乌冈栎天然居群叶表型变异
- 格式:pdf
- 大小:545.00 KB
- 文档页数:8

香榧天然群体表型性状遗传多样性分析闵会;程诗明;康志雄;陈友吾【摘要】以香榧主要分布区6个天然群体的92个样株为试验材料,对其针叶、果实、种子的8个表型性状进行分析,讨论群体间和群体内的表型多样性.结果表明:香榧各表型性状在群体问和群体内都存在显著差异,群体间表型分化系数占10.8%,群体内占89.2%,群体间的变异小于群体内.各群体表型性状中平均变异系数最小的为果实宽(CVv =13.95%),最大的为单粒重(CVv=34.71%);不同群体各形态性状平均变异系数由大到小顺序为:黄山>磐安>嵊州>钟家岭>诸暨东白湖、绍兴.通过表型性状的聚类分析可以将香榧6个群体划分为3个类群.【期刊名称】《浙江林业科技》【年(卷),期】2009(029)002【总页数】5页(P24-28)【关键词】香榧;天然群体;遗传;表型多样性;表型分化系数【作者】闵会;程诗明;康志雄;陈友吾【作者单位】浙江林学院,浙江,临安,311300;浙江省林业科学研究院,浙江,杭州,310023;浙江省林业厅,浙江,杭州,310020;浙江省林业科学研究院,浙江,杭州,310023【正文语种】中文【中图分类】S718.54遗传多样性是生物多样性的基本组成部分,其测定对于了解物种起源、预测种源适应性及估算基因资源分布具有重要意义。
遗传标记是研究遗传多样性的主要方法,包括形态学(表型)、细胞学、生化和分子标记4大类[1]。
表型性状的变异通常是遗传变异的重要线索,既具有变异性又具有稳定性,虽然表型性状的变异还不能把表型性状和基因及其位点相对应起来,而且一些性状特别是数量性状也很容易受到环境的作用而发生改变,不能作为好的遗传标记,但是形态特征毕竟是基因型与环境共同作用的结果,可在一定程度上反映生物遗传变异的程度,是理解物种适应机制的重要方法[2]。
香榧(Torreya grandis)属裸子植物红豆杉科(Taxaceae)香榧属(Torreya),常绿乔木,又称榧树、玉榧、野杉子,为第三纪孑遗植物,也是我国特有的珍惜树种。

苏皖4种同域分布栎树的遗传变异与基因渐渗栎属(Quercus)是壳斗科(Fagaceae)中最大的属,是典型的风媒传粉、异交植物,栎树常同域分布,种间存在着较强的基因交流,是研究植物遗传多样性、物种形成与进化、种间基因渐渗与杂交的模式植物。
中国是栎属植物的起源地,也是现代栎树的分布中心之一,栎树是我国重要经济和生态树种。
本研究以江苏省、安徽省4块样地中同域分布的麻栎(Quercus acutissima)、栓皮栎(Q. variabilis)、白栎(Q.fabri)和短柄袍栎(Q. glandulifera var. brevipetiolata)居群为研究对象,利用叶表型性状、核基因组大小和核SSR标记分析了4种栎树居群的遗传多样性、居群和个体的变异情况,以探明栎树种间基因渐渗和杂交的水平。
主要研究结果如下:(1)表型性状变异。
在种内居群水平上,麻栎的侧脉对数与叶柄长存在着显著变异,白栎除叶柄长外其它表型性状指标上均存在显著变异,短柄袍栎在叶片长、叶片长/宽、侧脉对数等3个指标上存在显著变异,栓皮栎在所有指标上均不存在显著变异。
聚类分析表明,4种栎树居群内均存在着表型变异,部分白栎、麻栎和栓皮栎个体表现出明显的种间变异表型。
(2)基因组大小变异。
通过对4种传统的细胞核分离缓冲液进行比较,并利用正交设计法对经典Marie’s缓冲液中的β-疏基乙醇和Tween 20组分进行了优化,成功构建了分别适合于麻栎、栓皮栎、白栎和短柄袍栎的流式细胞术试验体系。
并测定了江苏紫金山样地4种栎树共120个个体的核基因组大小。
发现麻栎居群与栓皮居群间在基因组大小上存在着显著变异,二者与白栎居群和短柄袍栎居群间也均存在着显著变异,但白栎居群与短柄袍栎居群间无显著变异。
聚类显示,同组的栎树间表现出广泛的种间变异,不同组的栎树间有部分个体存在着种间变异。
(3)居群遗传结构。
从来自于近缘物种的92对核SSR引物中,选出能在4种栎树中扩增出多态性的核SSR引物27对,并对4种栎树共300个个体的DNA样品进行了PCR扩增反应,进行了4种栎树总体水平、种水平和种内居群水平遗传多样性的分析。

几种国外栎属树种引种育苗试验
刘春林;曹基武;吴毅;彭重华;李百胜
【期刊名称】《林业科技开发》
【年(卷),期】2008(22)1
【摘要】栎属植物是亚热带常绿阔叶林的主要建群种.在湖南长沙中南林业科技大学植物园引种8个国外栎属树种,观察其种子发芽情况及苗木生长表现.结果表明,8个树种露地栽培均能发芽,其中弗吉尼亚栎、柳栎、南方红栎和娜塔栎在发芽率、生长发育和观赏性状等方面均优于其他4个树种,有望在城市绿化中推广应用.【总页数】3页(P78-80)
【作者】刘春林;曹基武;吴毅;彭重华;李百胜
【作者单位】中南林业科技大学,长沙,410004;中南林业科技大学,长沙,410004;中南林业科技大学,长沙,410004;中南林业科技大学,长沙,410004;中南林业科技大学,长沙,410004
【正文语种】中文
【中图分类】S7
【相关文献】
1.几种木兰科常绿树种在南京郊区引种育苗试验 [J], 朱云华;徐友新
2.6个桉树种质引种育苗试验及适应性研究 [J], 周文卫;张晓勉;向继云;高智慧;岳春雷;高大海;李贺鹏;陈斌;王泳
3.4种国外彩叶树种引种育苗试验 [J], 王济成;吕晓雪;张志成;周海涛;王程
4.几种国外主要造林树种在集安的引种表现 [J], 陆志民;谢振东
5.宝鸡市几种观赏树种生境调查及引种育苗试验 [J], 李让乐;段雯蓓;杨虎田;杨会兵;郝丽兰
因版权原因,仅展示原文概要,查看原文内容请购买。

蒙古栎常见病虫害蒙古栎常见病虫害简介蒙古栎(学名:Quercus mongolica)是一种常见的大型落叶乔木,广泛分布于我国的北方地区。
它具有较高的经济价值和生态功能,但常受到各种病虫害的威胁。
本文将介绍蒙古栎常见的病虫害,并提供相应的防治措施。
病害蒙古栎大斑病病原菌:Rhodotorula glutinis症状:病害主要表现为蒙古栎叶片上出现大型圆形斑点,斑点呈褐色,随着病情发展,斑点中心出现黑色小点,逐渐扩大并融合。
防治措施:及时清除病叶和感染的树枝,合理修剪树冠,以提高通风和光照条件,可以使用化学药剂进行喷洒防治。
蒙古栎斑点病病原菌:Phomopsis sp.症状:病害主要表现为蒙古栎叶片上出现小型圆形斑点,斑点呈褐色或灰色,具有明显的边缘,叶片逐渐干枯,发生早期脱落。
防治措施:采取合理的修剪措施,保持树冠的通风和光照,及时清除病叶和枯枝,喷洒适量的杀菌剂进行防治。
蒙古栎叶蜂蛾病原虫:Leucoptera scitella症状:叶蜂蛾的幼虫主要在蒙古栎叶片上进行取食,造成叶片大面积的骨化,形成白色的短片状斑块。
防治措施:进行适时的整形修剪,移除受害叶片,并喷洒相应的昆虫防治药剂。
虫害蒙古栎蛀幼虫病原虫:Anoplophora chinensis症状:蛀幼虫主要在蒙古栎树干内进行取食,导致树干内部出现大型蛀洞,树干表面出现树脂渗出、裂纹和破碎。
防治措施:及时进行巡查,发现受害树干及时切除,对严重受损的蒙古栎进行伐树处理,防止虫害的扩散。
蒙古栎叶蛾病原虫:Lymantria dispar症状:叶蛾的幼虫主要在蒙古栎叶片上进行取食,导致叶片被大面积啃食,出现漏食现象,极大地影响了蒙古栎的光合作用。
防治措施:喷洒适量的杀虫剂进行防治,及时清除受害的叶片。
防治措施1. 保持环境卫生:及时清除病叶、感染的树枝,减少病虫害的滋生环境。
2. 修剪树冠:合理修剪蒙古栎的树冠,提高通风和光照条件,减少病虫害的发生。

第52卷第3期东㊀北㊀林㊀业㊀大㊀学㊀学㊀报Vol.52No.32024年3月JOURNALOFNORTHEASTFORESTRYUNIVERSITYMar.20241)科技部基础资源调查专项(2019FY100505)㊂第一作者简介:李一,男,1997年11月生,东北林业大学林学院,硕士研究生㊂E-mail:li13772449244@163.com㊂通信作者:穆立蔷,东北林业大学林学院,教授㊂E-mail:mlq0417@163.com㊂收稿日期:2023年9月1日㊂责任编辑:段柯羽㊂蒙古栎叶片功能性状变化特征及影响因素1)李一㊀潘立本㊀赵雯㊀穆立蔷韩文静㊀(东北林业大学,哈尔滨,150040)㊀㊀(中国林业科学研究院林业研究所)㊀㊀㊀摘㊀要㊀为明确不同物候期叶片功能性状,揭示叶片在生长发育过程中资源分配及生态适应策略,以黑龙江小北湖自然保护区的蒙古栎(Quercusmongolica)为研究对象,分析在不同物候期其叶片功能性状特征及各功能性状与气候因子的关系㊂结果表明:蒙古栎叶片功能性状在不同物候期间均存在显著差异(P<0.05)㊂展叶期,叶片含水率㊁比叶面积㊁总酚质量分数㊁过氧化氢酶活性达到最大值;花期,叶绿素质量分数㊁类胡萝卜素质量分数㊁淀粉质量分数㊁过氧化物酶活性最高;果期,叶干质量㊁叶厚达到最大值;变色期,可溶性糖质量分数㊁总黄酮质量分数㊁超氧化物歧化酶活性最高㊂蒙古栎叶片不同功能性状间存在显著相关性㊂比叶面积与可溶性糖质量分数呈显著负相关,与淀粉质量分数呈显著正相关(P<0.05)㊂总酚质量分数与叶片含水率呈极显著正相关(P<0.01)㊂可溶性糖质量分数与淀粉质量分数呈极显著负相关(P<0.01)㊂总黄酮质量分数与叶绿素㊁类胡萝卜素质量分数呈极显著性负相关,相关系数分别为-0.946㊁-0.830;与可溶性糖质量分数呈极显著正相关,相关系数为0.816(P<0.01)㊂过氧化氢酶活性与叶片含水率㊁总酚质量分数呈极显著正相关(P<0.01)㊂月累计降水量对叶片功能性状的影响最大,贡献率为52.1%,其次为月平均温度及月累计日照时间,贡献率分别为21.8%及21.0%㊂蒙古栎叶片各功能性状间存在相互作用,且其顺应自身发育并受外部环境变化影响㊂关键词㊀蒙古栎;叶片功能性状;物候期;气候因子;冗余分析分类号㊀S792.18FunctionalTraitVariationCharacteristicsandInfluencingFactorsofQuercusmongolicaLeaf//LiYi,PanLiben,ZhaoWen,MuLiqiang(NortheastForestryUniversity,Harbin150040,P.R.China);HanWenjing(ResearchInstituteofForestry,ChineseAcademyofForestry)//JournalofNortheastForestryUniversity,2024,52(3):10-15.Toclarifythefunctionaltraitsofleavesduringdifferentphenologicalperiodsandrevealtheresourceallocationandec⁃ologicaladaptationstrategiesofleavesduringgrowthanddevelopment,theQuercusmongolicaintheXiaobeiLakeNatureReserveinHeilongjiangProvincewasselectedastheresearchobject.Thefunctionaltraitsofleavesandtheirrelationshipswithclimaticfactorswereanalyzedduringdifferentphenologicalperiods.TheresultsshowedthatthereweresignificantdifferencesinthefunctionaltraitsofQ.mongolicaleavesduringdifferentphenologicalperiods(P<0.05).Duringtheleafexpansionstage,leafmoisturepercentage,specificleafarea,totalphenoliccontent,andcatalaseactivityreachedtheirmaximumvalues.Duringthefloweringstage,chlorophyllcontent,carotenoidcontent,starchcontent,andperoxidaseac⁃tivitywerehighest.Duringthefruitingstage,leafdrymassandleafthicknessreachedtheirmaximumvalues.Duringthecolor⁃changingstage,solublesugarcontent,totalflavonoidcontent,andsuperoxidedismutaseactivitywerehighest.ThereweresignificantcorrelationsamongdifferentfunctionaltraitsofQ.mongolicaleaves.Specificleafareawassignificantlynegativelycorrelatedwithsolublesugarcontentandsignificantlypositivelycorrelatedwithstarchcontent(P<0.05).Totalphenoliccontentwashighlypositivelycorrelatedwithleafmoisturepercentage(P<0.01).Solublesugarcontentwashighlynegativelycorrelatedwithstarchcontent(P<0.01).Totalflavonoidcontentwashighlynegativelycorrelatedwithchloro⁃phyllandcarotenoidcontent,withcorrelationcoefficientsof-0.946and-0.830,respectively,andhighlypositivelycorre⁃latedwithsolublesugarcontent,withacorrelationcoefficientof0.816(P<0.01).Catalaseactivitywashighlypositivelycorrelatedwithleafmoisturepercentageandtotalphenoliccontent(P<0.01).Monthlycumulativeprecipitationhadthegreatestimpactonthefunctionaltraitsofleaves,withacontributionrateof52.1%,followedbymonthlyaveragetempera⁃tureandmonthlycumulativesunshineduration,withcontributionratesof21.8%and21.0%,respectively.Therewerein⁃teractionsamongthevariousfunctionaltraitsofQ.mongolicaleaves,andtheyrespondedtotheirowndevelopmentandwereinfluencedbyexternalenvironmentalchanges.Keywords㊀Quercusmongolica;Leaffunctionaltrait;Phenologicalperiod;Climatefactor;Redundancyanalysis㊀㊀不同生长发育时期,植物对内部资源分配及其对外界环境适应策略存在差异㊂叶片作为植物获取资源的重要器官,与外界环境接触面积大㊁对气候条件变化敏感[1]㊂因此,学者们多以叶片形态及其生理特征作为研究环境对植物影响的切入点[2]㊂有研究发现,叶龄增长对叶片结构产生重要影响,具体表现为:叶厚整体呈先变厚再变薄的趋势[3],叶片含水率及比叶面积在展叶期最大[4],叶绿素质量分数在变色期时最小[5]㊂然而,这些研究多以经济作物或灌木草本为研究对象,且聚焦于植物在单一物候期的叶片功能性状特征[4-6],缺乏全物候期叶片功能性状变化及在不同物候期叶片功能性状间差异性的研究㊂不同叶片功能性状之间存在显著相关性[7],叶片可溶性糖质量分数与淀粉呈显著负相关[8],这表明叶片内部资源存在相互转化的过程㊂当外部环境发生变化时,叶片通过差异化的资源分配来实现对环境变化的生态适应[9],且不同气候因子对植物的影响程度不同[10]㊂高温时,叶片抗氧化酶活性㊁酚类及黄酮类质量分数有所升高[11];光照增加㊁降水量减少时,植物通过叶片变厚㊁比叶面积降低的方式,最大程度保存水分,以确保其在该环境条件下维持生存[10]㊂目前,多数研究侧重于探讨单一气候因素(光照㊁温度㊁降水量等)对叶片功能性状的影响,但在自然环境中,叶片功能性状往往同时受多种气候因子的共同影响㊂因此,综合考虑不同气候因子对叶片功能性状的影响有助于更全面地理解植物对环境的适应性及其响应机制㊂蒙古栎(Quercusmongolica),又称蒙栎㊁柞树,壳斗科(Fagaceae)栎属(QuercusL.)落叶乔木,是国家二级珍贵树种,主要分布于辽宁㊁吉林㊁黑龙江等地[12]㊂蒙古栎具有耐干旱㊁耐贫瘠㊁耐寒等优良特性,树形优美,叶色变化丰富,果实独特,有较高的观赏价值㊂蒙古栎叶片提取物中含有抗炎活性的酚类物质,可用于治疗痤疮[13]㊂目前,对于该树种的研究主要集中在幼苗生长发育特征㊁光合特性和病原菌等方面,其叶片功能性状变化特征及对外部环境的适应机制尚不清楚㊂因此,我们以蒙古栎叶片为材料,测定不同物候期叶片功能性状指标,明确其变化规律,采用Pearson相关性分析探究不同功能性状间的相关关系,采用冗余分析研究气候因子对叶片功能性状的影响㊂本研究旨在明确蒙古栎在不同物候期的叶片资源分配特征及其生态适应性,为合理开发利用蒙古栎资源提供参考㊂1㊀试验区概况试验地位于黑龙江省宁安市小北湖国家级自然保护区(以下简称 小北湖保护区 ),地理坐标为128ʎ33ᶄ7ᵡ 128ʎ45ᶄ48ᵡE㊁44ʎ3ᶄ16ᵡ 44ʎ18ᶄ59ᵡN,占地面积20834hm2㊂土壤类型为暗棕色森林土,土层深厚㊂小北湖保护区属温带季风性气候,年平均气温2.5ħ,年平均降水量650mm,降水分配不均,集中在7 8月,占全年降水的60% 70%,无霜期90 100d[14]㊂2㊀研究方法2.1㊀试验材料样树选取:2022年4月 2022年10月下旬,在小北湖保护区蒙古栎天然林中,选择地势平坦㊁地力相对均匀的20mˑ20m地块,从中选取树龄为10a㊁生长状况良好且长势基本一致的10株蒙古栎作为固定取样树,进行标号㊁物候观测及叶片采集㊂物候期观测:为准确掌握植物物候现象,每7d观测1次,每次观测于上午10:00进行㊂经观测,蒙古栎物候分为4个时期,分别为展叶期㊁花期㊁果期㊁变色期㊂展叶期为枝条嫩叶露出,叶面积增大,叶片厚度逐渐加厚,第一㊁二片叶片完全平展的阶段(2022年5月4日 5月25日);花期为树体中部枝条上花序轴明显,花药饱满并有花朵开始开放的阶段(2022年6月2日 6月17日);果期为树体中部枝条上壳斗和种子逐渐增大的阶段(2022年7月15日 8月19日);变色期为树体中部叶片逐渐由绿变红再变黄,枝条由灰绿变为红褐色的阶段(2022年9月9日 2022年10月7日)㊂叶片采集:分别在展叶期(5月13日)㊁花期(6月10日)㊁果期(8月8日)㊁变色期(9月26日)进行叶片采集(图1)㊂从每株植株冠层中部的东㊁南㊁西㊁北4个方位,分别采集完整㊁无病虫害的叶片各4片,每株采集16片,共采集10株作为1个混合样,带回实验室㊂将采集好的叶片放入自封袋中,用蒸馏水冲洗干净,擦干备用㊂图1㊀不同物候期的蒙古栎叶片2.2㊀叶片功能性状指标测定本研究从形态特征㊁生理指标质量分数㊁抗氧化酶活性3个方面,共计选取了13个功能性状指标㊂叶片形态指标测定:在每个物候期采集的样品中选择30片蒙古栎叶片(3组,每组10片),用游标卡尺分别测量每组叶片上㊁中㊁下部的厚度,取3个部分的平均值为该组叶片厚度,各物候期的单叶厚度为该时期3组叶片厚度的平均值㊂使用电子天平测量并计算每组单叶鲜质量后,将样品放入烘箱105ħ杀青10min,之后在60ħ下烘48h至恒质量,测量并计算该组单叶干质量,各物候期单叶鲜质量㊁单叶干质量均为3组数据的平均值㊂用NIKONZ50相机拍摄每组叶片图像,采用ImageJ软件计算各组的总叶面积,再结合叶片数量计算单叶面积,各物候期的单叶面积为3组单叶面积的平均值㊂叶片含水率及比叶面积计算公式为:叶片含水率=[(单叶鲜质量-单叶干质量)/单叶鲜质量]ˑ100%;比叶面积=单叶面积/单叶干质量㊂叶片生理指标质量分数测定:光合色素质量分数测定采用Arnon法[15],用UV 1750型分光光度计测量665㊁649㊁470nm波长下的吸光度(Aλ)值,再依据公式计算叶绿素a㊁叶绿素b㊁类胡萝卜素质量分数㊂可溶性糖及淀粉质量分数采用蒽酮比色法测定[16]㊂11第3期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀李一,等:蒙古栎叶片功能性状变化特征及影响因素叶片总酚及总黄酮质量分数依据周香辉等[17]的方法加以修改后使用紫外分光光度计法测定㊂叶片抗氧化酶活性测定:过氧化氢酶(CAT)活性测定采用紫外吸收法,过氧化物酶(POD)活性测定采用愈创木酚法,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光还原法[18]㊂2.3㊀气候因子获取研究区气候资料观测点位为宁安市气象观测站,站点距离研究区直线距离62.9km㊂于中国历史天气网站(https://rp5.ru)获取研究区2022年5月(展叶期)㊁6月(花期)㊁8月(果期)㊁9月(变色期)的日平均气温㊁日降水量后,计算各月份平均气温和累计降水量㊂累计日照时间于中国气象资料服务中心(https://data.cma.cn/)获取㊂降水量在花期最大,果期最小;气温在果期最高,展叶期最低;各物候期日照时间相近,展叶期日照时间略高,果期略低(表1)㊂表1㊀2022年研究区各物候期累计降水量㊁累计日照时间和平均气温月份月平均气温/ħ月降水量/mm月日照时间/h513.932392.300237.724618.262460.900210.346820.157264.200183.962915.774352.200209.8852.4㊀数据处理采用单因素方差分析(One-wayANOVA)和事后多重比较(LSD)检验不同时期蒙古栎叶片指标的差异显著性;采用Pearson相关性分析检验各指标间的相关显著性;消除趋势对应分析(DCA)的辨别结果为0.29,故采用冗余分析(RDA)探究气候因子对蒙古栎叶片功能性状的影响㊂使用Excel2013㊁SPSS25.0和CANOCO5.0软件进行数据整理分析㊂3㊀结果与分析3.1㊀不同物候期蒙古栎叶片功能性状的动态变化3.1.1㊀不同物候期蒙古栎叶片形态特征的动态变化蒙古栎叶片形态在不同时期存在显著差异(P<0.05,表2)㊂展叶期时植株叶厚显著低于其他时期,叶片含水率显著高于其他时期,其他3个时期间叶厚及叶片含水率无明显差异㊂叶干质量呈先升高后降低的变化趋势,在果期时达到最大值,为0.56g㊂果期的叶干质量显著高于花期和展叶期(P<0.05),但与变色期时无显著差异㊂比叶面积从展叶期到变色期呈逐渐减小的趋势,展叶期的比叶面积显著高于其他时期,花期及果期的显著高于变色期(P<0.05),展叶期时比叶面积约是变色期的2倍㊂表2㊀不同物候期叶片形态特征物候期叶干质量/g叶厚/mm叶片含水率/%比叶面积/m2㊃kg-1展叶期(0.13ʃ0.01)c(0.21ʃ0.02)b(0.72ʃ0.00)a(25.52ʃ1.34)a花期(0.34ʃ0.01)b(0.27ʃ0.02)a(0.59ʃ0.03)b(20.02ʃ2.61)b果期(0.56ʃ0.06)a(0.30ʃ0.02)a(0.60ʃ0.01)b(17.51ʃ2.46)b变色期(0.48ʃ0.06)ab(0.27ʃ0.02)a(0.57ʃ0.03)b(12.51ʃ2.60)c㊀㊀注:表中数据为 平均值ʃ标准误 ,数据后同列不同小写字母表示差异显著(P<0.05)㊂3.1.2㊀不同物候期蒙古栎叶片生理指标的动态变化蒙古栎叶片中光合色素质量分数在不同时期存在显著差异(P<0.05),总体呈先升高后降低的趋势(表3)㊂叶绿素质量分数在花期达到最大值,为4.65mg㊃g-1,显著高于其他时期;在变色期达到最小值,为0.09mg㊃g-1,显著小于其他时期(P<0.05)㊂类胡萝卜素质量分数在花期及果期时显著高于展叶期及变色期(P<0.05),约是展叶期及变色期的3倍㊂如表3所示,可溶性糖质量分数变化总体呈先降低后升高的趋势,其在花期时最低,为98.04mg㊃g-1,变色期时显著升高,达到最高值,展叶期及果期的可溶性糖质量分数与其他物候期时差异不显著㊂淀粉质量分数变化呈先升高后降低的趋势,其在花期时达到最高值,为68.59mg㊃g-1,果期时出现显著降低,变色期的淀粉质量分数也较低㊂蒙古栎叶片总酚质量分数呈先降低后略微升高的趋势(表3)㊂展叶期总酚质量分数显著高于其他物候期(P<0.05),约是其他时期的1.5倍;花期㊁果期㊁变色期的总酚质量分数呈逐渐升高的趋势,但无显著差异㊂蒙古栎叶片总黄酮质量分数在变色期时达到最高值,为32.62mg㊃g-1,显著高于其他物候期;展叶期的总黄酮质量分数虽高于花期及果期的,但三者间差异不显著㊂表3㊀不同物候期叶片生理指标质量分数物候期叶绿素质量分数/mg㊃g-1类胡萝卜素质量分数/mg㊃g-1可溶性糖质量分数/mg㊃g-1淀粉质量分数/mg㊃g-1总酚质量分数/mg㊃g-1总黄酮质量分数/mg㊃g-1展叶期(1.67ʃ0.13)c(0.21ʃ0.03)b(115.22ʃ12.59)ab(58.72ʃ3.56)a(118.82ʃ9.85)a(23.09ʃ1.21)b花期(4.65ʃ0.07)a(0.59ʃ0.01)a(98.04ʃ12.39)b(68.59ʃ6.27)a(74.75ʃ0.28)b(17.46ʃ0.42)b果期(4.11ʃ0.02)b(0.58ʃ0.01)a(127.96ʃ4.60)ab(20.05ʃ4.93)b(87.95ʃ8.72)b(19.51ʃ0.13)b变色期(0.09ʃ0.04)d(0.18ʃ0.04)b(159.67ʃ11.50)a(20.99ʃ7.12)b(92.17ʃ5.25)b(32.62ʃ1.07)a㊀㊀注:表中数据为 平均值ʃ标准误 ,数据后同列不同小写字母表示差异显著(P<0.05)㊂21㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀东㊀北㊀林㊀业㊀大㊀学㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第52卷3.1.3㊀不同物候期蒙古栎叶片抗氧化酶活性的动态变化蒙古栎叶片过氧化物酶活性总体呈 上升-下降-上升 的趋势㊂过氧化物酶活性在花期时达到峰值,且显著高于其他物候期(P<0.05);之后下降,在果期出现显著低于其他时期的最低值,为33.33U㊃g-1㊃min-1;随后在变色期再次上升㊂超氧化物歧化酶活性仅在果期时显著低于其他时期(P<0.05),且其他时期间无显著差异㊂过氧化氢酶活性总体呈下降趋势,仅在展叶期具有较高的活性水平,且显著高于其他时期(P<0.05),约是其他时期的3倍(表4)㊂3.2㊀蒙古栎叶片不同功能性状间的相关性分析相关性分析结果表明,蒙古栎叶片不同功能性状间存在显著相关性(表5)㊂比叶面积与可溶性糖质量分数呈显著负相关,与淀粉质量分数呈显著正相关(P<0.05)㊂总酚质量分数与叶片含水率呈极显著正相关(P<0.01)㊂可溶性糖质量分数与淀粉质量分数呈极显著负相关(P<0.01)㊂总黄酮质量分数与叶绿素㊁类胡萝卜素质量分数呈极显著性负相关,相关系数分别为-0.946㊁-0.830;与可溶性糖质量分数呈极显著正相关,相关系数为0.816(P<0.01)㊂过氧化氢酶活性与叶片含水率㊁总酚质量分数呈极显著正相关(P<0.01)㊂表4㊀不同物候期叶片抗氧化酶活性物候期过氧化物酶活性/U㊃g-1㊃min-1超氧化物歧化酶活性/U㊃g-1过氧化氢酶活性/U㊃g-1㊃min-1展叶期(62.67ʃ4.69)b(292.55ʃ21.41)a(164.00ʃ9.71)a花期(84.67ʃ5.00)a(310.62ʃ11.53)a(48.40ʃ1.83)b果期(33.33ʃ7.37)c(217.01ʃ16.02)b(29.50ʃ5.32)b变色期(53.33ʃ6.66)b(324.65ʃ3.28)a(45.25ʃ9.78)b㊀㊀注:表中数据为 平均值ʃ标准误 ,数据后同列不同小写字母表示差异显著(P<0.05)㊂表5㊀叶片各功能性状间的相关系数相关因素各相关因素间的相关系数LDWLTLWCSLACHLCARLSSLSTPATFPODSODCATLT0.799∗∗1.000LWC-0.817∗∗-0.738∗∗1.000SLA-0.828∗∗-0.581∗0.866∗∗1.000CHL0.1210.421-0.1510.2691.000CAR0.3770.602∗-0.3870.0160.962∗∗1.000LSS0.5210.183-0.303-0.663∗-0.710∗∗-0.5161.000LS-0.765∗∗-0.5310.4290.676∗0.3540.149-0.777∗∗1.000TPA-0.531-0.758∗∗0.792∗∗0.426-0.536-0.673∗0.1290.0421.000TF0.165-0.176-0.162-0.548-0.946∗∗-0.830∗∗0.816∗∗-0.5180.3071.000POD-0.605∗-0.3460.1270.4010.1730.038-0.5660.854∗∗-0.208-0.2451.000SOD-0.360-0.326-0.035-0.111-0.484-0.5020.1700.363-0.0040.4550.631∗1.000CAT-0.884∗∗-0.889∗∗0.915∗∗0.739∗∗-0.326-0.548-0.2230.4990.828∗∗0.0230.2290.2151.000㊀㊀注:LDW为叶干质量;LT为叶厚;LWC为叶片含水率;SLA为比叶面积;CHL为叶绿素质量分数;CAR为类胡萝卜素质量分数;LSS为可溶性糖质量分数;LS为淀粉质量分数;TPA为总酚质量分数;TF为总黄酮质量分数;POD为过氧化物酶活性;SOD为超氧化物歧化酶活性;CAT为过氧化氢酶活性,∗表示显著相关(P<0.05);∗∗表示极显著相关(P<0.01)㊂3.3㊀蒙古栎叶片功能性状与气候因子之间的关系冗余分析前2轴的累计贡献率为80.47%,其能够解释蒙古栎叶片功能性状与气候因子之间的相关关系(图2)㊂月累计降水量对叶片功能性状的影响最明显,贡献率为52.1%,月平均温度及月累计日照时间的影响程度相近,贡献率分别为21.8%㊁21.0%(表6)㊂月累计降水量及日照时间与叶干质量㊁叶厚㊁叶绿素质量分数㊁类胡萝卜素质量分数㊁可溶性糖质量分数呈负相关,且与叶片形态指标的相关程度大;与叶片含水率㊁比叶面积㊁淀粉质量分数㊁总酚质量分数㊁总黄酮质量分数㊁过氧化物酶活性㊁超氧化物歧化酶活性㊁过氧化氢酶活性呈正相关,且与抗氧化酶活性的相关程度大㊂月平均气温与叶片功能性状之间的相关性方向与月累计降水量㊁月累计日照时间的相反㊂LDW为叶干质量;LT为叶厚;LWC为叶片含水率;SLA为比叶面积;CHL为叶绿素质量分数;CAR为类胡萝卜素质量分数;LSS为可溶性糖质量分数;LS为淀粉质量分数;TPA为总酚质量分数;TF为总黄酮质量分数;POD为过氧化物酶活性;SOD为超氧化物歧化酶活性;CAT为过氧化氢酶活性;TM为月平均温度;MS为累计降水量;MPr为累计日照时间㊂图2㊀叶片功能性状与气候因子的冗余分析结果31第3期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀李一,等:蒙古栎叶片功能性状变化特征及影响因素表6㊀气候因子对蒙古栎叶片功能性状的贡献率气候因子贡献率/%F值P月累计降水量52.110.90.002月累计日照时间21.07.00.006月平均温度21.833.70.0104 结论与讨论植物在生长发育各阶段,叶片的形态和生理指标均有较大差异,这与外界环境条件和叶片结构建成资源投入有关[19]㊂叶厚度㊁叶干质量㊁叶片含水率㊁比叶面积等形态指标能够反映叶片生长状况及植物获取及利用资源的能力[20]㊂本研究中,叶片含水率㊁比叶面积在展叶期达到最大值[21],且展叶期与其他时期时存在显著差异㊂展叶期以营养生长为主,叶片代谢速率高,故含水率高[4],同时,该时期叶片迅速展开,叶面积增大,因此比叶面积大㊂展叶期完成后,叶片面积增速减缓,而骨架支撑结构[22]及营养物质增多[23],所以比叶面积开始减小㊂在荒漠植物叶片(合头草(Sympegmaregelii)㊁荒漠锦鸡儿(Caraganaroborovskyi)等)的研究中,为减少水分散失,细胞扩张由纵向转为横向,导致叶片厚度增大[24]㊂本研究中果期气温最高,植物蒸腾作用最强,故叶厚值最大㊂此外,果期时叶干质量最大,该时期叶片需要为果实的形成准备充足的营养物质[25]㊂在叶片生理指标方面,光合色素在花㊁果期时质量分数高,在变色期时质量分数急剧下降到最低㊂受植物体养分回收策略等因素的影响[26],变色期光合色素的分解速率大于合成速率,因此叶片内光合色素质量分数降低㊂可溶性糖质量分数在花期时最低,这是因为花芽的分化和成花过程会消耗大量的可溶性糖[27]㊂可溶性糖质量分数在变色期时最大,这与唐生森等[28]的研究不一致,其原因是该时期淀粉会转化为可溶性糖以调解渗透压来抵御低温[8]㊂淀粉质量分数在果期时最低,原因是该时期淀粉分解,以为果实的生长发育提供能量[29]㊂总酚质量分数在展叶期时最高,之后降低[30]㊂变色期蒙古栎叶色逐渐发生转变,黄酮类代谢途径增强,总黄酮质量分数出现显著升高[31],并达到最大值㊂叶片抗氧化酶活性总体呈一定的协同趋势㊂然而,与红树莓叶片研究结果不同[32],本研究中3种抗氧化酶活性在果期时最低,这说明该时期蒙古栎叶片内部环境稳定㊂蒙古栎叶片内部环境稳定是由于叶片生长已达到一定程度,生长速率减缓,且果期时叶片受外界环境胁迫程度相对较小㊂展叶期蒙古栎叶片中过氧化氢酶活性最高且与其他时期存在显著差异,这是因为展叶期叶片生长迅速,细胞内自由基代谢失衡,从而诱导酶活性上升,以维持膜系统在生长发育中的正常功能[33]㊂植物叶片生长发育过程中,各功能性状间相互影响,以形成特定的资源分配和生态适应策略[34]㊂研究发现,叶片功能性状在不同类别(形态特征㊁生理指标㊁抗氧化酶活性)间存在显著相关性㊂比叶面积与可溶性糖㊁淀粉质量分数均存在显著相关性,这是因为可溶性糖和淀粉作为主要非结构性碳水化合物参与了叶片骨架的构建[22],从而影响叶片形态㊂叶片含水率高时,叶片生长迅速,产生大量过氧化氢,此时需要过氧化氢酶㊁总酚等抗氧化物质及时清除过氧化氢以维持稳态[11],因此,过氧化氢酶活性及总酚质量分数与叶片含水率呈显著正相关㊂此外,叶片功能性状指标间也存在显著相关性,其中,叶厚㊁叶干质量与比叶面积呈显著负相关㊂在程雯等[35]对喀斯特高原峡谷植物的研究中可以看出,植物将更多干物质投入叶片构建以提高抗逆性㊂可溶性糖与淀粉之间相互转化[8],两者存在显著负相关关系㊂总黄酮与叶绿素㊁类胡萝卜素㊁可溶性糖质量分数间存在显著相关性,这验证了总黄酮质量分数与叶色变化息息相关[36]㊂本研究不仅探究了叶片功能性状间的内在关联,还为深入了解植物在生长发育过程中的内部资源分配提供了重要线索,有助于更全面地理解蒙古栎等植物的生态适应策略㊂叶片作为植物地上部分获取资源的主要器官,长期暴露在外界环境中,其功能性状也受环境因子影响㊂本研究中,月累计降水量对叶片功能性状的贡献率最大,为52.1%,这是因为降水量过高会造成水涝胁迫,抑制植物生长,导致叶片厚度㊁叶干质量有所下降㊂水涝胁迫导致植物体内活性氧等物质升高,使抗氧化酶活性有所提高[11],因此,叶片含水率与过氧化氢酶活性㊁总酚质量分数呈显著正相关㊂平均气温及日照时间对叶片功能性状的贡献率相对较小,这是因为研究区内平均气温和日照时间在叶片生长发育阶段(5 9月)相对稳定,植物生长受温度胁迫和光胁迫的程度较小㊂研究认为,降水量是影响蒙古栎叶片功能性状的关键气候因子㊂叶片功能性状之间的相互关系不仅适用于植物在不同生长发育阶段对内部资源分配的调整,还适用于气候环境对其的影响㊂本研究中,平均气温与叶厚㊁叶干质量㊁比叶面积显著相关,这与槭树等应对夏季高温时的生态策略一致[37]㊂深入研究气候因子对叶片功能性状的影响有助于全面理解植物在各种环境条件下的生态适应策略㊂本研究为预测气候变化及极端天气事件对蒙古栎的影响提供了参考,为及时制定41㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀东㊀北㊀林㊀业㊀大㊀学㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第52卷相应的人为干预措施以实现对植物管理和保护奠定了基础㊂本研究从叶片形态特征㊁生理指标质量分数㊁抗氧化酶活性3个方面选取了13个指标,通过对各指标进行测定分析,发现蒙古栎叶片功能性状在不同物候期间存在显著差异㊂展叶期,叶片含水率㊁比叶面积㊁总酚质量分数㊁过氧化氢酶活性达到最大值;花期,叶绿素㊁类胡萝卜素㊁淀粉质量分数㊁过氧化物酶活性最高;果期,叶干质量㊁叶厚达到最大值;变色期,可溶性糖㊁总黄酮质量分数㊁过氧化物歧化酶活性最高㊂研究进一步探究了植物自身及外界环境对叶片功能性状的影响,发现蒙古栎叶片的不同功能性状间相互影响且降水量是影响叶片功能性状的关键气候因子㊂参㊀考㊀文㊀献[1]㊀徐倩,李华雄,黄文沛,等.GA3和Spd对杜鹃开花期光合特性和抗氧化系统的影响[J].热带亚热带植物学报,2019,27(4):439-445.[2]㊀黄庆阳,谢立红,曹宏杰,等.五大连池火山山杨叶功能性状的变异特征[J].北京林业大学学报,2021,43(2):81-89.[3]㊀SCHMITTS,TRUEBAS,COSTES,etal.Seasonalvariationofleafthickness:anoverlookedcomponentoffunctionaltraitvaria⁃bility[J].PlantBiology,2022,24(3):458-463.[4]㊀彭仲韬,郭嘉兴,王艺璇,等.不同生长期小兴安岭3种槭树叶性状变异及相关性分析[J/OL].南京林业大学学报(自然科学版),2023:1-12[2023-05-31].https://kns.cnki.net/kcms2/detail/32.1161.s.20230530.1431.004.html.[5]㊀赖玖鑫.枫香(Liquidambarformosanahance.)叶色季节性变化及常彩叶机理研究[D].北京:中国林业科学研究院,2020.[6]㊀尚小娇,王迪海.不同物候期枣树细根和叶片可溶性糖㊁蛋白和C㊁N㊁P含量的变化[J].园艺学报,2016,43(11):2243-2250.[7]㊀李琴,王磊,张雪,等.不同种植年限中间锦鸡儿叶片功能性状的差异性研究[J].林业科学研究,2023,36(4):173-182.[8]㊀王睿照,毛沂新,云丽丽,等.氮添加对蒙古栎叶片碳氮磷化学计量与非结构性碳水化合物的影响[J].生态学杂志,2022,41(7):1369-1377.[9]㊀WANGCY,ZHOUJW,XIAOHG,etal.VariationsinleaffunctionaltraitsamongplantspeciesgroupedbygrowthandleaftypesinZhenjiang,China[J].JournalofForestryResearch,2017,28(2):241-248.[10]㊀杨永,黄玥,卫伟.西北旱区典型灌草叶片性状沿气候梯度的变化[J].生态学杂志,2021,40(12):3769-3777.[11]㊀YANGJ,CUIJY,CHENJX,etal.Evaluationofphysico⁃chemicalpropertiesinthreeraspberries(Rubusidaeus)atfiveripeningstagesinNorthernChina[J].ScientiaHorticulturae,2020,263.doi:10.1016/j.scienta.2019.109146.[12]㊀李春明,赵丽芳,李利学.基于混合效应模型和零膨胀模型方法的蒙古栎林分水平枯损模型[J].林业科学,2019,55(11):27-36.[13]㊀KIMM,YINJ,HWANGIH,etal.Anti⁃acneVulgariseffectsofpedunculaginfromtheleavesofQuercusmongolicabyanti⁃in⁃flammatoryactivityand5α⁃reductaseinhibition[J].Molecules,2020,25(9).doi:10.3390/molecules25092154.[14]㊀项凤影,孙继旭,关艳辉,等.黑龙江小北湖国家级自然保护区外来入侵植物研究[J].安徽农业科学,2023,51(3):136-138,146.[15]㊀ARNONDI.Copperenzymesinisolatedchloroplasts.polyphe⁃noloxidaseinbetavulgaris[J].PlantPhysiology,1949,24(1):1-15.[16]㊀董涵君,王兴昌,苑丹阳,等.温带不同材性树种树干非结构性碳水化合物的径向分配差异[J].植物生态学报,2022,46(6):722-734.[17]㊀周香辉,莫晓宁,凌楠,等.广西产 桂桑优 桑叶不同溶剂提取物的抗氧化及降糖活性分析[J].食品科技,2022,47(12):193-199.[18]㊀李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.[19]㊀RENLA,HUANGYM,PANYP,etal.Differentialinvest⁃mentstrategiesinleafeconomictraitsacrossclimateregionsworldwide[J].FrontiersinPlantScience,2022,13.doi:10.3389/fpls.2022.798035.[20]㊀朱济友,于强,徐程扬,等.植物功能性状及其叶经济谱对硬化地表的响应[J].农业机械学报,2019,50(3):204-211.[21]㊀姚婧,李颖,魏丽萍,等.东灵山不同林型五角枫叶性状异速生长关系随发育阶段的变化[J].生态学报,2013,33(13):3907-3915.[22]㊀NIINEMETSÜ,PORTSMUTHA,TENAD,etal.Doweunder⁃estimatetheimportanceofleafsizeinplanteconomics?dispropor⁃tionalscalingofsupportcostswithinthespectrumofleafphysiog⁃nomy[J].AnnalsofBotany,2007,100(2):283-303.[23]㊀RÖMERMANNC,BUCHERSF,HAHNM,etal.Plantfunc⁃tionaltraits⁃fixedfactsorvariabledependingontheseason?[J].FoliaGeobotanica,2016,51(2):143-159.[24]㊀李善家,苏培玺,张海娜,等.荒漠植物叶片水分和功能性状特征及其相互关系[J].植物生理学报,2013,49(2):153-160.[25]㊀高鹏,南志标,吴永娜,等.新疆阿勒泰地区8种生态型罗布麻光合生理及生长特性研究[J].西北植物学报,2015,35(10):2069-2077.[26]㊀BRANTAN,CHENHYH.Patternsandmechanismsofnutri⁃entresorptioninplants[J].CriticalReviewsinPlantSciences,2015,34(5):471-486.[27]㊀董晓晓,别沛婷,袁涛.3个牡丹品种花芽分化过程形态及叶片碳水化合物质量分数变化[J].东北林业大学学报,2020,48(7):34-39.[28]㊀唐生森,陈虎,覃永康,等.枫香秋季变色期叶色变化及其生理基础[J].广西植物,2021,41(12):2061-2068.[29]㊀赵世荣,齐延巧,周伟权,等. 冬杏 果实和叶片中可溶性总糖㊁淀粉含量与淀粉酶活性间的相关性研究[J].新疆农业大学学报,2016,39(5):353-359.[30]㊀MIKULICPM,STAMPARF,VEBERICR.Seasonalchangesinphenoliccompoundsintheleavesofscab⁃resistantandsuscepti⁃bleapplecultivars[J].CanadianJournalofPlantScience,2009,89(4):745-753.[31]㊀SHENJZ,ZOUZW,ZHANGXZ,etal.Metabolicanalysesrevealdifferentmechanismsofleafcolorchangeintwopurple⁃leafteaplant(CamelliasinensisL.)cultivars[J].HorticultureRe⁃search,2018,5.doi:10.1038/s41438-017-0010-1.[32]㊀范青,李程,李明,等.红树莓叶片生长代谢过程中酚类物质含量及抗氧化酶活性分析[J].食品科学,2023,44(8):238-246.[33]㊀WRIGHTIJ,ACKERLYDD,BONGERSF,etal.Relation⁃shipsamongecologicallyimportantdimensionsofplanttraitvaria⁃tioninsevenneotropicalforests[J].AnnalsofBotany,2007,99(5):1003-1015.[35]㊀程雯,喻阳华,熊康宁,等.喀斯特高原峡谷优势种叶片功能性状分析[J].广西植物,2019,39(8):1039-1049.[36]㊀GEW,WANGXX,LIJY,etal.RegulatorymechanismsofleafcolorchangeinAcerpictumsubsp.mono[J].Genome,2019,62(12):793-805.[37]㊀刘阳,许丽颖,魏统超,等.4种槭树叶功能性状及其关系对季节变化的响应[J].植物研究,2023,43(2):242-250.51第3期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀李一,等:蒙古栎叶片功能性状变化特征及影响因素。
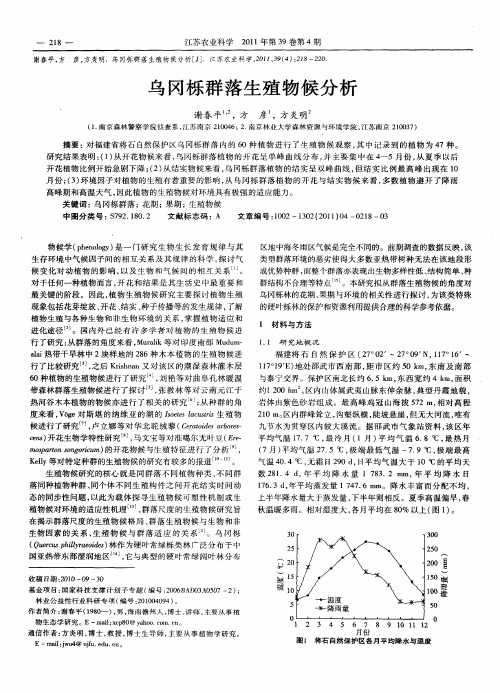
㊀Guihaia㊀Mar.2024ꎬ44(3):488-500http://www.guihaia-journal.comDOI:10.11931/guihaia.gxzw202211057林毅喆ꎬ朱艺耀ꎬ付厚华ꎬ等ꎬ2024.闽南花岗岩堡状峰乌冈栎种群的表型变异[J].广西植物ꎬ44(3):488-500.LINYZꎬZHUYYꎬFUHHꎬetal.ꎬ2024.PhenotypicvariationofQuercusphillyreoidespopulationsoncastlepeakofSouthFujianꎬChina[J].Guihaiaꎬ44(3):488-500.闽南花岗岩堡状峰乌冈栎种群的表型变异林毅喆1ꎬ朱艺耀2ꎬ付厚华1ꎬ马㊀良3ꎬ陈世品1∗(1.福建农林大学林学院ꎬ福州350002ꎻ2.云霄县林业局ꎬ福建漳州363399ꎻ3.福建卫生职业技术学院ꎬ福州350101)摘㊀要:堡状峰是类似城堡外貌而凸出于周围环境之上的特殊山峰ꎬ位于中国闽南乌山风景区堡状峰地貌上的乌冈栎(壳斗科)种群的表型发生了变异ꎬ其与原类型的显著不同在于叶背具宿存性黄褐色星状毛ꎮ为解释这一现象ꎬ该研究对堡状峰顶部的乌冈栎种群分布进行调查ꎬ包括其中的变异型㊁原始型㊁过渡型ꎬ以及堡状峰山脚㊁另一异域的典型原始型种群被研究ꎬ基于叶表皮解剖ꎬ利用方差分析统计显微性状的差异ꎬ结合生态因子ꎬ探究变异的原因及演化的可能途径ꎮ结果表明:(1)变异型在山顶西侧形成以其为优势种的常绿阔叶灌丛群落ꎬ东侧的小片灌丛以原始型为主ꎬ后者内部还有少量过渡型ꎮ(2)各种群单细胞毛状体基(STB)与复合毛状体基(CTB)在叶上表皮的密度很小甚至小于每平方毫米1个ꎬ宏观的表现为无毛ꎮ原始型㊁过渡型㊁变异型的STB密度均值在叶下表皮递增ꎮ变异型种群与原始种群相比ꎬ叶下表皮CTB或典型星状毛(Tst)的密度由每平方毫米0.481~1.122个变异至每平方毫米57.2个ꎬ宏观表现为形成密毛ꎮ气孔长宽比在原始型(1.187~1.205)至变异型(1.258)间递增ꎮ(3)方差分析结果显示ꎬ变异型叶下表皮STB与CTB密度及气孔长宽比较原始型显著增加(大)(P<0.01)ꎬ但气孔密度与尺寸在种群间未表现出规律与差异性(P>0.05)ꎮ乌山堡状峰顶的土壤贫瘠㊁水热条件差ꎮ综合生境与统计的结果表明ꎬ变异型乌冈栎叶背毛状体及气孔长宽比的变异具抵抗干旱或极端气温的功能ꎮ由于花粉流未曾间断ꎬ因此相比于地理隔离ꎬ特殊生境促使乌冈栎发生适应性演化应是该变异产生的主要原因ꎮ该研究为亚热带特殊花岗岩地貌促进种群分化和物种形成提供了直接的证据ꎮ关键词:陆地岛屿系统ꎬ乌冈栎ꎬ叶显微性状ꎬ自然选择ꎬ种群分化中图分类号:Q944㊀㊀文献标识码:A㊀㊀文章编号:1000 ̄3142(2024)03 ̄0488 ̄13PhenotypicvariationofQuercusphillyreoidespopulationsoncastlepeakofSouthFujianꎬChinaLINYizhe1ꎬZHUYiyao2ꎬFUHouhua1ꎬMALiang3ꎬCHENShipin1∗(1.CollegeofForestryꎬFujianAgricultureandForestryUniversityꎬFuzhou350002ꎬChinaꎻ2.YunxiaoCountyForestryBureauꎬZhangzhou363399ꎬFujianꎬChinaꎻ3.FujianHealthCollegeꎬFuzhou350101ꎬChina)Abstract:Castlepeakisamountainthatresemblestheappearanceofacastleandprotrudesaboveitssurroundings.ThephenotypesofQuercusphillyreoides(Fagaceae)populationslocatedoncastlepeakintheWushanScenicSpotofsouthern收稿日期:2023-04-11㊀接受日期:2023-08-22基金项目:国家重点研发计划项目(2019YFD1000400)ꎻ君子峰国家级自然保护区维管植物资源调查项目(闽财资环指[2019]15号)ꎻ中医药公共卫生服务补助专项(财社[2019]39号)ꎮ第一作者:林毅喆(1999 )ꎬ硕士研究生ꎬ主要从事生物多样性研究ꎬ(E ̄mail)linyizhe_0220@163.comꎮ∗通信作者:陈世品ꎬ博士ꎬ教授ꎬ主要从事植物分类学研究ꎬ(E ̄mail)fjcsp@126.comꎮFujianꎬChinaꎬarevariegated.Itdifferssignificantlyfromtheoriginalforminthepresenceofpersistentyellow ̄brownstellatehairsontheleafabaxialsurface.ToexplainthisphenomenonꎬthedistributionofQ.phillyreoidespopulationsatthesummitofcastlepeakwasinvestigatedꎬwherevariantꎬoriginalꎬandintermediateformsꎬaswellastypicaloriginalpopulationsatthefootofcastlepeakandanotherallopatricareaꎬwerestudied.DifferencesinmicromorphologicaltraitswerecountedusingANOVAbasedonleafepidermalanatomy.Ecologicalfactorswerelinkedwithaviewtodiscoveringthecausesofvariationandpossiblepathwaysofevolution.Theresultswereasfollows:(1)Thevariantformformedanevergreenbroad ̄leavedscrubcommunitywithitasthedominantspeciesonthewestsideofthesummitꎬandthesmallpatchofscrubontheeastsideisdominatedbytheoriginalformꎬwithafewintermediateformindividualswithinthelatter.(2)Thedensitiesofsingletrichomebase(STB)andcompoundtrichomebase(CTB)intheleafadaxialepidermisofvariouspopulationswasveryloworevenlessthan1No. mm ̄2ꎬandthemacroscopicexpressionwasglabrous.ThemeanvaluesofSTBdensityoforiginalꎬintermediateꎬandvariantformswereincreasingintheleafabaxialepidermis.ThemeandensityofCTBortypicalstellatetrichome(Tst)ontheleafabaxialepidermisvariedfrom0.481-1.122No. mm ̄2to57.2No. mm ̄2inthevariantformpopulationcomparedwiththeoriginalformpopulationsꎬwhichmacroscopicallyshowedtheformationofdensehairs.Thestomatallength ̄widthratioincreasesfromoriginalform(1.187-1.205)tovariantform(1.258).(3)ANOVAresultsshowedthatvariantformleafabaxialepidermalSTBꎬCTBdensityandstomatallength ̄widthratioweresignificantlyincreased(larger)thantheoriginalform(P<0.01)ꎬbutstomataldensityandsizedidnotshowregularityandvariabilityamongpopulations(P>0.05).PoorsoilꎬwaterandheatconditionsatthesummitoftheWushancastlepeakꎬandcombiningthebackgroundwiththeobtainedresultsindicatethatthevariationinthevariantQ.phillyreoidesleafabaxialtrichomesandstomatalaspectratiowereresistanttodroughtortemperatureextremes.SincepollenflowhasnotbeeninterruptedꎬtheadaptiveevolutionofQ.phillyreoidesduetospecifichabitatsshouldbethemainreasonforthisvariationcomparedtogeographicalisolation.Themicroscopictraitsshowednosignificantdifferences(P>0.05)betweentheoriginalformpopulationonthetopofcastlepeakandtherestoriginalformpopulationsconfirmsthatthevariationhasbeenfixedbylong ̄termselectionandtheshort ̄termenvironmentalinfluencesplayalimitedroleinshapingthevarianttraits.Thisstudyprovidesdirectevidencethatsubtropicalexceptionalgraniticlandformspromotepopulationdifferentiationandspeciesformation.Keywords:continentalislandsystemꎬQuercusphillyreoidesꎬleafmicroscopiccharacterꎬnaturalselectionꎬpopulationdifferentiation㊀㊀花岗岩堡状峰(castlepeak)(下称堡状峰)是四周由陡直崖壁围限呈城堡状的一种花岗岩地貌类型(梁诗经和文斐成ꎬ2010)ꎬ可连绵成墙状山体ꎮ中国东南沿海的堡状峰以新近纪(Neogene)后地质构造发生中度抬升为基础ꎬ经化学风化㊁流水下切侵蚀塑造而来(崔之久等ꎬ2007)ꎮ堡状峰实际为介于石林(stoneforest)与石蛋(pebbly)之间的一种花岗岩过渡地貌类型ꎮ其特点为四周陡峻ꎬ岩壁裸露ꎬ峰顶平坦ꎬ可沉积土壤ꎮ较周边丘陵ꎬ堡状峰突兀地伫于山间ꎬ生态特征与低地截然不同(Porembskiꎬ2007)ꎮ热带地区的另一种花岗岩地貌 岛山 (inselberg)与堡状峰相似ꎬ但区别在于前者构造抬升轻微ꎬ经化学风化在平原上形成孤立的岩石露头(崔之久等ꎬ2007)ꎮ岛山上的植物群落常与周边地区不同ꎬ被认为是孤立山体特殊生境产生的效果ꎻ同时ꎬ其孤立程度取决于周围植被类型ꎬ较于真实岛屿ꎬ生态隔离弥补了岛山之间地理距离相对较短的不足(Porembskiꎬ2007)ꎮ生态与距离隔离以及不同于山脚的特殊生境共同作用于岛山的物种演化ꎮ就物种多样性而言ꎬ有研究显示其他因素(如面积㊁环境筛)而非隔离距离与物种丰富度间有更大关联(Conceiçãoetal.ꎬ2007ꎻHenneronetal.ꎬ2019)ꎬ表明在较小空间尺度内ꎬ在异质性生境下发生了适应性辐射ꎮ大量研究表明ꎬ一些植物种群形态与遗传结构在或远或近的岛山上发生变异ꎬ基因流发生不同程度的限制ꎬ种群内遗传多样性降低ꎬ促进了种群分化与物种形成(Barbaráetal.ꎬ2008ꎻByrneetal.ꎬ2019ꎻFelicianoetal.ꎬ2022)ꎮ岛山作为陆地岛屿系统用于观察生态现象与物种形成ꎬ在温带与热带受到长期的关注9843期林毅喆等:闽南花岗岩堡状峰乌冈栎种群的表型变异(Porembski&Barthlottꎬ2012)ꎮ中国拥有不同的地质史ꎬ喜马拉雅山脉㊁阿尔泰-天山地区㊁中国西南山地这类称作 天空之岛 (空岛ꎬskyislands)的陆地岛屿近年的关注度较高ꎬ其中地形地貌及其改变与随之产生的气候变迁对谱系地理结构的影响研究较多(He&Jiangꎬ2014)ꎮ西南地区的盆地㊁峡谷在更新世冰川期为物种提供相互间隔的小生境避难所ꎬ这些避难所便成为遗传分化的中心(Xieetal.ꎬ2012ꎻYangZYetal.ꎬ2012)ꎬ青藏高原的抬升还让异地分化的物种二次相遇发生偶然的杂交种化事件(YangFSetal.ꎬ2012ꎻZhangetal.ꎬ2019)ꎬ现有证据表明空岛上的高寒植物共享了一连串地质运动引发的谱系地理结构与进化历史(Luoetal.ꎬ2016)ꎮ然而ꎬ在中国亚热带的花岗岩堡状峰㊁峰丛㊁峰墙㊁石林等地貌上开展有关进化与生态的研究甚为少见ꎮ狭义栎属(Quercus)全世界约300种ꎬ中国约51种㊁14变种㊁1变型(中国科学院中国植物志编辑委员会ꎬ1998)ꎬ由于复杂的网状进化历史ꎬ具有极高物种多样性(Hippetal.ꎬ2020)ꎬ栎属植物受地理与古气候的影响在各地发生适应性演化(Yangetal.ꎬ2021)ꎬ叶表皮性状常视为重要特征之一被用于种间界定(Dengetal.ꎬ2017)ꎮ2019年11月ꎬ我们在福建省南部漳州市云霄和诏安两县交界的乌山风景区开展全国中药资源普查(图1:AꎬC)ꎬ于乌山主峰红旗岩(Hongqiyan)顶发现一叶柄㊁叶背显著被星状毛的乌冈栎(Quercusphillyreoides)群体(其余性状未表现出显著不同)ꎮ经过1年的持续观察ꎬ发现其成熟叶片毛被不脱落ꎮ乌冈栎是中国秦岭淮河以南广泛分布的壳斗科(Fagaceae)栎属冬青砾组(SectionIlex)常绿阔叶树ꎬ日本也有分布ꎬ新叶有时疏被毛ꎬ老叶无毛或仅叶背中脉被疏柔毛(中国科学院中国植物志编辑委员会ꎬ1998)ꎮ对此ꎬ我们推测该种群叶表皮特征或多或少发生变异ꎬ故对乌山风景区堡状峰山顶㊁山脚及另一异域的乌冈栎种群做显微形态的比较研究ꎬ通过叶解剖获取表皮性状ꎬ所得性状数据经方差分析检验与比较ꎬ拟探讨以下问题:(1)变异种群哪些叶表皮性状发生了变异ꎬ变异的规律与趋势如何ꎻ(2)变异产生与堡状峰生境的关系ꎬ以及其是否具备生态合理性ꎻ(3)种群分化的历史进程ꎬ以及其是否具备演化合理性ꎮ通过对以上问题的探讨ꎬ以期为亚热带特殊花岗岩地貌促进种群分化提供证据ꎮ1㊀材料与方法1.1研究样品采集样品之前ꎬ我们查阅了乌冈栎模式标本及中国数字标本馆(ChineseVirtualHerbariumꎬCVHꎬhttps://www.cvh.ac.cn/)的所有普通标本(有照片的标本)ꎬ2份模式标本(CavalerieꎬJ.3626)均为叶无毛被类型ꎬ209份普通标本中有1份采自广西的标本与我们在红旗岩顶所见相似(叶背显著具毛ꎬ陈照宙50295)ꎬ采集签记录的生境㊁生活型亦类同ꎮ然而ꎬ中国分布之北界未见该性状出现ꎬ表明叶背具稳定毛被的性状不是环境直接作用的结果ꎬ亦非乌冈栎表型可塑性可简单解释ꎮ与此同时ꎬ我们在红旗岩顶不但发现叶背具毛的种群ꎬ典型原始型亦有分布ꎮ为了解释这些现象ꎬ我们采集了红旗岩顶原始型㊁变异型与假定的过渡型ꎬ以及红旗岩脚原始型的叶片与凭证标本ꎮ由表1可知ꎬ野外观察到的过渡型是相对原始型叶背的几乎完全无毛以及变异型的叶背几乎完全为毛状体覆盖(叶下表皮不可见)而言ꎬ即过渡型虽然拥有宿存性的毛ꎬ但其叶下表皮在全株均明显可见ꎬ介于原始型和变异型两者之间ꎮ由于原始型的稳定性ꎬ允许我们以任意异域环境的乌冈栎种群代表全生境范围的所有乌冈栎群体ꎬ来说明红旗岩顶的这种变异ꎮ5个(极小)种群被研究ꎬ每种群随机选择3个成年个体(个体间大于等于20m或最大间隔距离)ꎬ每个体东西南北重复取样(共4片)ꎻ考虑到毛状体对气孔性状的遮盖将不利于后期观察与统计ꎬ对变异型每个方向额外采集1片叶子ꎬ利用解剖刀去除毛被ꎮ叶片被切割成叶尖㊁叶中㊁叶基3段ꎬ保存于FAA固定液(38%福尔马林ʒ冰醋酸ʒ70%乙醇=1ʒ1ʒ18ꎬ体积比)中ꎮ样品采集在2d内完成ꎬ采集时间和天气均相同ꎮ叶表皮切片的制作方法参考Deng等(2017)ꎮ1.2研究地概况漳州乌山主体位于福建南部与广东交界的诏安㊁云霄两县境内ꎬ为东南海岸平原㊁东北漳江㊁西南东溪环绕ꎻ南北走向ꎬ南北约45kmꎬ东西约20kmꎬ面积约900km2ꎮ乌山风景区位于乌山中部ꎬ主峰海拔1050.6mꎬ山体四周悬崖绝壁ꎬ稍有棱角ꎬ顶部平坦ꎬ与附近山谷高差达215m(图1:A)ꎮ乌山基岩为花岗岩ꎬ新近纪后局部构造中度抬升094广㊀西㊀植㊀物44卷表1㊀种群与叶片样品凭证信息Table1㊀Voucherinformationofpopulationsandleafsamples编号Number种群类型Populationform凭证标本Voucher采集信息Collectioninformation1变异型VariantformFJFC20210904005(FJFC)红旗岩顶ꎬ乌山风景区ꎬ诏安ꎬ福建ꎬ中国ꎮ117ʎ10ᶄ26.97ᵡEꎬ23ʎ55ᶄ16.75ᵡNꎬ海拔1032mꎬ2021年9月4日TopoftheHongqiyanꎬWushanScenicAreaꎬZhao anꎬFujianꎬChina.117ʎ10ᶄ26.97ᵡEꎬ23ʎ55ᶄ16.75ᵡNꎬalt.1032mꎬ4Sep.20212过渡型IntermediateformFJFC20211212004(FJFC)红旗岩顶ꎬ与种群1同地ꎮ117ʎ10ᶄ27.29ᵡE㊁23ʎ55ᶄ16.9ᵡNꎬ海拔1028mꎬ2021年12月12日TopoftheHongqiyanꎬco ̄locatedwithPopulation1.117ʎ10ᶄ27.29ᵡEꎬ1.23ʎ55ᶄ16.9ᵡNꎬalt.1028mꎬ12Dec.20213原始型1Originalform1FJFC20211212001(FJFC)红旗岩顶ꎬ与种群1同地ꎮ117ʎ10ᶄ26.86ᵡE㊁23ʎ55ᶄ16.59ᵡNꎬ海拔1028mꎬ2021年12月12日TopoftheHongqiyanꎬco ̄locatedwithPopulation1.117ʎ10ᶄ26.86ᵡEꎬ1.23ʎ55ᶄ16.59ᵡNꎬalt.1028mꎬ12Dec.20214原始型2Originalform2FJFC20211212005(FJFC)红旗岩脚ꎬ与种群1同地ꎮ117ʎ10ᶄ40.18ᵡE㊁23ʎ55ᶄ16.82ᵡNꎬ海拔918mꎬ2021年12月12日FootoftheHongqiyanꎬco ̄locatedwithPopulation1.117ʎ10ᶄ40.18ᵡEꎬ1.23ʎ55ᶄ16.82ᵡNꎬalt.918mꎬ12Dec.20215原始型3Originalform3FJFC20211209005(FJFC)山谷ꎬ上坡ꎬ青云山峡谷瀑布风景区ꎬ永泰ꎬ福建ꎬ中国ꎮ119ʎ0ᶄ28.39ᵡEꎬ25ʎ45ᶄ40.01ᵡNꎬ海拔818mꎬ2021年12月9日ValleyꎬuphillꎬQingyunMountainCanyonWaterfallScenicSpotꎬYongtaiꎬFujianꎬChina.119ʎ0ᶄ28.39ᵡEꎬ25ʎ45ᶄ40.01ᵡNꎬalt.818mꎬ9Dec.2021(图1:B)ꎮ乌山主峰红旗岩属幼年(juvenilestage)至青年(adolescencestage)发育期花岗岩地貌(Chenetal.ꎬ2009)ꎬ即为典型的堡状峰(图1:C)ꎮ山顶保留中新世统一夷平面的残留部分ꎬ山脚堆积深厚风化壳ꎬ发育成红壤(崔之久等ꎬ2007)ꎮ亚热带季风气候ꎬ年均降水1519.6mmꎬ夏季均降水624mmꎬ冬季均降水121mmꎻ白天均温27ħꎬ夜间均温20ħꎬ夏季最高温31ħꎬ冬季最低温12ħꎮ地带植被为常绿阔叶林ꎬ壳斗科㊁樟科(Lauraceae)㊁山茶科(Theaceae)等为该地带森林的优势类群(吴中伦和蒋有绪ꎬ1997)ꎮ1.3统计和分析方法利用BK5000型光学显微镜观察叶上下表皮细胞垂周壁㊁毛状体基㊁毛状体及气孔ꎮ叶表皮术语及特征描述参考Deng等(2017)ꎮ每切片选1个视野(200ˑ)拍摄记录ꎮ用ImageJ1.53f51(https://imagej.nih.gov/ij/index.html)计测视野内全部气孔的数目ꎬ并测量尺寸(保卫细胞外轮廓的长径及短径)(Arenaetal.ꎬ2017)ꎮ毛状体/毛状体基数目由人工在每个视野中计数得到ꎮ随后气孔㊁毛状体/毛状体基数目转化为密度ꎬ气孔尺寸计算出长宽比(长径与短径的比值)ꎮ将每片叶子叶尖㊁叶中㊁叶基各性状的数据求平均ꎬ以排除随机误差ꎮ由于4个方向的测量值并无明显的规律ꎬ因此将其全部纳入原始数据集ꎬ以减小抽样个体数较少可能存在的系统误差ꎮ用SPSS25对原始数据集各性状实施F检验与多重比较(LSD)ꎬ对结果具显著性的性状绘制箱型图ꎮP<0.01ꎬ认为差异极显著ꎻP<0.05ꎬ则认为差异显著ꎮ2㊀结果与分析2.1表型变异和空间分布变异种群生于红旗岩顶的花岗岩风化壳上(图2:A-D)ꎬ在山顶西侧大片生长ꎬ集群分布明显ꎬ形成以其为优势种的常绿阔叶灌丛群落ꎬ伴生种有凤凰润楠(Machilusphoenicis)㊁漳州越橘(Vacciniumzhangzhouense)㊁猴头杜鹃(Rhododendronsimiarum)等ꎮ东侧的小片灌丛以原始型为主ꎬ数量较少ꎻ而在变异种群外围亦见零星分布ꎮ北侧狭长㊁地势较低地带的变异型可能由山顶西扩散而来ꎮ在原始种群内部ꎬ发现叶背毛稍显发育的个体ꎬ视其为过渡型ꎮ唯独叶背密生成熟时不脱落的土黄色或黄褐1943期林毅喆等:闽南花岗岩堡状峰乌冈栎种群的表型变异色星状毛ꎬ一年生枝的星状毛也较密集而与原类型不同(图2:F-G)ꎮ红旗岩山顶乌冈栎变异型的花序㊁壳斗㊁坚果(图2:H)及其他叶性状(形状㊁大小㊁质地㊁颜色及叶缘等)均与原类型无异ꎮ2.2叶表皮性状和显著性结果2.2.1叶上表皮(图3:A-E)㊀叶上表皮细胞垂周壁厚度均匀ꎬ有4种形态ꎬ即平直㊁弧形㊁波状㊁蜿蜒(图3:AꎬD-E)ꎬ在各类各地种群间无规律ꎮ除在红旗岩脚原始型2(种群4)未在视野中观察到单细胞毛状体基(STB)以外ꎬ其余或多或少存在单细胞(图3:A)及复合毛状体基(CTB)(图3:BꎬCꎬE)2种类型ꎬ但CTB和STB密度均值小于每平方毫米7个ꎻ在青云山原始型3(种群5)观察到来源于STB的残存的头状毛状体(Ca)ꎬ然而ꎬ均未发现与CTB相对应的毛状体类型ꎮ综上表明ꎬ各类型种群STB与CTB在叶上表皮的密度很小甚至小于每平方毫米1个ꎬ然而即便存在毛状体基ꎬ毛状体也不一定发生或容易脱落ꎬ在宏观上的表现即为无毛ꎮ特征统计见表2ꎮ2.2.2叶下表皮(图3:F-L)㊀叶下表皮细胞垂周壁在各种群内多少表现出平直㊁弧形㊁波状之渐变ꎬ厚度均匀(图3:HꎬJ)ꎮ全部种群均观察到明显的头状毛状体(Ca)及STB(图3:I-L)ꎬ后者的密度为每平方毫米0~10.125个ꎮ由表3可知ꎬ原始型㊁过渡型㊁变异型的STB密度均值递增ꎮ虽然CTB各种群均存在(图3:FꎬH-IꎬK)ꎬ但是由其发生的典型星状毛(Tst)仅见于红旗岩顶变异型(种群1)㊁过渡型(种群2)及青云山原始型3(种群5)(图3:F-GꎬIꎬL)ꎮ然而ꎬ青云山原始型3的叶下表皮星状毛只局限于中脉的基部附近ꎬ对应的CTB密度均值则小于每平方毫米1个ꎻ相比之ꎬ红旗岩顶变异型的Tst存在于中脉㊁任意侧脉或次级脉上ꎬCTB密度均值达每平方毫米57.2个ꎬ中脉㊁侧脉㊁次级脉上的CTB面积逐级减小ꎻ过渡型的CTB密度均值介于两者之间ꎬ为每平方毫米26.076个ꎮ这表明变异型种群与其他种群相比ꎬCTB或Tst的增加很明显ꎬ宏观表现为密被星状毛ꎬ过渡型的叶背虽有星状毛ꎬ但叶下表皮也明显可见ꎬ原始型则几乎完全无毛ꎮ另外ꎬ气孔类型均为无规则型(图3:HꎬJ)ꎮ各种群气孔密度均值在每平方毫米453~516个之间ꎬ气孔尺寸之短径为24.494~25.812μmꎬ长径介于28.906~31.047μmꎬ变异型与原始型在这3个性状的变化无规律ꎮ气孔长宽比均值则在原始型(1.187~1.205)㊁过渡型(1.240)㊁变异型(1.258)间递增ꎮ相关特征统计见表3ꎮ2.2.3显著性检验㊀方差分析结果(表2ꎬ表3)显示ꎬ叶上表皮STB(P=0.0020)㊁叶下表皮STB(P=0.0060)与CTB(P=0.0005)密度及气孔长宽比(P=0.0020)在不同种群之间存在极显著性差异(P<0.01)ꎮ叶上表皮STB密度虽然不同种群差异极显著ꎬ但经事后检验发现变异型与原始型2㊁3ꎬ原始型1与原始型2㊁3共同贡献了该差异ꎬ表明该性状在叶上表皮的发生较随机ꎮ叶下表皮的事后检验结果显示叶下表皮STB与CTB密度及气孔长宽比的差异主要是由变异型引起(图4)ꎮ变异型STB与CTB密度显著大于原始型ꎬ无论原始型位于山顶㊁山脚或异地ꎮ变异型气孔平均长宽比为1.258ꎬ极大值1.333ꎻ原始型均值不大于1.205ꎬ极大值1.274ꎮ这一性状在变异型中在一定程度上有增大ꎬ尽管在影像中看上去并不明显ꎮ过渡型各性状值均居中间水平ꎮ以上结果表明ꎬ变异型在一些叶下表皮性状(STB与CTB密度㊁气孔长宽比)上与原始型相比发生了显著性变异(P<0.01)ꎮ但是ꎬ对生境颇为敏感的气孔密度与尺寸在山顶㊁山脚或异地的种群间未表现出差异性(P>0.05)(表3)ꎮ3㊀讨论与结论3.1堡状峰生境促进了种群分化同为花岗岩地貌的岛山导致种群变异和物种形成的案例已经屡见不鲜(Motaetal.ꎬ2020ꎻRuasetal.ꎬ2020ꎻTavaresetal.ꎬ2022)ꎬ这通常归因于距离隔离导致有限的基因流发生的遗传漂变和近交(Barbaráetal.ꎬ2007ꎻMotaetal.ꎬ2020)ꎬ然而有时异质性生境被认为起到了更大作用ꎬ驱使同一物种在不同种群间发生分化(Miláetal.ꎬ2010ꎻMalletetal.ꎬ2014)ꎮ虽然岛山生境通常不适合物种的生存ꎬ但是世界上一些地区的岛山却成为生物多样性的中心ꎬ如巴西东南部㊁马达加斯加㊁澳大利亚西南部ꎬ推测与隔离及物种长期的坚持促进分化有关(Porembskiꎬ2007)ꎬ因此岛山多特有种ꎮ乌山红旗岩是典型堡状峰地貌ꎬ与岛山类似ꎬ顶部生境与同纬南亚热带季风气候区迥异ꎬ与周边地区相比ꎬ具有一定的特殊性:夏季光照强度大ꎬ日照时间长ꎻ冬季夜间气温低ꎻ气温日较差大ꎻ294广㊀西㊀植㊀物44卷A.乌山红旗岩所在地ꎻB.年轻的堡状峰在乌山普遍存在(三角形示意红旗岩ꎬ箭头示意邻近堡状峰)ꎻC.乌山风景区堡状峰景观ꎮA.LocationofHongqiyaninWushanꎻB.YoungcastlepeaksareprevalentinWushan(triangleshowsHongqiyanꎬarrowsshowneighboringcastlepeaks)ꎻC.CastlepeaklandscapeinWushanScenicArea.图1㊀福建漳州乌山风景区Fig.1㊀WushanScenicAreaꎬZhangzhouꎬFujian降水滞留土壤时间短ꎬ雨水冲刷土壤造成流失ꎻ土壤本身贫瘠而薄ꎻ盛行风速大ꎮ一般情况下ꎬ同一物种在一定程度的干燥与适生条件下生长相比气孔密度增大ꎬ但当干旱进一步加强ꎬ密度将减小ꎬ气孔尺寸则在干旱条件下持续减小(Xu&Zhouꎬ2008)ꎮ然而ꎬ乌山不同生境下的乌冈栎并未表现出这一倾向ꎬ并且在更大空间尺度的栎属植物研究案例中也存在这一现象(Sánchez ̄Acevedoetal.ꎬ2022)ꎮ由于气孔密度与尺寸通常具有一定关联性(Hetherington&Woodwardꎬ2003)ꎬ因此不同种群乌冈栎的气孔尺寸与密度一样无显著性差异ꎮ为了应对乌山山顶特殊生境的挑战ꎬ在我们研究的中乌冈栎似乎倾向于依赖毛状体并调整气孔的形状ꎬ因为结果显示叶下表皮单细胞毛状体基(STB)㊁复合毛状体基(CTB)及气孔长宽比在不同种群间差异显著ꎮ毛状体有吸附空气中水滴㊁减少蒸腾的功能(Molesetal.ꎬ2020)ꎬ在冬季具有一定保暖效果(Pengetal.ꎬ2015)ꎮ就荒漠植物而言ꎬ在应对寒㊁旱㊁盐碱㊁高温㊁强辐射等不同非生物胁迫时表皮毛的变化不一ꎬ在受机械损伤㊁干旱㊁长日照㊁高光照㊁盐胁迫时ꎬ通常会导致表皮毛长度或密度的增大(马亚丽等ꎬ2015)ꎮ乌山顶变3943期林毅喆等:闽南花岗岩堡状峰乌冈栎种群的表型变异A㊁B.生境ꎻC.山顶种群(航拍)ꎻD.变异㊁原始㊁过渡型在山顶的分布ꎻE.灌木状植株ꎻF-H.分别示意变异型(a)与原始型(b)带叶枝条(F)㊁叶片(G)㊁壳斗与坚果(H)ꎮAꎬB.HabitatꎻC.Summitpopulations(aerialphoto)ꎻD.DistributionofvariantꎬoriginalꎬandintermediateformsonthesummitꎻE.ShrubbytreeꎻF-H.Illustratingvariantform(a)andoriginal(b)formofleaf ̄bearingbranches(F)ꎬleaves(G)ꎬcupulesandnuts(H)ꎬrespectively.图2㊀红旗岩顶乌冈栎种群的分布与性状变异Fig.2㊀DistributionandtraitvariationofQuercusphillyreoidespopulationsatthetopofHongqiyan494广㊀西㊀植㊀物44卷A-E.叶上表皮:A-C.变异型(种群1)ꎻD.原始型2(种群4)(示蜿蜒状细胞垂周壁)ꎻE.原始型3(种群5)ꎮF-L.叶下表皮:F-I.变异型(种群1):H的毛状体被人工去除ꎻJ.原始型3(种群5)ꎻK.原始型1(种群3)ꎻL.过渡型(种群2)ꎮ黑色箭头示意复合毛状体基ꎬ白色箭头示意单细胞毛状体基或头状毛状体ꎬ∗指代气孔ꎮ所有缩写如表2所示ꎮA-E.Leafadaxialepidermis:A-C.Variantform(Population1)ꎻD.Originalform2(Population4)(showingsinuousanticlinalwallofthecells)ꎻE.Originalform3(Population5).F-L.Leafabaxialepidermis:F-I.Variantform(Population1):trichomesofHareartificiallyremovedꎻJ.Originalform3(Population5)ꎻK.Originalform1(Population3)ꎻL.Intermediateform(Population2).BlackarrowsshowingtheCTBꎬwhitearrowsshowingtheSTBorCaꎬ∗indicatesstomata.AlltheabbreviationsareshowninTable2.图3㊀乌冈栎变异㊁原始㊁过渡型的叶表皮形态Fig.3㊀LeafepidermalmorphologyofvariantꎬoriginalꎬandintermediateformsofQuercusphillyreoides异型乌冈栎的叶背STB㊁CTB密度与各原始型相比均显著ꎬ尽管毛状体在极端环境中可能被催生ꎬ但山顶原始型1的存在ꎬ以及与原始型2㊁3之间无显著差异ꎬ证实环境的短期作用对塑造乌冈栎叶背毛状体的能力有限ꎮ显著增多叶背的星状毛(Tst)势必有助其适应山顶贫瘠土壤储水少㊁高蒸5943期林毅喆等:闽南花岗岩堡状峰乌冈栎种群的表型变异方框中的粗水平条表示中位数ꎬ细水平条表示上㊁下四分位数ꎬ方框外的水平条表示极值ꎮ字母a㊁b㊁ab等表示类别间的差异显著性ꎮThethickhorizontalbarintheboxindicatesthemedianꎬthethinhorizontalbarindicatestheupperandlowerquartilesꎬandthehorizontalbaroutsidetheboxindicatestheextremevalues.Thelettersaꎬbꎬabꎬetc.indicatesignificantdifferencesbetweencategories.图4㊀乌冈栎各类型叶下表皮复合毛状体基、单细胞毛状体基密度及气孔长宽比的箱形图Fig.4㊀Box ̄plotofthedensitiesofleafabaxialsurfaceCTBꎬSTBꎬandstomatalength ̄widthratioofeachformofQuercusphillyreoides散导致的干旱ꎬ以及冬季花岗岩山体比热小㊁散热快导致的冷害㊁冻害ꎮ头状毛状体具备合成㊁贮藏和分泌诸多代谢物的作用(张继伟等ꎬ2018)ꎬ增多的头状毛或能增加乌冈栎应对山顶环境胁迫的抗性ꎮ关于环境对毛状体的作用机制ꎬ就拟南芥(Arabidopsisthaliana)的研究显示ꎬ拥有丰富表皮毛的突变体的较于表皮毛缺失突变体其茎㊁叶蜡质层含有的C32+脂肪酸混合物含量显著增加ꎬ并且表皮毛所覆盖的蜡质C32+脂肪酸含量亦被证实高于周围细胞ꎬ表皮毛可联合蜡质层生物合成来利于减少增发和调控温度(Hegebarthetal.ꎬ2016)ꎮ对黄瓜(Cucumissativus)表皮毛突变体的研究表明ꎬGL1与TTG1为叶表皮发生的核心基因ꎬGL3㊁EGL3为必要基因ꎬ这些基因的转录产物构成正调控因子(Liuetal.ꎬ2016)ꎮ但是ꎬ在TRY㊁CPC等基因转录的负调控因子介导下ꎬ表皮毛的发生受到抑制(Schellmannetal.ꎬ2002)ꎮ毛状体的发生发育机理研究表明其由多基因调控(张继伟等ꎬ2018)ꎬ变异型乌冈栎叶背毛状体作为山顶环境胁迫响应的产物ꎬ可能经历了负调控因子突变㊁正调控因子转座或增强等过程ꎮ堡状峰顶环境因子与毛状体发生的内在联系需进一步研究方可明确ꎮ气孔长宽比实际上反映了气孔开度(stomatalaperture)ꎬ其变化依赖于受离子及糖控制膨压的副卫细胞㊁细胞骨架组织㊁跨膜运输及基因表达的联合作用ꎬ大量证据显示单一的刺激性信号通路不足以让气孔开度发生变化(Hetherington&Woodwardꎬ2003)ꎮ在本次研究的乌冈栎种群间ꎬ与毛状体同理ꎬ同一时刻原始型1的气孔开度似乎没有因居处堡状峰顶而与山脚或青云山的原始型2㊁3在这一性状上表现出显著差异ꎮ但是ꎬ在变异型中ꎬ气孔长宽比显著增大ꎬ即气孔形状发生了变异ꎬ干旱㊁叶温过高或过低经信号传递最终在叶片上表现出气孔开度的减小(姜籽竹等ꎬ2015ꎻ段骅等ꎬ2019)ꎮ由此直观地推测ꎬ在同等条件下变异型种群通过减少蒸腾以便应对干旱或极端气温ꎬ并在大风环境中关闭气孔使机动性得以增强ꎮ综上所述ꎬ堡状峰地貌是变异产生的渊缘ꎬ该地貌生境ꎬ特别是贫瘠而薄的土壤ꎬ导致水热条件差于周围地区ꎬSTB㊁CTB及气孔长宽比的变异对适应该生境有积极作用ꎮ这些性状变异应是稳定的ꎬ它们并不受环境胁迫快速而直接地形成ꎻ经过1年的观察ꎬ在变异型或原始型内ꎬ不同发育时期亦稳定不变ꎮ3.2种群分化的可能途径乌山直接形成于新近纪后中国东南海岸地带不同程度的隆起抬升与风化侵蚀(Chenetal.ꎬ694广㊀西㊀植㊀物44卷表2㊀调查种群叶上表皮性状统计Table2㊀Statisticsofleafadaxialsurfacecharacteristicsofinvestigatedpopulations编号Number种群类型Populationform叶上表皮Leafadaxialsurface垂周壁Anticlinalwall毛状体类型Trichometype毛状体基类型ꎬ密度Trichomebasetypeꎬdensity(No. mm ̄2)1变异型Variantformstr ̄cur ̄un/uniSTB/CTBꎬ6.239ʃ2.866a/4.556ʃ3.049a2过渡型Intermediateformstr ̄cur/uniSTB/CTBꎬ3.365ʃ2.896abc/5.608ʃ3.547a3原始型1Originalform1str ̄cur/uniSTB/CTBꎬ4.697ʃ2.282ab/1.823ʃ1.579a4原始型2Originalform2cur ̄un ̄sin/uniSTB/CTBꎬ-c/3.739ʃ1.713a5原始型3Originalform3str ̄cur ̄un/uniCaSTB/CTBꎬ1.963ʃ2.055bc/5.818ʃ3.495a㊀注:垂周壁表示叶表皮细胞垂周壁ꎮstr.平直ꎻcur.弧形ꎻun.波状ꎻsin.蜿蜒状ꎻuni.均一厚度ꎻCa.头状毛状体ꎻTst.典型星状毛状体ꎻSTB.单细胞毛状体基ꎻCTB.复合毛状体基ꎻ-表示密度小于1No. mm ̄2ꎮ字母a㊁b㊁ab等表示类别间的差异显著性ꎮ下同ꎮ㊀Note:Anticlinalwallindicatesanticlinalwallofleafepidermalcells.str.Straightꎻcur.Curvedꎻun.Undulateꎻsin.Sinuousꎻuni.UniformthicknessꎻCa.CapitateꎻTst.TypicalstellatetrichomeꎻSTB.Simple ̄celledtrichomebaseꎻCTB.Compoundtrichomebaseꎻ-indicatesdensityislessthan1No. mm ̄2.Thelettersaꎬbꎬabꎬetc.indicatesignificantdifferencesbetweencategories.Thesamebelow.表3㊀调查种群叶下表皮性状统计Table3㊀Statisticsofleafabaxialsurfacecharacteristicsofinvestigatedpopulations编号Number种群类型Populationform叶下表皮Leafabaxialsurface垂周壁Anticlinalwall毛状体类型Trichometype毛状体基类型ꎬ密度Trichomebasetypeꎬdensity(No. mm ̄2)气孔类型ꎬ密度Stomatatypeꎬdensity(No. mm ̄2)气孔尺寸Stomatasize(μm2)气孔长宽比Stomatalength ̄widthratio1变异型Variantformstr ̄cur ̄un/uniCa/TstSTB/CTBꎬ10.125ʃ4.101a/57.200ʃ21.847aAnomocyticꎬ492.299ʃ36.764a25.032ʃ1.452aˑ30.765ʃ2.135a1.258ʃ0.042a2过渡型Intermediateformstr ̄cur/uniCa/TstSTB/CTBꎬ7.430ʃ2.799ab/26.076ʃ7.042bAnomocyticꎬ487.651ʃ45.376a24.596ʃ1.357aˑ29.409ʃ1.581a1.240ʃ0.029a3原始型1Originalform1str ̄cur/uniCaSTB/CTBꎬ3.785ʃ2.743bc/1.122ʃ1.799cAnomocyticꎬ457.671ʃ56.611a25.812ʃ1.819aˑ31.047ʃ1.790a1.205ʃ0.027b4原始型2Originalform2cur ̄un/uniCaSTB/CTBꎬ-c/0.748ʃ0.648cAnomocyticꎬ515.081ʃ91.284a24.494ʃ0.928aˑ28.906ʃ0.877a1.187ʃ0.011b5原始型3Originalform3cur ̄un/uniCa/TstSTB/CTBꎬ4.233ʃ3.324bc/0.481ʃ0.600cAnomocyticꎬ453.885ʃ48.229a25.539ʃ0.861aˑ30.320ʃ1.118a1.194ʃ0.027b㊀注:Anomocytic.无规则型气孔ꎮ㊀Note:Anomocytic.Anomocytictypeofstomata.2009)ꎬ更新世末次冰期导致高海拔植物下潜到附近 避难所 (陈冬梅等ꎬ2011)ꎬ在间冰期或全新世(Holocene)很可能让乌冈栎重回山顶ꎮ啮齿类是壳斗科种子传播的主要贡献者ꎬ但为抵达堡状7943期林毅喆等:闽南花岗岩堡状峰乌冈栎种群的表型变异峰顶ꎬ最初的迁移可能依赖于鸟类(Ohꎬ2015)ꎮ原始型乌冈栎抵达山顶后ꎬ若受生境胁迫发生适应性演化ꎬ适应性强的个体具有优势ꎬ进而扩大成种群ꎮ然而ꎬ山顶却存在小范围分布或零散于变异型周围的原始型种群或个体ꎮ这表明原始型本身尚可忍耐干旱等极端生境ꎮ可以将其看作是基于自然选择与竞争排除原理种群残存的一个剖面ꎬ同时ꎬ鸟类传播这类偶然事件也可能对山顶群落演替产生重大影响ꎬ这均可导致当前原始型与变异型共存ꎬ并且通过杂交让过渡型产生ꎮ这表明过渡型之所以被观察到ꎬ可能是物种形成过程残留的随机变异个体ꎬ抑或是杂交的产物ꎮ但是作为风媒树种ꎬ四周低地的原始型花粉流应当从未断绝(Ashleyꎬ2021)ꎮ红旗岩堡状峰的地理隔离作用可能相当有限ꎬ堡状峰特殊生境显然较地理隔离对变异的产生起到了更大的促进作用ꎮ山顶的过渡型及山脚未调查到变异型的现状表明变异型可以与原始型融合而消失ꎮ但是就目前而言ꎬ堡状峰的确产生了一定的生态学岛屿效应ꎮ3.3祖先性状和平行演化较于气孔长宽比ꎬ毛状体的变异在栎属内有更多演化规律ꎮ首先需明确ꎬ乌山的变异型乌冈栎与拥有类似叶背毛状体的近缘种直线距离超过300kmꎬ可排除杂交起源ꎮDeng等(2017)研究表明ꎬSTB是栎属的祖征ꎬ不同演化支或多或少存在ꎬ可在种间存在显著的密度差异ꎬ乌山变异型与原始型乌冈栎叶上下表皮STB即如此ꎮ然而ꎬ栎属分子系统结果显示CTB在不同支系中多次起源或丢失(Yangetal.ꎬ2017ꎻHippetal.ꎬ2020)ꎬ冬青砾组不同种的情况不一ꎬ因此变异型发生CTB密度的变异是可能的ꎮ表皮毛发育受多基因直接与间接调控(普莉等ꎬ2003)ꎬ选择促进基因互作与重排ꎬ古老祖先多态性分选也可使类似性状反复产生ꎮ为解析它们的关系ꎬ需利用分子手段做各类型群体的研究ꎮ另外ꎬ乌冈栎的毛状体变异在广西的标本(陈照宙50295)ꎬ以及GlobalBiodiversityInformationFacility(GBIFꎬhttps://www.gbif.org/)额外检查的56份标本中的2份来自琉球群岛的标本(Wrightꎬ无号ꎻTypeofQuercuswrightii=Q.phillyreoides)中发现ꎬ若这不是单株突变ꎬ那就表明它们之间可能发生了平行演化ꎮ综上得到以下结论:福建漳州乌山红旗岩的乌冈栎变异型的叶下表皮STB㊁CTB㊁气孔长宽比与原始型相比发生了自然变异ꎬ即STB㊁CTB显著增多(P<0.01)ꎬ气孔长宽比显著增大(P<0.01)ꎮ乌山堡状峰特殊生境促使变异产生ꎬ其具备一定生态与演化合理性ꎮ这为亚热带特殊花岗岩地貌促进种群分化提供了有效证据ꎮ乌冈栎广泛分布于中国南方及日本ꎬ是毛状体发生与演化㊁平行物种形成等研究的理想对象ꎮ中国是世界上花岗岩分布最广的国家之一ꎬ又以广东和福建两省最多(占中国30%~40%)(崔之久等ꎬ2007)ꎮ构造运动加速花岗岩出露地表ꎬ形成极其多样的地貌景观ꎬ中㊁高度隆起形成的堡状峰㊁峰丛㊁石林㊁独立巨峰等在大小空间尺度上广泛存在ꎮ特殊花岗岩地貌上的植物区系㊁群落生态㊁极小种群㊁物种演化等有望被进一步关注ꎮ参考文献:ARENAETꎬRUEDENCTꎬHINERMCꎬetal.ꎬ2017.Quantitatingthecell:turningimagesintonumberswithImageJ[J].WileyInterdiscipRevDevBiolꎬ6(2):e260.ASHLEYMVꎬ2021.Answersblowinginthewind:Aquartercenturyofgeneticstudiesofpollinationinoaks[J].Forestsꎬ12(5):575.BARBARÁTꎬLEXERCꎬMARTINELLIGꎬetal.ꎬ2008.Within ̄populationspatialgeneticstructureinfournaturallyfragmentedspeciesofaneotropicalinselbergradiationꎬAlcantareaimperialisꎬA ̄geniculataꎬA ̄glaziouanaandA ̄regina(Bromeliaceae)[J].Heredity(Edinb)ꎬ101(3):285-296.BARBARÁTꎬMARTINELLIGꎬFAYMFꎬetal.ꎬ2007.Populationdifferentiationandspeciescohesionintwocloselyrelatedplantsadaptedtoneotropicalhigh ̄altitude inselbergs ꎬAlcantareaimperialisandAlcantareageniculata(Bromeliaceae)[J].MolEcolꎬ16(10):1981-1992.BYRNEMꎬKRAUSSSLꎬMILLARMAꎬetal.ꎬ2019.Persistenceandstochasticityarekeydeterminantsofgeneticdiversityinplantsassociatedwithbandedironformationinselbergs[J].BiolRevCambPhilosSocꎬ94(3):753-772.CHENYXꎬCUIZJꎬYANGJQꎬ2009.InfluenceofclimateandtectonicmovementsongranitelandformsinChina[J].JGeogrSciꎬ19(5):587-599.CHENDMꎬKANGHZꎬLIUCJꎬ2011.AnoverviewonthepotentialQuaternaryglacialrefugiaofplantsinChina894广㊀西㊀植㊀物44卷。