第七章-线粒体
- 格式:ppt
- 大小:2.92 MB
- 文档页数:33

第七章线粒体西北农林科技大学生命科学学院李绍军17334040@本章概要:7.1线粒体的形态与结构7.2线粒体的化学组成7.3线粒体的功能7.4线粒体质量控制与线粒体自噬7.5线粒体损伤与疾病7.6线粒体与细胞凋亡7.7线粒体的自主性问题7.8线粒体的分裂与增殖7.9线粒体的起源学习重点:1、线粒体的结构与功能。
2、线粒体与疾病、细胞凋亡之间的关系。
线粒体和叶绿体是能量转换的细胞器,细胞内各项生化活动所需的能量,主要由线粒体和叶绿体提供,故有细胞“动力厂”之称。
线粒体是在动物细胞中首先由本达(Benda,1897)发现的,命名为“mitochondria,mitochondrion”(来源于希腊字mitos=线,chondrion=粒),其后,在植物细胞(Meves,1904)中也看到了。
它们普遍地存在于真核生物的所有细胞中。
7.1线粒体的形态与结构7.1.1 形状、大小、数目和分布线粒体是细胞中最丰富的细胞器之一,通常占细胞质容积的20%~25%,其数目、形态受细胞对能量的需求的调节,因而是动态变化的。
线粒体各种结构类型模式图线粒体形状在一定的条件下是可以可逆的转变的,其转变方式有3种:①由线状断裂成小球或颗粒,②颗粒膨大呈中空的球状,③由球状、粒状或短棒状转变为线状。
6The Fusion and Fission of Mitochondria7.1.2 结构线粒体的基本结构可分为4 部分,①线粒体表面的外膜(outer membrane),内含脂肪和蛋白质,各占一半;②内膜(inner membrane),向内折叠伸出许多形式不同的嵴,形成复杂的内部膜系统,嵴内为嵴内腔;③内外膜间为8.5nm 厚的电子透明层,称为膜间隙(intermembrane space)它与嵴内腔相贯通;④在内膜以内的基质(matrix),为含有可溶性蛋白质和含钙的基质颗粒(matirx granule)等物质的溶液。



细胞生物学第七章线粒体与叶绿体知识点整理线粒体和叶绿体是细胞中两个重要的细胞器。
它们在细胞代谢和能量转换中发挥着重要的作用。
以下是关于线粒体和叶绿体的一些重要知识点:线粒体:1.结构:线粒体是一个由两层膜包围的细胞器。
它包含一个外膜和一个内膜,内膜形成了许多内突起,称为线粒体内膜嵴。
2.能量转换:线粒体是细胞中的能量生产中心。
它通过细胞呼吸过程中的氧化磷酸化来产生能量,将食物分子中的化学能转化为细胞可以使用的三磷酸腺苷(ATP)。
3. 基因组:线粒体具有自己的基因组,称为线粒体DNA(mtDNA)。
它主要编码细胞呼吸过程中所需的蛋白质。
mtDNA由母亲遗传给子代,因此线粒体DNA有助于研究人类的遗传和进化。
4.线粒体疾病:线粒体功能障碍可以导致许多疾病,如线粒体脑肌病、线粒体糖尿病和阿尔茨海默病。
这些疾病通常会影响能量的产生和细胞的正常功能。
叶绿体:1.结构:叶绿体是植物和一些原生生物中的细胞器。
它也是由两层膜包围,并且内膜形成了一系列叫做叶绿体嵴的结构。
2.光合作用:叶绿体是光合作用的主要场所,其中光能转化为化学能以供细胞使用。
叶绿体中的叶绿素能够吸收太阳能,并将其转化为光合作用的产物,如葡萄糖。
3. 基因组:叶绿体也具有自己的基因组,称为叶绿体DNA(cpDNA)。
它主要编码参与光合作用和叶绿体功能的蛋白质。
4.叶绿体疾病:类似于线粒体疾病,叶绿体功能障碍也会导致一系列疾病,在植物中称为叶绿体遗传病。
这些疾病通常会导致叶绿体的正常结构和功能受损。
1.起源:线粒体起源于古代原核生物,而叶绿体起源于古代蓝藻细菌。
这些细菌进化成为现代细胞中的线粒体和叶绿体。
2.功能:线粒体主要参与能量转换,而叶绿体主要参与光合作用。
它们在细胞代谢中的角色不同,但都与能量生产和细胞功能密切相关。
3.基因组:线粒体和叶绿体都有自己的基因组,具有其中一种程度的自主复制和表达能力。
不过,线粒体基因组比较小,叶绿体基因组比较大。


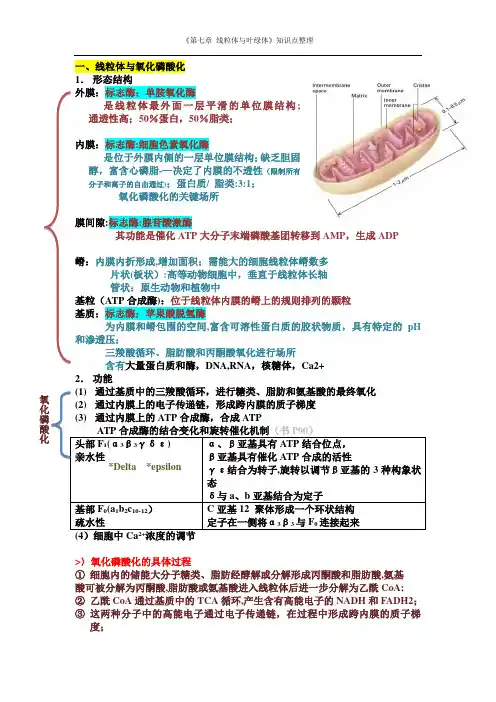
《第七章 线粒体与叶绿体》知识点整理一、线粒体与氧化磷酸化 1. 形态结构 外膜:标志酶:单胺氧化酶 是线粒体最外面一层平滑的单位膜结构; 通透性高;50%蛋白,50%脂类; 内膜:标志酶:细胞色素氧化酶 是位于外膜内侧的一层单位膜结构;缺乏胆固醇,富含心磷脂-—决定了内膜的不透性(限制所有分子和离子的自由通过);蛋白质/ 脂类:3:1; 氧化磷酸化的关键场所 膜间隙:标志酶:腺苷酸激酶 其功能是催化ATP 大分子末端磷酸基团转移到AMP ,生成ADP 嵴:内膜内折形成,增加面积;需能大的细胞线粒体嵴数多 片状(板状):高等动物细胞中,垂直于线粒体长轴 管状:原生动物和植物中 基粒(ATP 合成酶):位于线粒体内膜的嵴上的规则排列的颗粒 基质:标志酶:苹果酸脱氢酶 为内膜和嵴包围的空间,富含可溶性蛋白质的胶状物质,具有特定的pH 和渗透压; 三羧酸循环、脂肪酸和丙酮酸氧化进行场所 含有大量蛋白质和酶,DNA,RNA ,核糖体,Ca2+ 2. 功能 (1) 通过基质中的三羧酸循环,进行糖类、脂肪和氨基酸的最终氧化 (2) 通过内膜上的电子传递链,形成跨内膜的质子梯度 (3) 通过内膜上的ATP 合成酶,合成ATP ATP 合成酶的结合变化和旋转催化机制(书P90)头部F 1(α3β3γδε) 亲水性 α、β亚基具有ATP 结合位点,β亚基具有催化ATP 合成的活性 γε结合为转子,旋转以调节β亚基的3种构象状态δ与a 、b 亚基结合为定子基部F 0(a 1b 2c 10-12) 疏水性 C 亚基12 聚体形成一个环状结构定子在一侧将α3β3与F 0连接起来>〉氧化磷酸化的具体过程① 细胞内的储能大分子糖类、脂肪经酵解或分解形成丙酮酸和脂肪酸,氨基 酸可被分解为丙酮酸,脂肪酸或氨基酸进入线粒体后进一步分解为乙酰CoA;② 乙酰CoA 通过基质中的TCA 循环,产生含有高能电子的NADH 和FADH2; ③ 这两种分子中的高能电子通过电子传递链,在过程中形成跨内膜的质子梯度; 氧化磷酸化*Delta *epsilon《第七章 线粒体与叶绿体》知识点整理④ 质子梯度驱动ATP 合成酶将ADP 磷酸化成ATP,势能转变为化学能。

第七章线粒体疾病的遗传几乎每个人体细胞中都含有数以百计的线粒体,每个线粒体内膜上富含5种具有传递电子功能的酶复合体,组成呼吸链-氧化磷酸化系统,将代谢过程中产生的电子通过连锁反应逐步传递给氧并将质子从线粒体基质转移到膜间腔,形成跨内膜的质子梯度,复合物Ⅴ(ATP合酶)利用质子顺浓度回流到基质中所产生的势能,使ADP磷酸化生成ATP,为细胞提供进行各种生命活动所需要的能量。
线粒体内还含有DNA 分子,被称为人类第25号染色体,是细胞核以外含有遗传信息和表达系统的细胞器,其遗传特点表现为非孟德尔遗传方式,又称核外遗传。
1981年Anderson 等人完成了人类线粒体基因组的全部核苷酸序列的测定,1988年,Wallace等发现Leber视神经病的发生与线粒体DNA(mitochondrial DNA,mtDNA)突变有关。
近年来,人们发现线粒体不仅在细胞的生长代谢中起重要作用,而且mtDNA突变是许多人类疾病的重要病因。
第一节人类线粒体基因组 mtDNA编码线粒体中部分蛋白质和全部的tRNA、rRNA,能够独立进行复制、转录和翻译,但所含信息量小,呼吸链-氧化磷酸化系统的80多种蛋白质亚基中,mtDNA仅编码13种,绝大部分蛋白质亚基和其他维持线粒体结构和功能的蛋白质都依赖于核DNA (nuclear DNA,nDNA)编码,在细胞质中合成后,经特定转运方式进入线粒体。
此外,mtDNA基因的表达受nDNA的制约,线粒体氧化磷酸酶化系统的组装和维护需要nDNA和mtDNA的协调,二者共同作用参与机体代谢调节,因此线粒体是一种半自主细胞器,受线粒体基因组和核基因组两套遗传系统共同控制(图7-1),nDNA与mtDNA基因突变均可导致线粒体中蛋白质合成受阻,细胞能量代谢缺陷。
图7-1 mtDNA与nDNA的协同作用一、线粒体基因组线粒体基因组是人类基因组的重要组成部分,全长16569bp,不与组蛋白结合,呈裸露闭环双链状,根据其转录产物在CsCl中密度的不同分为重链和轻链,重链(H链)富含鸟嘌呤,轻链(L链)富含胞嘧啶。
第七章线粒体与过氧化物酶体7.1线粒体7.1.1线粒体的结构、组成与特性(1)形态与分布棒杆状结构,可以有分支,而且有时多个线粒体可以形成网络结构。
两层膜,内膜内褶形成层状或管状,增大了膜面积。
线粒体在不同类型细胞内的数目不同,依其需能多少而定。
其具有自由移动的特性,能够移动到细胞需能的部位或附近。
(2)组成与特性两膜两空间。
线粒体的蛋白质的比重比较大,含有丰富的心磷脂和较少的胆固醇。
①外膜外膜脂类与蛋白的比例比内膜高,含有孔蛋白,具有较高的通透性。
同时外膜上有参与被氧化物质初步分解的酶。
标志酶为单胺氧化酶。
②内膜内膜脂类与蛋白的比例小,含有大量的心磷脂,其蛋白可以分为三类:转运蛋白;电子传递链复合体和ATP合酶;合成酶类。
其通透性非常低。
标志酶为细胞色素氧化酶③膜间间隙其成分由于外膜的通透性差的缘故,跟细胞胞质溶胶的成分相近。
标志酶为腺苷酸激酶。
④线粒体基质含有参与三羧酸循环、脂肪酸氧化、氨基酸降解的酶系,以及DNA,rRNA,tRNA 基因转录的酶系及核糖体。
标志酶为苹果酸脱氢酶。
7.1.2线粒体蛋白的定位7.1.2.1蛋白质定位的机制游离核糖体合成的蛋白质及膜结合核糖体合成的蛋白质的定位方式不同,定位到不同的细胞器。
(1)信号序列一般位于蛋白质N端的一段序列,蛋白质的标签,包括信号定位信号,死亡信号,修饰信号三种信号。
有时在其下游还有一段信号序列,或者有停止转运序列。
(2)蛋白质转运方式①翻译后转运蛋白质寻靶:游离核糖体上的蛋白质完全合成后再在转运肽的导向下定位。
转运肽的介导的蛋白质定位步骤为,在分子伴侣的协助下,蛋白质解折叠,伸出转运肽;转运肽与膜上受体作用,从内外膜形成接触点上穿过运输蛋白,这一过程需要ATP水解及膜电位的协助;进入基质的蛋白质首先在分子伴侣的协助下重新折叠,同时转运肽酶水解掉转运肽。
②共翻译转运蛋白质分选:膜结合核糖体上合成的蛋白质一边翻译,一边通过信号序列进入内质网。
第七章细胞的能量转换――线粒体和叶绿体生物的基本能量来源于太阳光的辐射能。
但生物体不能直接利用太阳光的辐射能,必须先使之转换成化学能,再为生物体利用。
叶绿体通过光合作用把光能转换为化学能,并储存于糖类、脂肪和蛋白质等大分子有机物中。
线粒体是一种高效地将有机物转换为细胞生命活动的直接能源ATP的细胞器。
因此,线粒体和叶绿体是细胞内的两种产能细胞器。
线粒体和叶绿体都具有环状DNA及自身转录RNA与翻译蛋白质体系。
很多学者把线粒体和叶绿体的遗传信息系统称为真核细胞的第二遗传信息系统,或称为核外基因及其表达体系。
线粒体和叶绿体都是半自主性细胞器。
第一节线粒体与氧化磷酸化人体内的细胞每天要合成几千克的ATP,且95%的ATP是由线粒体中的呼吸链所产生,因此线粒体被称为细胞内的“能量工厂”(power plants)。
线粒体通过氧化磷酸化作用,进行能量转换,为所需要的细胞进行各种生命活动提供能量。
一、线粒体的形态结构1、线粒体的形态、大小、数量与分布1)线粒体的形状线粒体的形状各种各样,以线状和颗粒状最常见。
也可呈环形、哑铃形、枝状或其他形状。
2)线粒体的大小线粒体的一般直径为0.5-1.0um,长,1.5-3.0um。
有的长达5um(如肝细胞)或10-20 um(胰腺细胞)或40 um(人的成纤维细胞)。
3)线粒体的数量线粒体的数目由数百――数千个不等。
如利什曼原虫中只有一个巨大的线粒体,海胆卵细胞则多达30万个。
4)线粒体的分布线粒体在细胞中的分布一般是不均匀的。
二、线粒体的超微结构在电镜下观察到线粒体是由两层单位膜套叠而成的封闭的囊状结构。
主要由外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)、基质(matrix)或内室(inner chamber)4部分组成。
图7-11.外膜外膜是包围在线粒体最外面的一层单位膜,光滑而有弹性,厚约6 um。