第六章 细胞的能量转换——线粒体和叶绿体
- 格式:docx
- 大小:26.37 KB
- 文档页数:3



第六章细胞的能量转换——线粒体和叶绿体一、填空题1.能对线粒体进行专一染色的活性染料是____。
2.线粒体膜围腔有2个,它们是____和____,而叶绿体膜围腔有3个,自外向里分别是____,____和____。
3.线粒体各部分结构中有各自特殊的标记酶,它们分别在外膜是____,外腔是____,内膜是____,内腔是____。
4.三羧酸循环是在线粒体____中进行,氧化磷酸化过程是在____上,由____和____完成。
5.根据接受代谢物上脱下的氢的原初受体的不同,可以将细胞中的呼吸链分为两种典型的类型,分别为____和____。
6.ATP合成酶合成ATP的直接能量来自____。
7.在线粒体电子传递链的4种复合物中既是电子传递体又是质子位移体的是____、____和____。
8.在线粒体电子传递链中电子传递方向按氧化还原电势____的方向传递。
9.叶绿体在显微结构上主要分为____、____、____。
10.叶绿体的超微结构可分为____、____、____、____、____几部分。
11.叶绿体基质中的类囊体可以根据形态不同分为____和____。
12.氧是在植物细胞中____部位上所进行的____的过程中产生的。
13.叶绿体类囊体膜上色素分子按照其作用可以分为两大类,分别为____和____。
14.捕光色素和反应中心构成了____,它是进行光合作用的最小结构单位。
15.光合作用按照是否需要光可分为____和____两步,其中光反应又可分为____和____两步。
光反应发生在____,暗反应发生在____。
16.电子沿光合电子传递链传递时,根据最终电子受体的不同,光合磷酸化可分为____和____两条通路。
17.植物细胞中具有特异的质体细胞器,主要分为____、____、____。
18.线粒体的增殖是由____而来。
19.叶绿体的发育是由____分化而来。
20.线粒体和叶绿体的生长和增殖是受____两套遗传系统控制,所以称为半自主性细胞器。

第六章细胞的能量转换--线粒体和叶绿体线粒体和叶绿体是细胞内两个能量转换细胞器,它们能高效地将能量转换成ATP。
线粒体广泛存在于各尖真核细胞,而叶绿体仅存在于植物细胞中。
它们的形态结构都呈封闭的双层结构,内膜都演化为极其扩增的特化结构,并在能量转换中起主要作用。
线粒体和叶绿体以类似的方式合成ATP。
线粒体和叶绿体都是半自主性细胞器。
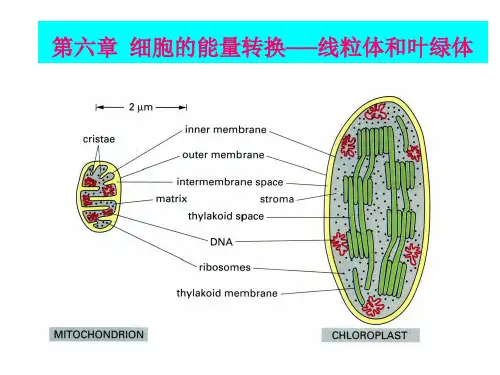
第一节线粒体与氧化磷酸化一、线粒体的形态结构(一)线粒体的形态与分布线粒体多呈线状和粒状,是由两层单位膜套叠而成的封闭囊状结构,由外膜、内膜、膜间隙及基质4部分构成。
(二)线粒体的结构与化学组成线粒体的化学成分主要是蛋白质和脂类。
线粒体的蛋白可分为可溶性与不溶性两类,可溶性蛋白质大多数是基质中的酶和膜外周蛋白;不溶性蛋白是膜的镶嵌蛋白、结构蛋白和部分酶蛋白。
线粒体脂类主要成分是磷脂。
蛋白质(线粒体干重的65~70%)脂类(线粒体干重的25~30%):磷脂占3/4以上,外膜主要是卵磷脂,内膜主要是心磷脂,线粒体脂类和蛋白质的比值:0.3:1(内膜);1:1(外膜)线粒体内外膜在化学组成上根本不同是脂类和蛋白质的比值不同。
内膜的脂类与蛋白质的比值低,外膜中比值较高。
1 外膜外膜:厚约6nm,含孔蛋白(porin),通透性高。
2 内膜厚约6-8nm,通透性低,只有不带电荷的小分子才能通过。
内膜向内褶叠形成嵴。
内膜和嵴的基质面上有许多排列规则的基粒,基粒由头部和基部组成,头部又叫F1,基部又叫F0。
3 膜间隙内外膜之间宽8nm的空隙,它延伸到嵴的轴心部(嵴内隙),内含许多可溶性酶类、底物和辅助因子。
4 线粒体基质内膜和嵴包围的空间,内含蛋白质性质的胶状物质。
基质中有催化三羧酸循环、脂肪酸 -氧化、氨基酸氧化、蛋白质合成等有关的酶类和其它成分,如环状DNA、RNA、核糖体及较大的致密颗粒,其作用主要是贮存Ca+。
线粒体酶的定位线粒体约有140种酶,分布在各个结构组分中,有的可作为某一部位所特有的标志酶,如外膜的单胺氧化酶,膜间隙的腺苷酸激酶,内膜的细胞色素氧化酶,基质中的苹果酸脱氢酶。



![细胞生物学[第六章细胞的能量转换—一线粒体和叶绿体]课程预习](https://uimg.taocdn.com/7b24cc1c14791711cc791745.webp)
第六章细胞的能量转换—一线粒体和叶绿体线粒体和叶绿体是细胞内的两种产能细胞器。
它们最初的能量来源有所不同,但却有着相似的基本结构,而且以类似的方式合成ATP。
线粒体是一种高效地将有机物转换为细胞生命活动的直接能源ATP的细胞器。
叶绿体通过光合作用把光能转换为化学能,并储存于糖类、脂肪和蛋白质等大分子有机物中。
线粒体和叶绿体都具有环状DNA及自身转录RNA与翻译蛋白质的体系。
线粒体和叶绿体都是半自主性的细胞器。
一、线粒体和氧化磷酸化线粒体通过氧化磷酸化作用,进行能量转换,为所需要的细胞进行各种生命活动提供能量。
(一)线粒体的形态结构1.线粒体的形态与分布线粒体一般呈粒状或杆状,但因生物种类和生理状态而异,可呈环形、哑铃形、线状、分权状或其他形状。
主要化学成分是蛋白质和脂类,其中蛋白质占线粒体干重的65%~70%,脂类占25%~30%。
一般直径0.5~1μm,长1.5~3.0μm,在胰脏外分泌细胞中可长达10~20μm,称巨线粒体。
数目一般数百到数千个,植物因有叶绿体的缘故,线粒体数目相对较少;肝细胞约1300个线粒体,占细胞体积的20%;单细胞鞭毛藻仅1个,酵母细胞具有一个大型分支的线粒体,巨大变形中达50万个;许多哺乳动物成熟的红细胞中无线粒体。
通常结合在维管上,分布在细胞功能旺盛的区域。
线粒体在细胞质中可以向功能旺盛的区域迁移,微管是其导轨,由马达蛋白提供动力。
2.线粒体的结构与化学组成线粒体的超微结构在电镜下观察到线粒体是由两层单位膜套叠而成的封闭的囊状结构。
主要由外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)、基质(matrix)或内室(inner chamber)四部分组成。
(1)外膜是包围在线粒体最外面的一层单位膜,光滑而有弹性,厚约6μm。
外膜上有排列整齐的筒状圆柱体,其成分为孔蛋白(porin),圆柱体上有小孔。
(2)内膜位于外膜内侧,把膜间隙与基质(内室)分开。
第六章细胞的能量转换—一线粒体和叶绿体线粒体和叶绿体是细胞内的两种产能细胞器。
它们最初的能量来源有所不同,但却有着相似的基本结构,而且以类似的方式合成ATP。
线粒体是一种高效地将有机物转换为细胞生命活动的直接能源ATP的细胞器。
叶绿体通过光合作用把光能转换为化学能,并储存于糖类、脂肪和蛋白质等大分子有机物中。
线粒体和叶绿体都具有环状DNA及自身转录RNA与翻译蛋白质的体系。
线粒体和叶绿体都是半自主性的细胞器。
一、线粒体和氧化磷酸化线粒体通过氧化磷酸化作用,进行能量转换,为所需要的细胞进行各种生命活动提供能量。
(一)线粒体的形态结构1.线粒体的形态与分布线粒体一般呈粒状或杆状,但因生物种类和生理状态而异,可呈环形、哑铃形、线状、分权状或其他形状。
主要化学成分是蛋白质和脂类,其中蛋白质占线粒体干重的65%~70%,脂类占25%~30%。
一般直径0.5~1μm,长1.5~3.0μm,在胰脏外分泌细胞中可长达10~20μm,称巨线粒体。
数目一般数百到数千个,植物因有叶绿体的缘故,线粒体数目相对较少;肝细胞约1300个线粒体,占细胞体积的20%;单细胞鞭毛藻仅1个,酵母细胞具有一个大型分支的线粒体,巨大变形中达50万个;许多哺乳动物成熟的红细胞中无线粒体。
通常结合在维管上,分布在细胞功能旺盛的区域。
线粒体在细胞质中可以向功能旺盛的区域迁移,微管是其导轨,由马达蛋白提供动力。
2.线粒体的结构与化学组成线粒体的超微结构在电镜下观察到线粒体是由两层单位膜套叠而成的封闭的囊状结构。
主要由外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)、基质(matrix)或内室(inner chamber)四部分组成。
(1)外膜是包围在线粒体最外面的一层单位膜,光滑而有弹性,厚约6μm。
外膜上有排列整齐的筒状圆柱体,其成分为孔蛋白(porin),圆柱体上有小孔。
(2)内膜位于外膜内侧,把膜间隙与基质(内室)分开。
第六章细胞的能量转换--线粒体和叶绿体(本章内容详见生物化学)一、线粒体的形态结构(一)线粒体的形态与分布线粒体是一个动态细胞器,在生活细胞中具有多样性、易变性、运动型和适应性等特点,其形状、大小、数量与分布在不同细胞内变动很大,机制在同一细胞,随着代谢条件的不同也会发生变化。
线粒体的形状多种多样,但以线状和粒状最常见。
(二)线粒体的结构与化学组成线粒体只有外层彼此平行的单位膜套叠而成的封闭的囊状结构。
外膜(outer membrane)起界膜作用,内膜(inner membrane)向内折叠形成嵴(cristae)。
外膜和内膜将线粒体分割成两个区室:一个是内外膜之间的腔隙,称为膜间隙(intermembrane space);另一个是内膜所包围的空间,称为基质(matrix)。
1、外膜线粒体最外面的一层单位魔结构,厚约6nm,光滑而有弹性。
外膜中蛋白质和脂质约各占50%。
外膜含有孔蛋白(porin),孔蛋白是由β链形成桶状结构,中心是一直径为2~3nm的小孔,即内部通道。
外膜的标志酶是单胺氧化酶(monoamine oxidase)。
2、内膜内膜是位于外膜的内侧把膜间隙与基质分开的一层单位膜结构,厚6~8nm。
相对外膜而言,内膜有很高的蛋白质/脂质比(质量比>3:1)。
内膜缺乏胆固醇,富含心磷脂(cardiolipin),约占磷脂含量的20%,心磷脂与离子的不可渗透性有关。
线粒体内膜除含有多种转运系统外,还含有大量的合成ATP的装置。
在内膜上有许多排列规则的颗粒,称为线粒体基粒(elementary particle),又称耦联因子1(coupling factor 1),简称F1,实质是ATP合酶(ATP synthesis)的头部。
3、膜间隙线粒体内、外膜之间的腔隙,宽6~8nm,但在细胞进行活跃呼吸时,膜间隙可扩大。
膜间隙中充满无定型液体,含有可溶性的酶、底物和辅助因子。
其中腺苷酸激酶是膜间隙的标志酶。
4、线粒体基质内膜所包围的嵴外空间为线粒体基质。
(P130)二、线粒体的功能线粒体是物质最终彻底氧化分解的场所,其主要功能是进行三羧酸循环及氧化磷酸化合成ATP,为细胞生命活动提供直接能量。
此外,线粒体海域细胞中氧自由基的生成,调节细胞艳华还原电位和信号转导,调控细胞凋亡、基因表达、细胞膜内多种离子的跨膜转运及电解质稳定平衡,包括线粒体对细胞中Ca2+的稳态调节等有关。
(一)线粒体中的氧化代谢在细胞中,线粒体是氧化还原的中心,是糖类、脂质和蛋白质最终氧化释能的场所。
线粒体中的三羧酸循环(tricarboxylic acid cycle),简称TCA循环,是物质氧化的最终共同途径,氧化磷酸化是生物获得能量的主要途径。
细胞之中糖酵解产生的NADH虽不能透过内膜浸入线粒体氧化,但是NDAH上的电子去能通过两种“穿梭”途径进入线粒体:①苹果酸-天冬氨酸穿梭途径(malate-asparate shuttle);②甘油-3-磷酸穿梭途径(glycerol-3-phosphate shuttle)。
(P132)当NADH中的1对电子传递到O2时,有10个H+被泵出,二FADH2中的1对电子的传递有6个H+被泵出。
(二)电子传递链与电子传递在线粒体内膜上存在的有关氧化磷酸化的脂蛋白复合物,是由一系列能可逆地接受和释放电子或H+的化学物质组成,它们在内膜上相互关联的有序排列构成呼吸链,称为电子传递链(electron transport chain)或呼吸链(respiratory chain),是典型的多酶体系。
电子通过呼吸链的流动,称为电子传递。
1、电子载体(electron carrier)参与电子传递链的电子载体有5种:黄素蛋白、细胞色素、泛醌、铁硫蛋白和铜离子。
黄素蛋白(flavoprotein),是由一条多肽与黄素蛋白腺嘌呤单核苷酸(FMN)或黄素腺嘌呤二核苷酸(FAD)紧密结合组成的结合蛋白。
细胞色素(cytochrome),是一种带有含铁血红素辅基二队可见光具有特征性强吸收的蛋白。
泛醌(ubiquinone,UQ)或称辅酶Q(coenzyme Q,CoQ),或简称为Q,是一种脂溶性的、带有一条的类异戊二烯侧链的苯锟。
铁硫蛋白(iron-sulfur protein),是一类含非血红素铁的蛋白质。
铜离子(copper atom),位于线粒体内膜的单个蛋白质分子内。
通过Cu2+和Cu+两种状态的变换,传递单个电子。
2、电子载体排列顺序(P135)呼吸链的最终受体是氧,氧接受电子后与H+结合生成水。
3、电子转运复合物复合物Ⅰ:NADH-CoQ还原酶,又称NADH脱氢酶。
既是电子传递体又是质子移位体。
组成:由25条以上多肽链组成,以二聚体形式存在,至少6个Fe-S中心和1个黄素蛋白,是呼吸链中最大最复杂的复合物;作用:催化NADH氧化,从中获得2个高能电子→辅酶Q;泵出4 H+(质子跨膜输送)复合物Ⅱ:琥珀酸-CoQ还原酶,又称琥珀酸脱氢酶。
是三羧酸循环中唯一一种结合在膜上的酶。
组成:含FAD辅基,2Fe-S蛋白和一个细胞色素b;作用:催化电子从琥珀酸→FAD→Fe-S→辅酶Q (无H+泵出)复合物Ⅲ:CoQ-细胞色素还原酶,以二聚体形式存在,既是电子传递体又是质子移位体。
组成:2个cytb(b562,b566),1cyt c11Fe-S蛋白;作用:催化电子从CoQ→cyt c;泵出4 H+;复合物Ⅳ:细胞色素C氧化酶,有6-13条多肽链组成,以二聚体形式存在。
组成:cyt a, cyt a3 ,2个Cu原子(CuA,CuB);作用:催化电子从cyt c→分子O2形成水。
(三)质子转移与质子驱动的形成(P137)(四)ATP形成机制——氧化磷酸化将ADP转变为ATP的过程成为磷酸化(phosphorylation)。
ADP磷酸化有2种途径:①底物水平的磷酸化;②氧化磷酸化(oxidative phosphorylation)。
1、ATP合酶的结构与组成ATP合酶是生物体能量转换的核心酶。
ATP合酶包括两个基本组分:球状的F1头部和嵌于内膜的F0基部。
2、能量耦联与ATP合酶的作用机制1979年,Boyer提出结合变构机制(binding change mechanism),①质子梯度的作用并不是用于形成ATP,而是使ATP从酶分子上解脱下来。
②ATP合酶上的3个β亚基的氨基酸序列是相同的,但它们的构象不同。
第二节叶绿体与光合作用植物细胞的质体具有不同的形态和功能,通常分为叶绿体(chloroplast)、有色体(chromoplast)和白色体(leucoplast)3大类。
第三节线粒体和叶绿体是半自主性细胞器一、半自主性细胞器的概念:自身含有遗传表达系统(自主性);但编码的遗传信息十分有限,其RNA转录、蛋白质翻译、自身构建和功能发挥等必须依赖核基因组编码的遗传信息(自主性有限)。
二、线粒体和叶绿体的DNA1、mtDNA /ctDNA形状、数量、大小(1)双链环状(除绿藻mtDNA,草履虫mtDNA)(2)mtDNA大小在动物中变化不大,但在植物中变化较大高等植物,120kbp~200kbp;2、mtDNA和ctDNA的复制(1)以半保留方式进行自我复制(2) mtDNA复制的时间主要在细胞周期的S期及G2期,DNA先复制,随后线粒体分裂。
ctDNA复制的时间在G1期。
(3) 复制仍受核控制.三、线粒体和叶绿体的蛋白质合成1、线粒体和叶绿体合成蛋白质的种类十分有限2、线粒体或叶绿体蛋白质合成体系对核基因组具有依赖性3、不同来源的线粒体基因,其表达产物既有共性,也存在差异4、参加叶绿体组成的蛋白质来源有3种情况:(1)由ctDNA编码,在叶绿体核糖体上合成;(2)由核DNA编码,在细胞质核糖体上合成;(3)由核DNA编码,在叶绿体核糖体上合成。
四、线粒体和叶绿体蛋白质的运送与组装1、线粒体蛋白质的运送与组装(1)定位于线粒体基质的蛋白质的运送(2)定位于线粒体内膜或膜间隙的蛋白质运送2、叶绿体蛋白质的运送及组装(1)定位于类囊体中的蛋白运送:前体蛋白N端转运肽分为两个区域,分别引导两步转运,其N端含有导向基质的序列,引导进入基质,而C端有导向类囊体的序列,又引导其穿过类囊体膜进入类囊体腔,转运肽经历两次水解,一次在基质中,一次在类囊体腔;(2)定位于基质中的蛋白运送:转运肽仅有导向基质的序列,在基质中被切除,转运肽成为成熟蛋白。
第四节线粒体和叶绿体的增殖与起源一、线粒体和叶绿体的增殖1、线粒体的增殖:(1)间壁分离,分裂时先由内膜向中心皱褶,将线粒体分类两个,常见于鼠肝和植物产生组织中。
(2)收缩后分离,分裂时通过线粒体中部缢缩并向两端不断拉长然后分裂为两个,见于蕨类和酵母线粒体中。
(3)出芽,见于酵母和藓类植物,线粒体出现小芽,脱落后长大,发育为线粒体。
2、叶绿体的发育和增殖:(1)个体发育:由前质体(proplastid)分化而来。
(2)增殖:分裂增殖二、线粒体和叶绿体的起源1、内共生起源学说(1)1905年,Mereschkowsky提出叶绿体起源于细胞内共生的蓝藻:(2)1970年,Margulis提出:线粒体的祖先-原线粒体是一种革兰氏阴性细菌;叶绿体的祖先是原核生物的蓝细菌,即蓝藻。
(3)内共生起源学说的主要论据:①基因组在大小、形态和结构方面与细菌相似。
②有自己完整的蛋白质合成系统,能独立合成蛋白质,蛋白质合成机制有很多类似细菌而不同于真核生物。
③两层被膜有不同的进化来源,外膜与细胞的内膜系统相似,内膜与细菌质膜相似。
④以分裂的方式进行繁殖,与细菌的繁殖方式相同。
⑤能在异源细胞内长期生存,说明线粒体和叶绿体具有的自主性与共生性的特征。
⑥线粒体的祖先很可能来自反硝化副球菌或紫色非硫光合细菌。
⑦发现介于胞内共生蓝藻与叶绿体之间的结构--蓝小体,其特征在很多方面可作为原始蓝藻向叶绿体演化的佐证。
(4)不足之处:①从进化角度,如何解释在代谢上明显占优势的共生体反而将大量的遗传信息转移到宿主细胞中?②不能解释细胞核是如何进化来的,即原核细胞如何演化为真核细胞?③线粒体和叶绿体的基因组中存在内含子,而真细菌原核生物基因组中不存在内含子,如果同意内共生起源学说的观点,那么线粒体和叶绿体基因组中的内含子从何发生?2、非共生起源学说:(1)主要内容:真核细胞的前身是一个进化上比较高等的好氧细菌。
(2)成功之处:解释了真核细胞核被膜的形成与演化的渐进过程。
(3)不足之处①实验证据不多②无法解释为何线粒体、叶绿体与细菌在DNA分子结构和蛋白质合成性能上有那么多相似之处③对线粒体和叶绿体的DNA酶、RNA酶和核糖体的来源也很难解释。
④真核细胞的细胞核能否起源于细菌的核区?。