细胞膜与细胞表面
- 格式:ppt
- 大小:14.92 MB
- 文档页数:136
![[细胞生物学]细胞膜与细胞表面](https://uimg.taocdn.com/898e34056c175f0e7cd13712.webp)



细胞质膜简述细胞膜的生理作用1.限定细胞的范围,维持细胞的形状2.具有高度选择性,(为半透膜)并能进行主动运输使细胞内外形成不同离子浓度并保持细胞内物质和外界环境之间的必要差别3.是接受外界信号的传感器,使细胞对外界环境的变化产生适当的反应4.与细胞新陈代谢、生长繁殖分化及癌变等重要生命活动密切相关生物膜的化学组成及其特点和意义构成生物膜的主要成分是脂类和蛋白质。
其中脂类包括磷脂、糖脂和硫脂等,几乎都是两性分子,在水相中磷脂分子亲水的头部朝向水相,疏水的尾部相对,自发排列成疏水双分子层,而且双分子膜一旦破损也能自我闭合。
磷脂双分子层的这种自我装配、自我闭合的特点赋予细胞细胞膜对细胞起保护作用,使每一个细胞成为一个相对独立的整体。
脂双层分子具有流动性,有利于嵌在膜内的功能蛋白的旋转和转移,便于其发挥相应的作用细胞膜中的蛋白质多种多样:从组成看有单纯蛋白质、糖蛋白和脂蛋白等。
从结合状态看有不同的镶嵌方式;从功能来分,有载体蛋白、受体蛋白和各种酶等。
由此保证有控制细胞内外的物质交换的作用和细胞间相互识别以及传递各种信息的作用、感受和传递各种刺激的作用等多种功能,还使细胞具有多样性,保证了不同组织细胞和不同发育时期细胞膜功能的差异性。
生物膜的基本结构特征是什么?与它的生理功能有什么联系?(指导)生物膜的基本结构特征:1.磷脂双分子层组成生物膜的基本骨架,具有极性的头部和非极性的尾部的脂分子在水相中具有自发形成封闭膜系统的性质,以非极性尾部相对,极性头部朝向水中。
这一结构特点为细胞核细胞器的生理活动提供了一个相对稳定的环境,使细胞与外界、细胞器与细胞器之间有了一个界面2.蛋白质分子以不同的方式镶嵌其中或者结合与表面,蛋白质的类型、数量多少、蛋白质分布的不对称性以及脂分子的协同作用赋予生物膜不同的特性和功能,这些结构有利于物质的选择运输,提供细胞识别位点,为多种酶提供了结合位点,同时参与形成不同功能的细胞表面结构特征。


《细胞生物学》题库第四章细胞膜与细胞表面一、名词解释1. 脂质体——脂质体是根据磷脂分子可在水相中形成稳定的脂双层膜的趋势而制备的人工膜,脂质体中可以裹入不同的药物或酶等具有特殊功能的生物大分子。
2. 流体镶嵌模型——主要强调:1.膜的流动性,膜脂和膜蛋白均可侧向运动2.膜蛋白分布的不对称性3. 细胞膜——又称质膜,是指围绕在细胞最外层,由脂质和蛋白质组成的生物膜。
4. 去垢剂——是一端亲水一端疏水的两性小分子,是分离与研究膜蛋白的常用试剂。
5. 膜内在蛋白——又称整合蛋白,多数为跨膜蛋白,与膜紧密结合。
6. 细胞外被——又称糖萼,曾用来指细胞膜外表面覆盖的一层粘多糖基质,实际上细胞外被中的糖与细胞膜的蛋白分子或脂质分子是共价结合的,形成糖蛋白和糖脂,所以,细胞外被应是细胞膜的正常结构组分,它不仅对膜蛋白起保护作用,而且在细胞识别中起重要作用。
7. 细胞外基质——是指分布于细胞外空间,由细胞分泌的蛋白和多糖所构成的网络结构。
细胞外基质将细胞粘连在一起构成组织,同时,提供一个细胞外网架,在组织中或组织之间起支持作用。
8. 透明质酸——是一种重要的糖胺聚糖,是增殖细胞和迁移细胞胞外基质的主要成分,尤其在胚胎组织中。
9. 细胞连接——是多细胞有机体中相邻细胞之间通过细胞质膜相互联系,协同作用的重要组织方式。
10. 细胞粘着——在细胞识别的基础上,同类细胞发生聚集,形成细胞团或组织的过程。
11. 整联蛋白家族——细胞膜上能够识别并结合各种能够含RGD三肽顺序的受体称整联蛋白家族。
12. 连接子——构成间隙连接的基本单位。
13. 免疫球蛋白超家族的CAM——分子结构中具有与免疫球蛋白类似的结构域的CAM超家族。
6.C7.A8.C9.C 10. B 11.C 12.C 13.B 14.D 15.A 16.B 17.B 18.D 19.C 20.D 21.B 22.C1. 膜脂的主要成分包括①磷脂②糖脂③胆固醇④中性脂质2. 膜脂分子有4种运动方式,其中生物学意义最重要的是.侧向运动3. 与细胞质基质接触的膜面称为质膜的.PS4. 细胞外被又称D.糖萼5. 胶原是胞外基质最基本成分之一。



第四章细胞膜与细胞表面第一节细胞膜与细胞表面特化结构细胞膜(cell membrane)又称质膜(plasma membrane):是指围绕在细胞最外层,由脂质和蛋白质组成的生物膜。
细胞膜不仅是细胞结构上的边界,使细胞有一个相对稳定的内环境,同时在细胞与环境之间进行物质、能量的交换及信息传递过程中也起着决定性的作用。
生物膜(biomembrane):真核细胞内部存在着由膜围绕构建的各种细胞器。
细胞内的膜系统与细胞膜统称为生物膜。
它们具有共同的结构特征。
一、细胞膜的结构模型人们用光学显微镜发现了细胞,但到20世纪50年代初,在电镜下显示出了质膜的超微结构。
但人们并未感到惊奇,因为此前细胞生理学家在研究细胞内渗透压时已证明了质膜的存在。
1925年E. Gorter和F. Grendel研究红细胞发现膜脂单层分子为红细胞表面积的二倍,提示了质膜是由双层脂分子构成的。
随后,人们发现质膜的表面张力比油—水界面的表面张力低得多,若脂滴表面吸附有蛋白成分则表面张力降低,因此Davson和Danielli提出“蛋白质—脂质—蛋白质”的三明治式的质膜结构模型。
这一模型影响达20年之久。
1959年,J. D. Robertson发展了三明治模型,提出了单位膜模型(unit membrane model),并推断所有的生物膜都由蛋白质—脂质—蛋白质的单位膜构成。
随后的一些实验,如免疫荧光标记技术等证明,质膜中的蛋白质是可流动的;冷冻蚀刻技术显示了双层膜脂中存在膜蛋白颗粒。
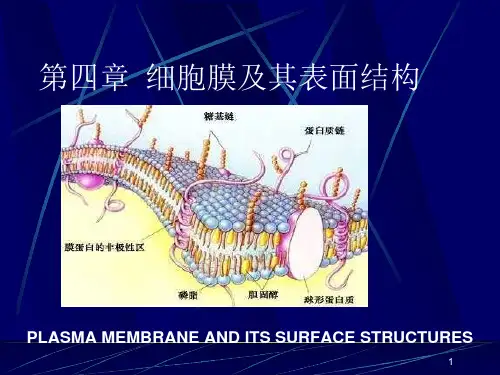
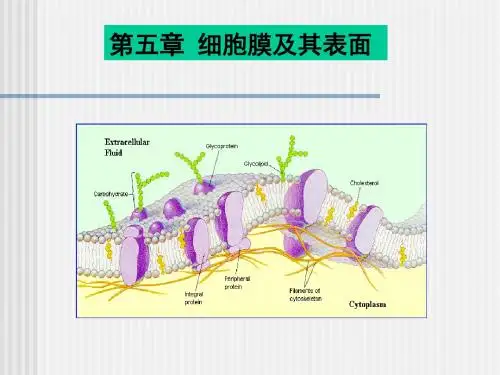
1972年,S. J. Singer和G. Nicolson在此基础上又提出了生物膜的流动镶嵌模型(fluid mosaic model) 。
这一模型随即得到各种实验结果的支持。
流动镶嵌模型主要强调:①膜的流动性,膜蛋白和膜脂均可侧向运动;②膜蛋白分布的不对称性,有的镶在膜表面,有的嵌入或横跨脂双分子层。
近年来有人提出脂筏模型(lipid rafts model),即在生物膜上富含胆固醇, 形成有序的脂相,如同“脂筏”一样, 并载有各种蛋白。