占基因总数的5%~10%。
也就是说,某些特定奢侈基因表达的结果生成一种类型的分化细胞,另一组奢侈基因表达的结果导致出现另一类型的分化细胞,这就是基因的差别表达。
其本质是开放某些基因,关闭某些基因,导致细胞的分化。
)③编码链:双链DNA中,不能进行转录的那一条DNA链,该链的核苷酸序列与转录生成的RNA的序列一致(在RNA中是以U取代了DNA中的T),又称有义链(sense strand)。
④模板链(template strand):可作为模板转录为RNA的那条链,该链与转录的RNA碱基互补(A-U,G-C)。
在转录过程中,RNA聚合酶与模板链结合,并沿着模板链的3'→5';方向移动,按照5'→3'方向催化RNA 的合成。
又称为反义链(antisence strand)。
⑤启动子(promoter):指RNA聚合酶特异性识别和结合的DNA序列。
启动子是基因(gene)的一个组成部分,控制基因表达(转录)的起始时间和表达的程度。
但是启动子本身并不控制基因活动,而是通过与称为转录(transcription)因子的这种蛋白质(proteins)结合而控制基因活动的。
一般分为广谱表达型启动子、组织特异性启动子、肿瘤特异性启动子等多种形式。


RNA的转录与转录后加工一、名词解释1、基因诊断2、RFLP3、启动子 4. 信号肽 5. 核受体 6.hnRNA7、基因治疗8、反义RNA9、核酶10、三链DNA11、SSCP12、管家基因13. 增强子14. 基础转录装置18. 重叠基因19.假基因20.RNA干扰21.酵母双杂交22.转录因子23.转录因子的结构24.衰减子25.内含子27.弱化子28.魔斑29.上游启动子元件30.DNA探针31.SD sequence 32.Ribozyme 33.Terminator二、填空题1、转录是以DNA一条链为模板的RNA的酶促合成。
我们把模板链称为-- --------。
2、数个生化反应可由----- -----------催化,这种具有催化功能的RNA可以剪切自身或其它的RNA分子,或者完成连接或自身剪接反应。
3.RNA酶的剪切分为()、()两种类型。
4.原核生物中有三种起始因子分别是()、()和()。
5.hnRNA与mRNA之间的差别主要有两点:(),()。
6.mRNA在转录开始后不久就与结合,形成颗粒,这种颗粒排列于mRNA 分子上,呈串珠状,就像核小体一样。
7、原始转录物的一些序列被_____________,叫做RNA编辑。
8. 真核生物mRNA的5'-帽子结构是_______,其3'末端有________结构。
9. 原核生物DNA指导的RNA聚合酶的核心酶的组成是___________.10. 真核生物RNA聚合酶III负责转录_________.11. 在转录过程中RNA聚合酶全酶的σ因子负责__________,核心酶负责________.三、选择题1、RNA合成的底物是------ ---------。
A dA TP, dTTP , dGTP , d CTPB A TP, TTP , GTP , CTPC A TP ,GTP, CTP,UTPD GTP, CTP,UTP,TTP2.模板DNA的碱基序列是3′—TGCAGT—5′,其转录出RNA碱基序列是:A.5′—AGGUCA—3′B.5′—ACGUCA—3′C.5′—UCGUCU—3′D.5′—ACGTCA—3′E.5′—ACGUGT—3′3、转录终止必需。
RNA转录与转录后加⼯第7章 RNA转录与转录后加⼯1 本章主要内容1)转录的基本概念2)⼤肠杆菌RNA聚合酶及其转录3)真核⽣物的RNA聚合酶及其转录4)RNA的转录后加⼯和反转录2 教学⽬的和要求通过本章学习,掌握转录的基本概念,原核转录的主要参与者(RNA聚合酶和启动⼦)以及原核转录的过程(起始、延伸和终⽌)。
1)掌握真核转录的三种主要RNA聚合酶、所转录的基因类型和参与转录过程各种因⼦等。
2)了解不同前体RNA的加⼯机制。
3)了解反转录的特点3 重点难点1) 转录2) ⼤肠杆菌RNA聚合酶、原核转录的过程3) 真核⽣物的RNA聚合酶、真核转录过程、转录因⼦4) RNA的转录后加⼯、反转录4 教学⽅法与⼿段讲授与交流互动相结合,采⽤多媒体教学。
5 授课内容1)RNA转录概述2)细菌基因的转录3)真核⽣物的转录4)RNA的转录后加⼯5) RNA的反转录第⼀节 RNA转录概述⼀、信使的发现1955年Brachet⽤洋葱根尖和变形⾍进⾏实验:–若加⼊RNA酶,则蛋⽩质合成就停⽌;–若再加⼊来⾃酵母的RNA,⼜可合成蛋⽩质。
这表明什么?同年Goldstein和Plaut⽤同位素标记变形⾍RNA前体——发现标记的RNA在核内。
标记追踪实验:经过⼀段时间⼜发现被标记的RNA在细胞质中,这表明什么?1956年E. Volkin和L.Astrachan:⽤同位素脉冲⼀追踪标记表明T2噬菌体新合成的RNA的碱基⽐和⾃⾝的DNA碱基⽐相似,⽽和细菌的碱基⽐不同。
T2感染细菌时注⼊的是DNA,⽽在细胞⾥合成的是RNA。
这表明什么?最令⼈信服的证据是Hall.B.D和Spiegeman,S的DNA-RNA的杂交实验:将T2噬菌体感染E.coli后产⽣的RNA分离出来,分别与T2和E.coli的DNA进⾏分⼦杂交。
结果这种RNA只能和T2的DNA形“杂种”链,⽽不能和E.coli的DNA进⾏杂交。
Jacob和Monod预⾔:(1)这种“ 信使”应是⼀个多核苷酸;(2)其分⼦平均不⼩于5 105bp,⾜以携带⼀个基因的遗传信息;(3)它们⾄少是暂时连在核糖体上;(4)其碱基组成反映了DNA的序列;(5)它们能⾼速更新。
一、原核生物(一)核糖体RNA:大肠杆菌共有7个核糖体RNA的转录单位,每个转录单位由16S、23S、5SRNA和若干转运RNA基因组成。
16S和23S之间常由转运RNA隔开。
转录产物在RNA酶III的作用下裂解产生核糖体RNA的前体P16和P23,再由相应成熟酶加工切除附加序列。
前体加工时还进行甲基化,产生修饰成分,特别是a-甲基核苷。
N4,2’-O二甲基胞苷(m4Cm)是16S核糖体RNA特有成分。
5S核糖体RNA一般无修饰成分。
(二)转运RNA:有60个基因,其加工包括:1.内切酶在两端切断,大肠杆菌RNA酶P是5’成熟酶2.外切酶从3’修剪,除去附加顺序。
RNA酶D是3’成熟酶3.3’端加上CCAOH,由转运RNA核苷酰转移酶催化,某些转运RNA已有,切除附加序列后即露出。
4.核苷的修饰:修饰成分包括甲基化碱基和假尿苷,修饰酶具有高度特异性。
甲基化对碱基和序列都有严格要求,一般以S-腺苷甲硫氨酸为甲基供体。
(三)信使RNA:细菌多数不用加工,转录与翻译是偶联的。
也有少数多顺反子信使RNA必须由内切酶切成较小的单位,然后翻译。
如核糖体大亚基蛋白与RNA聚合酶的b亚基基因组成混合操纵子,转录后需经RNA酶III切开,各自翻译。
因为RNA聚合酶的合成水平低得多,切开有利于各自的翻译调控。
较长的RNA会产生高级结构,不利于翻译,切开可改变其结构,从而影响其功能。
二、真核生物(一)核糖体RNA:基因拷贝数多,在几十到几千之间。
基因成簇排列在一起,由RNA聚合酶I转录生成一个较长的前体,哺乳动物为45S。
核仁是其转录、加工和装配成核糖体的场所。
RNA酶III等核酸内切酶在加工中起重要作用。
5SRNA基因也是成簇排列的,由RNA聚合酶III转录,经加工参与构成大亚基。
核糖体RNA可被甲基化,主要在核苷2’羟基,比原核生物甲基化程度高。
多数核糖体RNA没有内含子,有些有内含子但不转录。
(二)转运RNA:由RNA聚合酶III转录,加工与原核相似,但3’端的CCA 都是后加的,还有2’-O-甲基核糖。
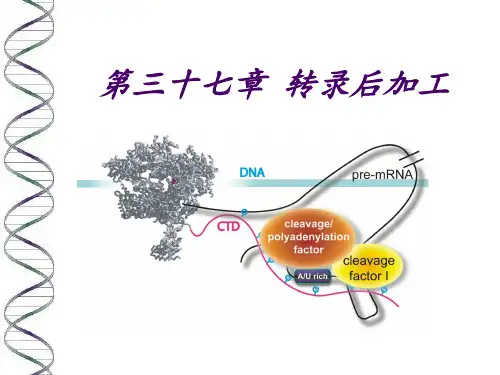
第十四章转录后加工和调节
********真核生物mRNA前体加工
转录起始于第1个外显子(exon)的第一个核苷酸,转录开始不久转录产物的5´端就被加上7-甲基鸟苷酸帽子(Cap)。
转录终止于最后1个外显子3´端下游大约0.5~2.OKb范围内多个可能位点中的1个。
核酸内切酶在polyA位点切除多余序列,并在polyA聚合酶的作用下加上长100~250个A。
接着,通过RNA剪接(splicing)将内含子(intron)切除,并将外显子连接起来。
最后,将成熟的mRNA分子输送到细胞质中。
以上过程称为mRNA前体加工(pre-mRNA processing)。
当RNA聚合酶Ⅱ的转录产物刚合成大约30个核苷酸时,其5´端就加上了1个甲基化鸟苷酸(m7G),甲基供体是S—腺苷蛋氨酸。
mRNA 5´端帽子结构的主要功能有;①在蛋白质合成起始中的重要作用类似于原核生物 mRNA的SD 序列,供核糖体小亚基(40S)识别与结合。
②保护合成中的转录产物免受核酸外切酶的降解。
③在成熟的
mRNA以外,动物所有的mRNA 3´端都有polyA尾。
这些A是初始转录产物经过核酸内切
mRNA的polyA尾上游10—35个核苷酸处都含有序列AAUAAA,在切割位点下游大约50个核苷酸以内还存在第二个加尾信号,其序列特征是富含G/U或U。
PolyA位点切割后,多聚腺苷酸化分为两个阶段进行。
前12个左右的A的多聚腺苷酸化的速度比较慢,而后面则很快,迅速加到200~250个A。
后者需要结合若干含有RNP motif的polyA结合蛋白Ⅱ(PABⅡ)。
------外显子—内含子交界处存在短共有序列 ,其中,出现机率为100%的只有pre-mRNA内含子5´端的GU和3´端的AG,这就是所谓剪接的GU—AG规则。
------ 六种富含U的snRNA大量存在于哺乳动物细胞核中,命名为U1~U6,它们参与RNA剪接。
这些snRNA和6~10种蛋白因子相结合,形成核内RNP小颗粒 (snRNP)。
--------反式剪接(trans—splicing)
产生于两个不同的RNA分子之间发生的剪接作用,这个过程叫作反式剪接。
高等真核生物的转录单位分为简单和复合两种类型。
简单转录单位只有1个polyA位点,其RNA剪
接方式也只有1种,只能产生mRNA 分子,这意味着1
复合转录单位的转录后加工,是高等真核生物基因转录后调节的1种重要途径。
(二)选择性剪接
高等真核生物复合转录单位的。
剪接过程中发生了外显子跳跃(exon skip),使得某些外显子在成熟的mRNA分子中不复存在, 除了外显子跳跃以外,有的选择性剪接通过包含或排除终止密码子来控制功能蛋白的表达。
---------有的复合转录单位含有多个polyA位点,通过选择性调节初始转录产物3´端的切割位点,以改变表达产物C端的氨基酸顺序,产生长短不同的多肽链,这一过程叫作polyA位点选择(polyAsitechoice),或选择性多聚腺苷酸化(alternative polyadenylation)。
,核仁小RNA(snoRNA,即small nucleolar RNA)可能催化pre—rRNA的切割加工,snoRNA和蛋白因子结合形成snoRNP,再与pre—rRNA结合。
snoRNA的还可能在线粒体DNA复制时参与合成RNA引物,它的功能有助于协调细胞生长(或分裂)与线粒体复制的关系。
真核基因内含子主要分为四种类型:①核内含子②I类内含子(Group I)。
⑧Ⅱ类内含子(Group Ⅱ)。
④tRNA基因内含子。
I类内含子具有两个共同特点;①自我剪接能力(self-splicing)。
②特殊的二级结构,包括9个茎环。
rRNA前体分子中,但这种内含子不如I类内含子普遍。
------核酶(ribozyme)泛指具有催化功能的各种RNA分子。
Rnase P,它在tRNA前体加工中起重要作用。
像核酸内切酶一样。
拟病毒(virusoid)和类病毒(viroid)的RNA分子在局部形成特殊的锤头结构(hammer head),具有自我切割能力(autocleavage)。
核糖体大亚基的rRNA具有肽酰转移酶的活性。
指导RNA(gRNA)在RNA编辑中也具有催化活性,并提供插入的核苷酸单位U。
--------- 所谓RNA编辑(editing),是在RNA水平上改变遗传信息的加工过程,导致成熟的RNA (主要是mRNA)编码序列和它的转录模板DNA序列之间不相匹配。
RNA编辑加工的方式有碱基插入、缺失和取代等。
内含子序列参与哺乳动物RNA编辑过程,RNA编辑加工过程可能出现于RNA剪接加工之前。
锥虫线粒体的RNA编辑其主要形式是插入或缺失U,在锥虫线粒体基因组中散布着一些独立的转录单位,它们编码1种小分子指导RNA(guide RNA,即gRNA)。
----- gRNA的5´端和未经编辑的mRNA的一小段锚定序列(anchor)互补, gRNA的3´端则作为编辑加工的模板,决定插入或去除U的部位和数目。
gRNA序列和经过编辑的 RNA正好互补,
polyA尾的主要功能:①是成熟的mRNA的标志之一。
只有成熟的mRNA才能从核内输送到胞质。
②影响mRNA在核内及胞质中的稳定性。
③在消耗能量和前体进行翻译之前,polyA尾和5´端帽子结构是核糖体判断mRNA是否完整的识别信号。
某些mRNA(如逆转录病毒基因组)的特征性分子结构可能形成再编码信号(recoding signal),使正常的翻译过程中止,并发生翻译移码(translationalframeshift),导致1个mRNA分子合成一种以上的蛋白。
此外,终止密码子周围特异的核苷酸序列还可能造成终止密码子渗漏 (1eaky),以至在多肽链末端插入若干额外的氨基酸。
再编码是转录后基因表达调控的一种方式。
除了翻译移码和天然校正(利用校正tRNA)以外,蛋白质合成支路也是再编码途径之一。
--------核糖体能够跳过一个终止密码子继续合成。
在多肽链合成过程中核糖体可以通过支路(非正常途径)移位,故称之为蛋白质合成支路或核糖体跳跃(ribosome jumping)或tRNA跳跃(hopping)。