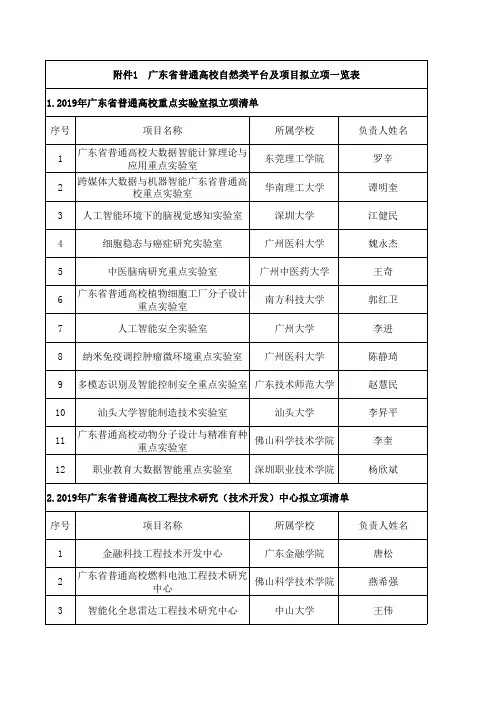
广东省普通高校自然类平台及项目拟立项一览表
- 格式:pdf
- 大小:249.92 KB
- 文档页数:12

大学生创新课题关于公布2009-2010学年学生课外科技学术活动立项项目及2010年广东省“大学生创新实验计划”项目的通知各学院、附属医院,教辅机构,根据《关于做好2009-2010学年学生课外科技学术活动课题申报及开展2010年广东省“大学生创新实验计划”项目申报的通知》(校教字[2010]395)号文件要求,由学生自主申请,各学院审核推荐,共有38项理科项目,10项文科项目被列入2010年广东省“大学生创新实验计划”项目,学校专项资金资助理科项目2000元,文科项目1000元,资助金额共计86000元。
共150个项目被批准为学生课外科技学术活动项目立项,其中自然科学类项目83项,每项资助800元,科技发明类项目20项,每项资助800元,哲学社会科学类项目47项,每项资助500元,资助金额共计105900元。
现予以公布,见附件,。
为了保证学生课外科研学术活动的顺利实施,并取得实质性成果,现就项目管理的有关事宜通知如下,一、广东省大学生创新实验项目1、广东省“大学生创新实验计划”项目开展时间从2010年6月30日开始计算。
大学生创新实验项目研究时间跨度为1-2年。
学校对项目进展进行跟踪管理,定期或不定期查阅实验工作记录、实施中期检查,查阅学生实验原始数据、实验报告和文献综述。
学校每年12月30日前向学校递交创新实验项目实施情况,填写中期检查报告,同结题验收书,。
学校根据实施进展情况进行总结并向教育厅报告项目进展情况。
2、项目结束后,由项目负责人提出结项申请,经学校教改项目管理部门组织专家组进行结项论证,省教育厅视学校提交结项材料情况或组织专家进行验收。
具体事项按有关规定执行,未在规定期限内完成的项目将予以公布或撤销。
评审结果报省教育厅备案。
3、根据《南方医科大学大学生创新性实验计划项目管理办法,试行,》和《南方医科大学大学生创新性实验计划项目实施方案,试行,》文件要求,请按《广东省大学生创新实验项目任务书》,填报一式四份于12月15日前报学校教务处综合计划科。


年广东高校省级重点平台和重大科研项目要求—————-——————---—————————————————作者:--—————--—-——-———————--———————-- 日期:附件12017年广东高校省级重点平台和重大科研项目认定申报要求一、重点建设平台只接受历年来获省教育厅立项的各类科研创新平台总数不超过6个的本科高校申报。
已通过省教育厅组织的验收,且已成功升格为省部级以上科研创新平台的,可不计入统计范围.(一)广东省普通高校重点实验室1。
学科建设基础好。
实验室所在学科应为省部级以上重点学科,原则上应具有硕士以上学位授予权(新兴学科、交叉学科或具有明显地方特色的学科除外)。
2。
研究方向和目标明确。
契合广东高校学科特点和重点学科布局,符合广东经济、社会发展的重大需求,研究领域属优先或重点发展方向,所从事的研究工作在本学科领域达到国内先进行列或较高水平,在国内同行中具备一定的优势,特色明显.具备承担国家或省部级重大、重点科研项目和培养高层次人才的能力,能积极开展国际合作研究与学术交流。
3。
学术团队结构合理.在所从事的研究领域内有知名度较高的学术带头人、优秀中青年学术骨干和团结协作、管理能力较强的领导班子;研究团队学术水平高、年龄和知识结构合理,研究人员中具有高级职称(含正高、副高)的人员在10名以上,实验室近5年承担省部级以上科研项目10项、并获得项目经费500万元以上;有良好的科研传统和学术氛围。
4。
科研实验条件良好。
实验室面积原则上不低于2000平方米,并相对集中;比较先进的、技术状态良好仪器设备原值不低于800万元(部分纯基础学科除外)。
有稳定的管理、技术人员队伍及比较健全的管理制度。
5.支持保障有力。
依托学校能保障实验室建设的经费投入,有必要的技术支撑、后勤保障、国内外合作与交流的条件。
(二)广东省普通高校工程技术研究(开发)中心1.工程研究中心应具备以下条件:①依托学校的优势学科专业,具有相关支撑学科、技术的系统集成条件,在某一技术领域有坚实的工程技术研发和成果转化的工作基础、特色和业绩.近三年来已与2个或以上企业合作,成功进行工程技术研发和成果转化2项或以上。

广东培训自然科学基金项目立项选题及研究方法全文共四篇示例,供读者参考第一篇示例:广东是中国南方的一个重要省份,也是我国经济最发达的地区之一。
在广东,为了促进科技创新和提升自然科学研究水平,广东省教育厅设立了广东培训自然科学基金项目,用于支持广东省内各高校和科研机构的自然科学研究项目。
立项选题是一个科研项目从立项到结题的全过程中最为关键的一环。
选择一个合适的研究选题不仅意味着项目的成功与否,更代表着研究者在这一领域的学术水平和科研能力。
在广东培训自然科学基金项目中,研究者需要遵循一定的评审标准和程序来选择合适的研究选题。
研究选题必须符合广东培训自然科学基金项目的研究方向和目标。
广东培训自然科学基金项目通常会设定一些研究方向和重点领域,研究者需要根据这些方向和领域来选择自己的研究选题。
广东培训自然科学基金可能会重点支持生物医学、新材料、清洁能源等方向的研究项目,研究者就需要在这些领域中选择一个能够符合基金项目要求的研究选题。
研究选题必须具有一定的新颖性和前瞻性。
广东培训自然科学基金项目鼓励研究者在科研领域中寻找研究热点和前沿问题,提出有创新性的研究选题。
在选择研究选题时,研究者要结合自己的科研领域和专长,找到一个能够在该领域中有所突破和贡献的研究课题。
研究选题必须具有一定的实用性和推广性。
广东培训自然科学基金项目是为了促进科研成果的转化和应用,因此研究选题必须具有一定的实用性和推广性。
研究者在选择研究选题时要考虑到该研究成果是否能够为社会和产业发展带来实际利益,是否具有一定的推广和应用前景。
在确定了研究选题后,研究者就需要设计合适的研究方法和方案来开展研究工作。
在广东培训自然科学基金项目中,研究方法的选择对于研究项目的成功至关重要。
研究者需要根据自己的研究选题和研究目标来确定合适的研究方法,以确保研究工作能够顺利进行并取得预期的成果。
第二篇示例:广东是我国经济最为发达的省份之一,广东的科技实力和创新能力一直备受关注。

广东省自然科学基金项目年度综合报告
(年度)
填报单位(盖章):
填报人(签章):
单位负责人(签章):
填报日期:
联系电话与传真:
E —mail 地址:
广东省科学技术厅基础研究处
2001年制
填表说明
一、每年12月15日前各单位将此文字材料与电子报表上报广
东省科学技术厅基础研究处;
二、上报材料应包括如下内容:(表格附后)
1、本年度在研项目、结题项目、跟踪项目清单;
2、本年度在研项目情况汇总表;
3、本年度结题项目情况汇总表;
4、本年度跟踪项目情况汇总表;
5、本年度重要在研项目、结题项目、跟踪项目简介;
6、本年度基金项目管理情况(经验、问题、建议);
7、本单位学科建设情况;
8、本单位重点实验室建设情况和承担在研项目、跟踪项
目的基本情况。
三、各类统计数据必须真实,必须有广东省基金资助的标注,
数据的审核请各单位严格把关;
四、年度统计时间为:上年度12月—本年度11月;
附:一、本年度在研项目、结题项目、跟踪项目清单表栏内容:
四、年度跟踪项目进展情况汇总表应跟踪:项,此处统计跟踪:项。

广东省自然科学基金自由申请项目重点博士启动申请书项目名称:多巴胺受体对吗啡诱导的信号传导和基因表达的调控作用申请者:张璐所在单位:南方医科大学(原第一军医大学)基础部邮政编码:510515通讯地址:广州同和南方医科大学申请者电话: 020-6传真:电子邮件:申请日期:2005-3-5广东省自然科学基金管理委员会二○○一年制填报说明一、填写申请书前,请先查阅广东省自然科学基金有关项目申请办法及规定。
申请书各项内容,应实事求是,逐条认真填写。
表达要明确、严谨,字迹清晰易辨。
外来语要同时用原文和中文表达。
第一次出现的缩写词,须注出全称。
二、申请书请用A4复印纸,于左侧装订成册。
第三页起各栏空格不够时,请自行加页。
一式八份(至少一份为原件),由所在单位审查签署意见后,按申报通知送广东省自然科学基金管理委员会办公室。
三、封面右上角“顺序号”由各单位根据广东省自然科学基金管理委员会办公室的要求填写;“项目类别”栏由申请者填写, 申请“省自然科学基金自由申请项目”此栏为“A”, “省自然科学基金重点项目”为“C”,“省自然科学基金博士启动项目”为“D”。
简表填写要求一、简表内容必须逐项认真填写,采用国家公布的标准简化汉字。
简表中所有代码,以国家自然科学基金规定的代码为准填写。
二、凡选择性栏目,将相应提示符A、B等之一填入该栏的右下角。
三、部分栏目填写要求:项目名称──应确切反映研究内容和范围,最多不超过25个汉字(包括标点符号)。
基础研究──指以认识自然现象、探索自然规律为目的,不直接考虑应用目标的研究活动。
应用基础研究──指有广泛应用前景,但以获取新原理、新知识、新方法为主要目的的研究。
申报学科──申请项目所属的最基础学科。
如涉及多学科可填写两个,先填写主学科。
申请金额──以万元为单位,用阿拉伯数字表示。
研究期限──研究期限一般从申请的次年1月算起。
终止时间为完成年度的12月。
所用实验室──系指研究项目将利用的实验室。
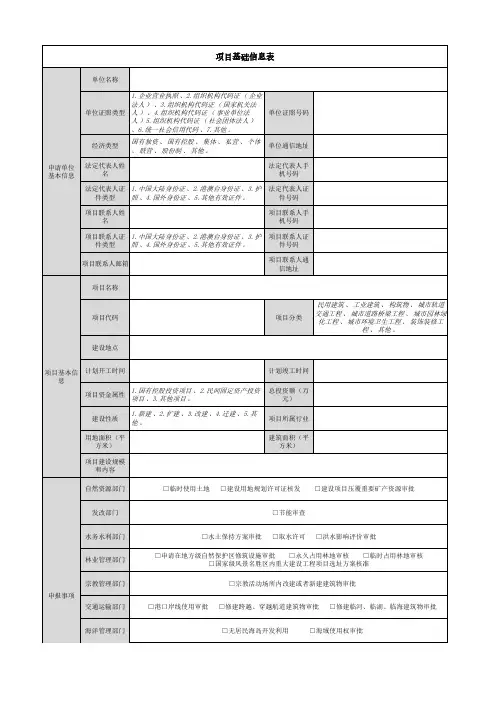
取水许可水源类型□普通□地热水□矿泉水□其他年申请取水量(万m3)取水地点地点名称: ______省(自治区) ______县(市、区) ______ 乡(镇)位置坐标:东经 ______ ° ______ ′ ______ ″,北纬 ______ ° ______ ′______ ″(坐标测量方法:____________ )取水方式□单井□井群□自流□单井□井群□自流□单井□井群□自流□单井□井群□自流计量方式最大取水流量(m³/s)日最大取水量(m3/d)申请取水期限申请在地方级自然保护区修筑设施审批/永久、临时占用林地审核使用林地性质使用期限应缴森林植被恢复费(元)使用林地类型总计防护林林地特用林林地用材林林地经济林林地薪炭林林地苗圃地其他林地面积(公顷)计国有集体蓄积(立方米)计国有集体林地保护等级国家级公益林地自然保护区林地级别面积(公顷)级别面积(公顷)级别面积(公顷)Ⅰ一国家级Ⅱ二省级Ⅲ三Ⅳ森林公园林地湿地公园林地风景名胜区林地级别面积(公顷)级别面积(公顷)级别面积(公顷)国家级国家级国家级省级省级省级重点保护野生动物栖息地重点保护植物及生境古树名木及保护范围使用地方公益林地面积(公顷)省级公益林其他公益林在草原上修建直接为草原保护和畜牧业生产服务的工程设施使用草原审批草原征占用类别□征用使用□临时占用□修建服务用工程设施拟征占用草原的时限拟征占用草原具体用途拟征占用草原的基本情地点______省(区、市)______市(地、州、盟)______ 县(市、旗) ______乡(镇) 村权属草原所有者或使用者状况草原承包经营权者证书编号面积总面积公顷(折合 亩)天然草原公顷(折合______亩)人工草地公顷(折合______亩)其中基本草原公顷(折合______亩)补偿及征收费用合计(万元)草地补偿费牧草补偿费安置补助费植被恢复费涉及国家安全事项的建设项目审批项目外文名称用地批号规划许可批号土地权属建筑高度层数。

㊀山东农业科学㊀2023ꎬ55(10):15~21ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.10.003收稿日期:2022-12-20基金项目:广东省基础与应用基础研究基金项目(2023A1515010336ꎬ2021A1515011236)ꎻ广东省普通高校重点领域专项(2022ZDZX4047)ꎻ韶关学院重点项目(SZ2022KJ05)ꎻ国家自然科学基金项目(31901537)ꎻ韶关学院博士启动基金项目(99000615)ꎻ国家级大学生创新创业训练计划项目(202310576009)作者简介:曾坚(1987 )ꎬ男ꎬ博士ꎬ副教授ꎬ从事植物基因功能研究ꎮE-mail:zengjian@sgu.edu.cn通信作者:胡伟(1982 )ꎬ男ꎬ博士ꎬ研究员ꎬ从事植物基因分子生物学研究ꎮE-mail:huwei2010916@126.com香蕉中ABA醛氧化酶基因家族的鉴定及其表达分析曾坚1ꎬ周洁薇1ꎬ王舒婷1ꎬ胡伟2(1.韶关学院广东省粤北食药资源利用与保护重点实验室/英东生物与农业学院ꎬ广东韶关㊀512005ꎻ2.中国热带农业科学院热带生物技术研究所ꎬ海南海口㊀571101)㊀㊀摘要:ABA醛氧化酶(abscisicaldehydeoxidaseꎬAAO)在植物生长发育和对生物/非生物胁迫的响应中起着重要作用ꎮ本研究从香蕉基因组中鉴定出3个MaAAOs基因ꎬ并对其遗传进化关系㊁结构域㊁理化性质及在不同组织㊁不同果实发育阶段和逆境处理下的表达情况进行了分析ꎮ结果表明ꎬ这3个MaAAOs分为两个亚族ꎬ相同亚族的基因呈现出类似的结构域组成和基因结构ꎬ与系统发育树的构建结果相一致ꎮMaAAO1基因在果实的发育和成熟阶段都表现出显著的高表达水平ꎬ暗示着其在香蕉果实发育过程中可能发挥着重要功能ꎮ另外ꎬMaAAO1和MaAAO2基因可能对渗透胁迫有响应ꎬMaAAO2基因还可能对Foc4病菌的侵染有响应ꎮ基于以上结果推断MaAAOs基因在香蕉的生长发育调控以及逆境响应中具有重要作用ꎮ该结果不仅丰富了我们对MaAAOs基因家族的认识ꎬ也为进一步揭示其在香蕉生长发育和应激响应机制方面的功能提供了线索ꎮ关键词:香蕉ꎻABA醛氧化酶(AA0)ꎻ基因鉴定ꎻ基因表达中图分类号:S668.1ꎻQ781㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)10-0015-07IdentificationandExpressionAnalysisofAAOGenesinBananaZengJian1ꎬZhouJiewei1ꎬWangShuting1ꎬHuWei2(1.GuangdongProvincialKeyLaboratoryofUtilizationandConservationofFoodandMedicinalResourcesinNorthernRegion/HenryFokSchoolofBiologyandAgricultureꎬShaoguanUniversityꎬShaoguan512005ꎬChinaꎻ2.InstituteofTropicalBioscienceandBiotechnologyꎬChineseAcademyofTropicalAgriculturalSciencesꎬHaikou571101ꎬChina)Abstract㊀Abscisicaldehydeoxidase(AAO)playsimportantrolesinplantgrowthanddevelopmentandresponsetobioticandabioticstresses.InthisstudyꎬthreeAAOgenes(MaAAOs)wereidentifiedfrombananagenomeꎬandtheirgeneticevolutionaryrelationshipꎬstructuraldomainꎬphysicalandchemicalpropertiesꎬandexpressionpatternindifferenttissuesꎬatdifferentfruitdevelopmentstagesandunderdifferentstresstreatmentswereanalyzed.TheresultsshowedthatthethreeMaAAOgenescouldbedividedintotwosubgroupsꎬandthegenesinthesamegroupexhibitedsimilarcombinationsofstructuraldomainsandgenestructuresꎬwhichwereconsistentwiththefindingsfromphylogenetictreeanalysis.TheMaAAO1genedisplayedsignificantlyhighex ̄pressionlevelsduringfruitdevelopmentandripeningperiodsꎬsuggestingitspotentialimportanceinbananafruitdevelopment.FurthermoreꎬbothMaAAO1andMaAAO2mightexhibitsensitiveresponsestoosmoticstressꎬandtheMaAAO2genemightberesponsivetotheinvasionofFoc4pathogen.BasedontheseresultsꎬitwasinferredthatMaAAOgenesplayedpivotalrolesinregulatingbananagrowthꎬdevelopmentandresponsestobioticandabioticstresses.ThesefindingswouldnotonlyenhanceourcomprehensiontotheMaAAOgenefami ̄lyinbananaꎬbutalsoprovideareferenceforfurtherunravelingtheirmechanismsofactioningrowthꎬdevel ̄opmentandstressresponsesofbanana.Keywords㊀BananaꎻAbscisicaldehydeoxidase(AAO)ꎻGeneidentificationꎻGeneexpression㊀㊀脱落酸(abscisicacidꎬABA)属于六种植物激素之一ꎬ1963年首次在脱落果实中被鉴定为生长抑制剂[1]ꎬ随后发现其在种子休眠㊁萌发㊁气孔开合㊁果实发育等多种生理过程中发挥着重要作用[2-4]ꎬ在水杨酸介导的生物胁迫以及高盐㊁干旱和寒冷等非生物胁迫中也起着关键作用[3ꎬ5]ꎮABA的代谢过程已有相关研究综述进行了详细总结[6]ꎮABA合成过程主要涉及玉米黄质环氧化酶(zeaxanthinepoxidaseꎬZEP)㊁9-顺环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoiddioxygen ̄asesꎬNCEDs)㊁短链醇脱氢酶/还原酶(short-chaindehydrogenases/reductasesꎬSDR)㊁ABA醛氧化酶(abscisicaldehydeoxidaseꎬAAO)[7]ꎬ其中AAO参与ABA合成途径中的最后一步ꎮ虽然ABA合成相关基因的研究比较多[8]ꎬ但关于AAO基因的研究较少ꎬ目前只对少数几个物种如拟南芥[9]㊁水稻[10]㊁小麦[11]等的AAO基因家族进行了鉴定分析ꎮ如:在拟南芥中ꎬAtAAO3基因的突变会导致ABA含量降低[9]ꎻ番茄ABA缺陷型突变体中的AAO活性远低于野生型的[12]ꎬ大麦中也存在类似的AAO缺失或活性降低的现象[13]ꎮ香蕉(Musassp.)是世界上重要的热带水果之一ꎬ也是重要的粮食作物之一[14]ꎮ香蕉在生长过程中会遭受到低温㊁干旱㊁香蕉枯萎病等不同逆境的影响ꎬ对其产量和最终果实品质产生重要影响[15-16]ꎮABA在这些过程中都扮演着重要的角色ꎬ因此ꎬ研究香蕉AAO家族基因种类及其在不同逆境处理下的表达情况具有重要意义ꎮ本研究从香蕉基因组中鉴定得到了AAO家族基因ꎬ并分析了它们的进化关系㊁基因结构和蛋白结构域ꎬ同时分析了其在不同组织㊁果实发育和成熟的不同阶段及对非生物/生物胁迫响应的表达模式ꎬ以期为进一步明确MaAAOs基因在香蕉生长发育和胁迫反应过程中的功能提供参考ꎮ1㊀材料与方法1.1㊀试验材料选用口味优良的香蕉品种粉蕉(MusaABBPisangAwakꎬFJ)进行试验ꎮ将粉蕉组培苗种植于塑料盆中(无菌土壤)ꎬ培养于生长室(28ħꎬ70%湿度ꎬ光周期为16h光/8h暗ꎬ光照强度为200μmol m-2 s-1)ꎮ组培苗种植取样周期为2015年6月 2016年8月ꎮ1.2㊀试验处理与样品采集方法(1)不同组织样品采集:组培苗种植约70天后达到五叶期ꎬ此时选取叶和根ꎻ组培苗种植约10个月开始开花ꎬ于开花后0天(0DAF)㊁20DAF和80DAF选取果实ꎻ组培苗种植12~13个月后采收ꎬ选取采收后3天(3DPH)和6DPH的果实ꎮ每个样本进行两次生物学重复ꎬ用于分析基因在不同组织和果实发育不同阶段的表达情况ꎮ(2)非生物胁迫处理方法及取样:分别用300mmol L-1NaCl和200mmol L-1甘露醇灌溉五叶期香蕉幼苗ꎬ进行盐胁迫和渗透胁迫处理ꎬ处理7d后取样ꎻ将五叶期香蕉幼苗置于4ħ生长室(70%湿度ꎬ光周期为16h光/8h暗ꎬ光照强度为200μmol m-2 s-1)进行冷胁迫处理ꎬ22h后取样ꎮ采集对应处理时间后的叶片样本进行非生物胁迫处理下基因的表达分析ꎮ(3)尖孢镰刀菌侵染处理及取样:将五叶期香蕉幼苗根部浸泡在F.oxysporumrace4(Foc4)孢子悬液(106个分生孢子/mL)中2hꎬ以浸入无菌蒸馏水(ddH2O)中的为对照ꎻ然后移栽到装有无菌土壤的塑料盆中ꎬ在生长室中培养(28ħꎬ70%湿度ꎬ光周期为16h光/8h暗ꎬ光照强度为200μmol m-2 s-1)ꎻ培养2d后ꎬ采集根系样品进行基因表达分析ꎮ每份样本包含两个生物重复样61㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀本ꎮ1.3㊀香蕉AAO基因家族的鉴定及系统发育分析利用拟南芥AtAAOs基因的序列构建HMM模型ꎬ从香蕉A基因组中搜索得到香蕉AAOs序列ꎻ利用得到的香蕉AAOs序列构建新的HMM模型ꎬ利用新的HMM模型从A基因组中搜索鉴定MaAAOs基因ꎮ使用保守结构域数据库(http://www.ncbi.nlm.nih.gov/cdd/)和PFAM数据库(ht ̄tp://pfam.sanger.ac.uk/)验证得到的MaAAOs基因ꎮ利用下载得到的AtAAOs和水稻OsAAOs蛋白序列ꎬ以MEGA-X中的MUSCLE方法进行序列比对ꎬ使用Neighbor-joining法构建系统发育树ꎬBootstrap值设置为1000ꎮ1.4㊀香蕉AAO基因家族的蛋白质特性和序列分析通过ExPASy数据库(http://expasy.org/)对分子质量和等电点等理化性质进行预测ꎮ利用MEME软件和InterProScan数据库对蛋白结构域序列进行鉴定ꎮ采用GSDS数据库对基因结构进行分析(http://gsds.cbi.pku.edu.cn/)ꎮ1.5㊀转录组分析RNA测序中的RNA提取㊁文库制备和测序等工作均由美吉生物技术有限公司(中国上海)完成ꎮRNA-seq分析样本的收集过程参考前人的研究[17]ꎮ每个样本包含两个生物重复序列ꎮ测序平台为IlluminaGAII(IlluminaꎬSanDiegoꎬCAꎬUSA)ꎮFASTX(http://hannonlab.cshl.edu/fastx_toolkit/)和FastQC(http://www.bioinformat ̄ics.babraham.ac.uk/projects/fastqc/)用于删除接头序列和低质量序列ꎮ通过TophatV.2.0.10将粉蕉样本的cleanreads与香蕉基因组进行比对[18]ꎬ使用Cufflinks进行转录组组装[19]ꎮ基因的表达值使用FPKM值表示ꎮ使用DEGseq工具鉴定差异表达基因(DEGs)(log2-basedfoldchange>1ꎻlog2-basedfoldchange<-1)ꎮ2㊀结果与分析2.1㊀MaAAOs基因的鉴定利用香蕉AAO基因序列的保守结构域构建HMM模型ꎬ从香蕉A基因组中鉴定得到了3个MaAAOs基因ꎬ分别命名为MaAAO1㊁MaAAO2㊁MaAAO3(表1)ꎬ其氨基酸残基数量分别为1365㊁1393㊁1399个ꎬ编码蛋白的分子量范围为15.05~15.26kuꎬ等电点范围是6.59~6.62ꎮ为分析MaAAOs的进化关系ꎬ分别下载了水稻和拟南芥AAO基因家族的蛋白质序列(表1)ꎬ采用NJ法构建了系统发育树(图1)ꎮ可见ꎬ所有AAO蛋白可以分成两类ꎬMaAAO2和MaAAO3与所有AtAAOs及大部分OsAAOs聚在一个亚类ꎬ而MaAAO1则与两个OsAAOs聚成一个亚类ꎮ㊀㊀表1㊀AAO基因信息基因名基因编号基因位置氨基酸数量/个分子量/ku等电点MaAAO1Ma06_t26720.1chr6:28580510-28613339136515.056.62MaAAO2Ma09_t10080.2chr9:6883097-6895047139315.196.37MaAAO3Ma09_t10100.1chr9:6896480-6908654139915.266.59AtAAO1AT5G20960.1chr5:7116455-7122747136814.966.34AtAAO2AT3G43600.1chr3:15511832-15517545132114.465.41AtAAO3AT2G27150.1chr2:11601727-11607199133214.676.63AtAAO4AT1G04580.1chr1:1252005-1257893133714.736.28OsAAOLOC_Os03g31550.1chr3:17985563-17998498137015.026.99LOC_Os03g31550.2chr3:17985563-17998498127313.997.27LOC_Os07g07050.2chr7:3476288-34718345486.037.47LOC_Os07g07050.1chr7:3476921-34717896056.688.44LOC_Os03g57680.1chr3:32877206-32869482135714.536.45LOC_Os03g57690.1chr3:32886266-32879566135614.516.17LOC_Os10g04860.1chr10:2358790-2368732135914.556.42LOC_Os07g18120.1chr7:10724599-10738019136614.857.24LOC_Os07g18154.1chr7:10748746-107608488459.216.65LOC_Os07g18154.2chr7:10748746-107608337277.926.78㊀㊀注:基因名中Ma代表香蕉ꎬAt代表拟南芥ꎬOs代表水稻ꎮ71㊀第10期㊀㊀㊀㊀㊀曾坚ꎬ等:香蕉中ABA醛氧化酶基因家族的鉴定及其表达分析图1㊀AAO家族蛋白系统进化树2.2㊀MaAAOs基因家族的结构域和基因结构分析利用MEME数据库从MaAAOs基因中鉴定得到10个保守结构域ꎬ并用InterPro数据库进行了注释ꎮ由结果(图2㊁表2)可见ꎬ3个MaAAOs基因表现出类似的结构域组成ꎬ仅MaAAO1缺少了Motif8ꎻ除了Motif7ꎬ其余9个Motif都含有结构域IPR016208ꎬ其功能被注释为醛氧化酶/黄嘌呤脱氢酶ꎮ随后对MaAAOs基因结构进行分析ꎬMaAAO2和MaAAO3有类似的基因结构ꎬ都有10个内含子ꎻ而MaAAO1的内含子则是14个(图3)ꎮ表明相同聚类有类似的结构域组成和基因结构ꎬ与系统发育树的分析结果相一致ꎮ图2㊀MaAAOs基因结构域㊀㊀表2㊀MaAAOs基因结构域的注释编号序列结构域(注释)Motif1CLTLLCSINFCSVITSEGLGNSKDGFHPIHERFAGFHASQCGFCTPGMCMIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif2LRRPVRMYLDRKTDMIMTGGRHPMKINYSVGFKSDGKITALHVDIFINAGIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif3EVEVDVLTGGTIILRTDLIYDCGQSLNPAVDLGQIEGAFVQGIGFFMLEEIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif4AIPDEDNCILVYSSTQCPEIAQGVIAKCLGIPDHNVRVITRRVGGGFGGKIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif5NSDGLVISDGTWTYKIPTIDBIPKQFNVKLLKSGHHEKRVLSSKASGEPPIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif6KITKFEAEKAIAGNLCRCTGYRPIVDVCKSFAABVDLIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif7GDAPEYTLPAJIDELASSADYLDRLEIIRHFNSCNKWRKRGISLVPVVY无Motif8ELHSNERLVFSKIADHMDKVASPFIRNMASLGGNLIMAQRSQFASDVATIIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif9YNWGALSFDARICKTNFPTKSAMRGPGDVQGSFIAESVIEHIPR016208(醛氧化酶/黄嘌呤脱氢酶)Motif10FTGPKLGCGEGGCGACVVLLSTYDPVSGQVKEFIPR016208(醛氧化酶/黄嘌呤脱氢酶)81㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀图3㊀MaAAOs基因的结构分析2.3㊀MaAAOs在粉蕉不同组织中的表达如图4A所示ꎬ粉蕉根中MaAAO1和MaAAO3基因高表达(FPKM>5)ꎬ叶中3个MaAAOs基因都表现出高表达ꎬ而果实(80DAF)中则只有MaAAO1表现出高表达ꎮ3个MaAAOs基因中ꎬ只有MaAAO1在粉蕉根㊁叶和果实中都呈现出高表达ꎮ图4㊀MaAAOs基因在香蕉不同组织(A)㊁果实不同发育阶段(B)㊁不同逆境处理(C)中的表达分析2.4㊀MaAAOs在粉蕉果实不同发育阶段中的表达为明确MaAAOs基因在香蕉果实发育过程中的功能ꎬ对其在不同发育阶段果实中的表达情况进行了分析ꎬ结果(图4B)显示ꎬ只有MaAAO1在果实发育早期(0DAF和20DAF)和后期(80DAF㊁3DPH)中表现出高表达(FPKM>5)ꎬ且以3DPH果实中的表达量最高ꎻMaAAO2在果实发育整个阶段的表达都极低ꎻMaAAO3在果实各发育阶段的表达水平相比MaAAO2要高ꎬ但除20DAF外均没有达到高表达ꎮ因此推测MaAAO1可能在果实发育过程中发挥着重要作用ꎮ2.5㊀MaAAOs在不同逆境处理下的表达为分析MaAAOs基因在粉蕉应对逆境胁迫中的功能ꎬ设置生物和非生物胁迫处理ꎬ分析3个MaAAOs基因在粉蕉植株根中的表达情况ꎬ结果(图4C)显示ꎬ在Foc4处理下ꎬMaAAO2基因表现出上调(log2-basedfoldchange>1)ꎻ在渗透胁迫下ꎬMaAAO1和MaAAO2基因表现出上调ꎻ在冷和盐胁迫下ꎬ3个基因均未表现出明显的上调表达ꎻMaAAO3基因在所有逆境处理中都没有表现出显著变化ꎮ3㊀讨论香蕉既是一种重要的热带和亚热带水果ꎬ也是全球130多个国家的主粮ꎬ但其研究进展相比其它作物要慢[20]ꎮABA在植物的生长发育和生物/非生物胁迫响应中起着重要作用[21]ꎬ而AAO是ABA合成途径中的重要合成酶ꎬ但香蕉AAO基因家族的情况仍不清楚ꎮ本研究从香蕉A基因组中鉴定出3个MaAAOs基因ꎬ保守结构域分析证明这3个基因属于AAO家族ꎬ并根据系统发育树将其分为两个亚类ꎬ其中MaAAO2和MaAAO3聚为一类ꎮ拟南芥的AAO基因数量是4个[9]ꎬ水稻中可能是5个[10]ꎬ小麦中是3个[11]ꎬ数量均较少ꎬ表明AAO蛋白可能属于小基因家族编码的蛋白质ꎮ91㊀第10期㊀㊀㊀㊀㊀曾坚ꎬ等:香蕉中ABA醛氧化酶基因家族的鉴定及其表达分析香蕉果实的发育和成熟过程决定着果实的品质和产量[14]ꎮ研究表明ꎬABA信号参与了果实的生长发育并影响着果实的成熟过程和最终品质ꎻ未成熟水果中的内源ABA含量通常较低ꎬ施加外源ABA能促进果实成熟及果肉软化ꎻ果实成熟过程中ABA合成相关基因的表达上调导致内源ABA大量积累[22]ꎮ这些结果表明ABA在水果成熟过程中发挥着重要作用ꎮ在本研究中ꎬ不同MaAAOs基因在粉蕉果实不同发育阶段表现出不同的表达模式ꎬMaAAO1基因在早期和晚期都表现出高表达ꎬ而MaAAO2和MaAAO3几乎不表达ꎻ此外ꎬMaAAO1在粉蕉叶㊁根㊁果实中均表现出高表达ꎬ推测MaAAO1基因在香蕉果实发育和成熟过程中可能具有重要作用ꎮ香蕉生长过程中常会遇到干旱㊁盐㊁寒冷以及枯萎病菌感染等非生物和生物逆境ꎬ对果实品质及产量造成影响[15-16]ꎮ在拟南芥中ꎬAtAAO3和AtAAO1基因表达明显受到干旱胁迫的诱导[9ꎬ23]ꎻ在拟南芥中过表达花生AhAAO2基因也能显著提高植株抵抗干旱胁迫的能力[24]ꎮ本研究发现ꎬMaAAO1和MaAAO2基因受渗透胁迫诱导上调表达ꎬ但受冷和盐胁迫的影响很小ꎻMaAAO3基因的表达在3种胁迫下均无显著变化ꎮ表明MaAAO1和MaAAO2基因可能在香蕉响应干旱胁迫中发挥作用ꎮ香蕉的种植生产也受到香蕉枯萎病的严重影响ꎮ有研究表明ABA能负向调控水杨酸(SA)介导的病原体反应ꎬ例如ꎬ提高植株中ABA的含量ꎬ会显著促进细菌的生长[25]ꎮ在本研究中ꎬ仅MaAAO2基因的表达显著受到Foc4处理的诱导ꎬ表明这个基因可能参与了响应Foc4侵染的过程ꎮ4㊀结论本研究从香蕉A基因组中鉴定得到了3个MaAAOs基因ꎬ经系统发育分析㊁蛋白结构域和基因结构分析ꎬ确定属于AAO基因家族ꎮMaAAOs基因参与了香蕉的生长发育㊁果实成熟和对生物/非生物胁迫的响应等过程ꎮ其中ꎬMaAAO1基因在果实发育早期和成熟阶段都表现出高表达ꎻMaAAO1和MaAAO2基因对渗透胁迫有响应ꎻMaAAO2基因的表达被Foc4诱导ꎬ可能响应该病菌的侵染ꎮ参㊀考㊀文㊀献:[1]㊀EaglesCFꎬWareingPF.DormancyRegulatorsinwoodyplants:experimentalinductionofdormancyinBetulapubescens[J].Natureꎬ1963ꎬ199(4896):874-875.[2]㊀ChernysJTꎬZeevaartJA.Characterizationofthe9 ̄cis ̄epoxy ̄carotenoiddioxygenasegenefamilyandtheregulationofab ̄scisicacidbiosynthesisinavocado[J].PlantPhysiologyꎬ2000ꎬ124(1):343-353.[3]㊀LeeSCꎬLuanS.ABAsignaltransductionatthecrossroadofbioticandabioticstressresponses[J].PlantCellandEnviron ̄mentꎬ2012ꎬ35(1):53-60.[4]㊀YoshidaTꎬMogamiJꎬYamaguchi ̄ShinozakiK.ABA ̄depend ̄entandABA ̄independentsignalinginresponsetoosmoticstressinplants[J].CurrentOpinioninPlantBiologyꎬ2014ꎬ21:133-139.[5]㊀ShinozakiKꎬYamaguchi ̄ShinozakiKꎬSekiM.Regulatorynet ̄workofgeneexpressioninthedroughtandcoldstressresponses[J].CurrentOpinioninPlantBiologyꎬ2003ꎬ6(5):410-417.[6]㊀ZeevaartJADꎬCreelmanRA.Metabolismandphysiologyofabscisicacid[J].AnnualReviewofPlantPhysiologyandPlantMolecularBiologyꎬ1988ꎬ39(1):439-473. [7]㊀NambaraEꎬMarion ̄PollA.Abscisicacidbiosynthesisandca ̄tabolism[J].AnnualReviewofPlantBiologyꎬ2005ꎬ56:165-185.[8]㊀ChenKꎬLiGJꎬBressanRayAꎬetal.Abscisicaciddynam ̄icsꎬsignalingꎬandfunctionsinplants[J].JournalofIntegra ̄tivePlantBiologyꎬ2020ꎬ62(1):25-54.[9]㊀SeoMꎬPeetersAJꎬKoiwaiHꎬetal.TheArabidopsisaldehydeoxidase3(AAO3)geneproductcatalyzesthefinalstepinab ̄scisicacidbiosynthesisinleaves[J].PNASꎬ2000ꎬ97(23):12908-12913.[10]BatthRꎬSinghKꎬKumariSꎬetal.Transcriptprofilingrevealsthepresenceofabioticstressanddevelopmentalstagespecificascorbateoxidasegenesinplants[J].FrontiersinPlantSci ̄enceꎬ2017ꎬ8:198.[11]ColasuonnoPꎬMarcotuliIꎬLozitoMLꎬetal.Characterizationofaldehydeoxidase(AO)genesinvolvedintheaccumulationofcarotenoidpigmentsinwheatgrain[J].FrontiersinPlantScienceꎬ2017ꎬ8:863.[12]SagiMꎬFluhrRꎬLipsSH.Aldehydeoxidaseandxanthinede ̄hydrogenaseinaflaccatomatomutantwithdeficientabscisicacidandwiltyphenotype[J].PlantPhysiologyꎬ1999ꎬ120(2):571-578.[13]Walker ̄SimmonsMꎬKudrnaDAꎬWarnerRL.Reducedaccu ̄mulationofABAduringwaterstressinamolybdenumcofactormutantofbarley[J].PlantPhysiologyꎬ1989ꎬ90(2):728-733.02㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀[14]HuWꎬZuoJꎬHouXWꎬetal.Theauxinresponsefactorgenefamilyinbanana:genome ̄wideidentificationandexpressionanalysesduringdevelopmentꎬripeningꎬandabioticstress[J].FrontiersinPlantScienceꎬ2015ꎬ6:742.[15]RashadYMꎬFekryWMEꎬSleemMMꎬetal.Effectsofmy ̄corrhizalcolonizationontranscriptionalexpressionofthere ̄sponsivefactorJERF3andstress ̄responsivegenesinbananaplantletsinresponsetocombinedbioticandabioticstresses[J].FrontiersinPlantScienceꎬ2021ꎬ12:742628. [16]XuYꎬLiuJHꎬJiaCHꎬetal.OverexpressionofabananaaquaporingeneMaPIP1ꎻ1enhancestolerancetomultipleabi ̄oticstressesintransgenicbananaandanalysisofitsinteractingtranscriptionfactors[J].FrontiersinPlantScienceꎬ2021ꎬ12:699230.[17]XuYꎬTieWWꎬYanYꎬetal.IdentificationandexpressionoftheBAHDfamilyduringdevelopmentꎬripeningꎬandstressre ̄sponseinbanana[J].MolecularBiologyReportsꎬ2021ꎬ48(2):1127-1138.[18]PollierJꎬRombautsSꎬGoossensA.AnalysisofRNA ̄SeqdatawithTopHatandCufflinksforgenome ̄wideexpressionanalysisofjasmonate ̄treatedplantsandplantcultures[J].MethodsinMolecularBiologyꎬ2013ꎬ1011:305-315.[19]TrapnellCꎬRobertsAꎬGoffLꎬetal.DifferentialgeneandtranscriptexpressionanalysisofRNA ̄SeqexperimentswithTo ̄pHatandCufflinks[J].NatureProtocolsꎬ2012ꎬ7(3):562-578.[20]HuWꎬWangLZꎬTieWWꎬetal.Genome ̄wideanalysesofthebZIPfamilyrevealtheirinvolvementinthedevelopmentꎬripeningandabioticstressresponseinbanana[J].ScientificReportsꎬ2016ꎬ6(1):30203.[21]HarakavaR.GenesencodingenzymesoftheligninbiosynthesispathwayinEucalyptus[J].GeneticsandMolecularBiologyꎬ2005ꎬ28(3):601-607.[22]杨方威ꎬ段懿菲ꎬ冯叙桥.脱落酸的生物合成及对水果成熟的调控研究进展[J].食品科学ꎬ2016ꎬ37(3):285-291. [23]YesbergenovaZꎬYangGHꎬOronEꎬetal.TheplantMo ̄hydroxylasesaldehydeoxidaseandxanthinedehydrogenasehavedistinctreactiveoxygenspeciessignaturesandareinducedbydroughtandabscisicacid[J].PlantJournalꎬ2005ꎬ42(6):862-876.[24]YangLꎬLiangJꎬZhouWꎬetal.IsolationandcharacterizationoftheAldehydeOxidase2genefromArachishypogaeaL.[J].PlantMolecularBiologyReportersꎬ2011ꎬ29(3):544-553. [25]FanJꎬHillLꎬCrooksCꎬetal.Abscisicacidhasakeyroleinmodulatingdiverseplant ̄pathogeninteractions[J].PlantPhys ̄iologyꎬ2009ꎬ150(4):1750-1761.12㊀第10期㊀㊀㊀㊀㊀曾坚ꎬ等:香蕉中ABA醛氧化酶基因家族的鉴定及其表达分析。


项目采购需求1.项目概况1.1.基本信息1.1.l.项目名称省自然资源厅“数字档案室”档案一体化平台运维运营服务(2024年)项目。
1.1.2.采购人广东省国土资源档案馆。
1.1.3.单位广东省国土资源档案馆。
1.1.4.项目总体目标基于广东省自然资源“数字档案室”档案一体化平台,结合广东省自然资源厅档案信息化工作需求,按照最新网络安全等级保护测评要求,提供平台基础设施和运维运营服务,提升馆信息化水平,夯实自然资源档案服务能力。
1.15服务地点广州市。
1.2.项目背景广东省国土资源档案馆高度重视档案信息化建设工作,2018年具体承担建成了广东省自然资源厅“全国示范数字档案室”,利用空间信息技术、云计算技术、大数据等技术,构建了功能完善、成熟先进、数字档案资源较为齐全完整的档案一体化平台,促进了机关档案工作转型升级。
为全面提高自然资源档案信息化管理水平和服务能力,根据自然资源档案信息化发展需要,并结合业务科室日常使用中发现的问题和提出的建议,需要在广东省自然资源厅“数字档案室”档案一体化平台建设成果的基础上,持续开展平台基础设施和运维运营工作,确保平台的安全、可靠、可用、可控,同时助推实现自然资源档案数据资产管理。
2.项目预算本项目总预算为97.59万元。
3.服务期限本项目委托服务期限为一年。
4.服务内容4.1.专业基础设施服务41.1.安全设备续租自2022年起租赁的安全设备,对馆内网络接入及行为做审计,保障网络安全,全面保障馆信息化建设有关网络安全防护的能力。
41.2..商用密码产品续租自2022年起租赁的商用密码产品,为网络和通信、应用和数据等各个层面的密码应用解决机密性、完整性、真实性的需求,以达到密码应用的合规性、正确性和有效性。
4.2.运行维护服务4.2.2.基础设施运行维护服务为馆机房的2台服务器、1套存储设备提供例行巡检、日常维护、响应支持、故障处理等运行维护服务。
4.2.3.软件系统运行维护服务提供广东省自然资源厅“数字档案室”档案一体化平台及其档案数据搜索引擎运行维护服务。
年度广东高校重点平台和科研项目申报网上申报指引、网址:。
、申报账号:已开通账号的学校科研管理部门以原账号密码登陆,学校社科管理员账号格式为“学校代码”;科技管理员账号格式为“学校代码”;教育科研管理账号格式为“学校代码”如果管理账号忘记密码可通过在申报平台点击“忘记密码”链接,按照提示发送传真到。
未开通账号的学校科研管理部门请直接与省教育厅科研处联系。
校内的项目申请人从学校科研管理人员处获取用户名和密码。
、下载申请书:项目申请人请登录省教育厅数字科研服务平台(),下载对应申报类别的最新版申请书(带宏版本);此前从其它渠道获得的申报书非正式版申报书,不保证能正常使用。
本次申报通知中的第类项目:特色创新类项目(教育科研项目),在申报平台中所对应的栏目是:教育科学研究项目(高校认定),在申报书的下载和上传过程中请注意对应相应栏目。
、填写申请书:申请书封面的所属领域编号上,申报人文社科类填,申报自然科学类填,申报教育科研项目根据研究所属学科领域选择相应编号;经费额度由各高校在本校高等教育“创新强校工程”经费中统筹安排,学校应根据项目实施的实际需求,科学合理地提出对本校推荐申报的每个平台和项目的具体资助经费,项目一经认定,原则上不得更改资助经费额度;申请书中的“计划开始日期”与“计划结束日期”,请按照申报通知中相应类别的具体要求并根据研究的实际需求自行确定,省教育厅不做统一界定;创新团队项目需以省部级重点平台为依托,在创新团队项目申请书“基本信息”中“原平台名称”一栏请按“依托平台名称”来填写,请各相关高校提醒申报者注意,在审查时也请注意此点。
学校申请人下载申请书后,离线填写,填写完成后必须设定密码保护文档,防止文档被修改。
、上传申报文档:项目申请人登录省教育厅项目申报网站,上传已保护的项目申报文档。
系统开放时间,对于第、、类项目为2018年2月19日-2019年1月11日;第、、类项目为年月日-年月日。