B.过渡阶段
C.饱和阶段
表观量子产额比理论值低,主要原因是:
①光没有全部被叶片吸收,存在反射和透射损失; ②非光合色素吸收了部分光能; ③光呼吸和暗呼吸对光合的负效应; ④形成的同化力(ATP、NADPH)没有全部用于CO2的还原; ⑤没有在饱和CO2浓度和最适温度下测定, 存在CO2扩散和 固定速率的限制等。
(4)消除乙醇酸
乙醇酸对细胞有毒害作用,它的产生在代谢 中是不可避免的。• 呼吸消除乙醇酸的代谢, 光 使细胞免受伤害。另外,光呼吸代谢中涉及 多种氨基酸的转化过程,它可能对绿色细胞 的氮代谢有利。
五、C3植物和C4植物的光合特征
C4植物比C3植物具有较强的光合作用,这与其结构特征 和生理特性有关。 (一)解剖结构特点
1.光强(light)
(1)光强-光合曲线 光补偿点(Light Compensation Point,LCP): 随着光强的增高,光合速率相应提高,当达到某一光强时,叶片 的光合速率与呼吸速率相等 ,净光合速率为零时的光强称为光补偿 点。 光饱和点(Light Saturation Point,LSP):
利用植物的需光特性确定合理的叶面积系数 (leaf area index, LAI)
单位土地面积上的叶面积
LAI= 单位土地面积
LAI是表示作物群体大小状况的一个指标。合理的LAI应 以作物的需光特性为标准,以最大限度的利用光能、地力为 原则,以高产、优质、低消耗为目的。从作物的需光特性上 讲,应使上部叶片处于光饱和点,中下部叶片处于光补偿点 的两倍以上,以保证下部叶片及根系的正常生长。
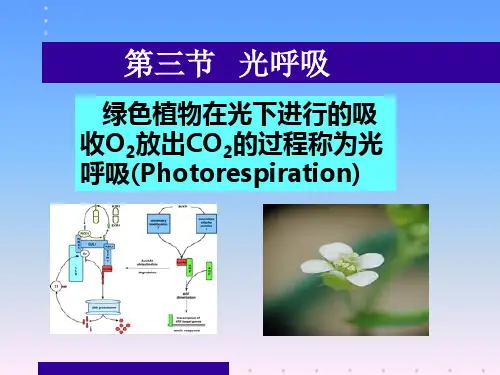
在叶绿体与过氧化体中吸收氧气,在线粒体中放出CO2
二、光呼吸的生化历程
叶绿体 3-PGA + 磷酸乙醇酸 乙醇酸 C3—cycle C2—cycle [O2] Rubisco RUBP 2 3—PGA [CO2] 过氧化体