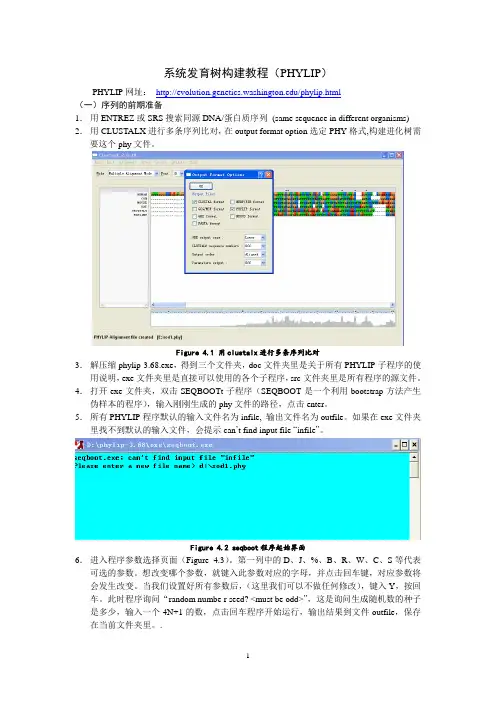
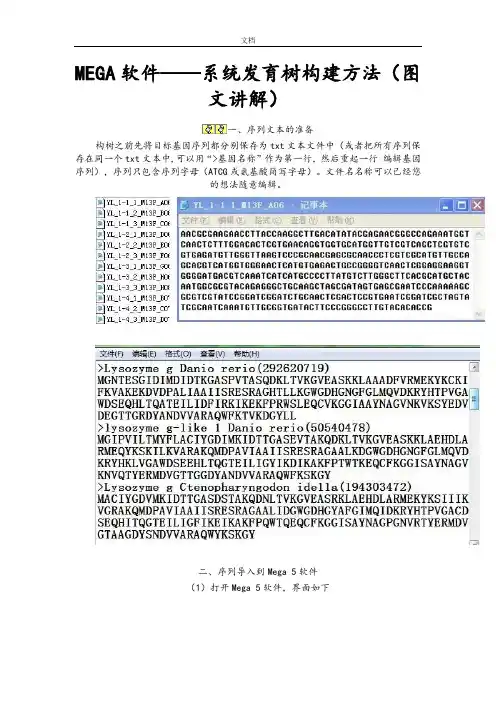
系统发育树的构建练习
- 格式:pdf
- 大小:539.54 KB
- 文档页数:4
通过16s rDNA 研究认识青鳉属内部物种的系统发育关系前言青鳉分布在东亚及东南亚,包括中国大陆、日本、韩国、越南、缅甸、泰国、老挝、柬埔寨、台湾的淡水水域。
本鱼每日产卵、胚胎透明、世代时间短、基因组小并已建立转基因技术[1]。
这些作为实验动物的优势在近缘物种中都有体现,为比较方法提供了理想的条件。
虽然青鳉已经引起生物学家的研究兴趣,并有青鳉的渗透调节[2]、免疫[3]和性选择[4]等不同领域研究报道,但是不同青鳉物种间系统发育关系仍有很多未解之谜[5]。
由于不同种的真细菌与古细菌间的16S rRNA基因(16S rDNA)是高度保守的[6],16S rDNA常被用于对各种生物进行的系统发生学方面的研究[7]这种运用16S rRNA 对生物进行系统发生学研究的方法由卡尔·沃斯(Carl Woese)开创[8]。
故通过16s rDNA研究认识辐鳍亚纲(Actinopterygii)颌针鱼目(Beloniformes)异鳉科(Adrianichthyidae)青鳉属(Oryzias)内部11个物种的系统发育关系。
方法从NCBI(https:///)下载已经报道的青鳉属和花鳉科相关类群的16S rRNA基因序列,所选用的物种信息见表1。
表1 本研究所选青鳉属和花鳉科物种序列和地理分布Table 1 Sequences and geographic distributions of the Oryzias and Poeciliidae species used in this study 物种序列编号序列长度/bp 采集点Species Sequence number length/bp Collection site深青鳉Oryzias profundicola AB188740.1 470 Indonesia:Sulawesi, Timampu, Lake Towuti 湄公河青鳉Oryzias mekongensis AB188736.1 475 Thailand:Nakhon Phanom印尼青鳉Oryzias matanensis AB188734.1 470 Indonesia:Sulawesi, Soroako, Lake Matano 吕宋青鳉Oryzias luzonensis AB188732.1 471 Philippines:Ilocos Norte, Solsona青鳉Oryzias latipes AB188731.1 471 China:Kunming爪哇青鳉Oryzias javanicus AB188725.1 470 Indonesia恒河青鳉Oryzias dancena AB188722.1 471 Thailand:Phuket弓背青鳉Oryzias curvinotus AB188720.1 471 Viet Nam:Caudien near Hanoi西里伯斯青鳉Oryzias celebensis AB188718.1 470 Indonesia:Sulawesi, Ujung Pandang苏拉威西青鳉Oryzias sarasinorum AB188742.1 470 Indonesia:Sulawesi, Lake Lindu花斑青鳉Oryzias marmoratus AB188733.1 470 Indonesia:Sulawesi, Timampu, Lake Towuti 剑尾鱼Xiphophorus helleri U80047.1 448孔雀鱼Poecilia reticulata U80051.1 4481.实验对象选择选择辐鳍亚纲,鳉形目,青鳉属的11个物种作为进行谱系进化比较的目标物种。
2.外群选择同时选取同样为辐鳍亚纲的鳉形目(Cyprinodontiformes)花鳉科(Poeciliidae)作为2个外源物种进行比较。
3.分析对象选择综合提取DNA的难度和稳定性,选择16S rDNA作为分析对象。
4.样品采集由于目标生物种类较少,故每个种选取5个个体进行采集,样本主要是背部肌肉。
均采自野外。
5.分子实验选取相应的试剂盒进行16S rDNA的提取,设计引物进行PCR扩增,之后将PCR 原液送往测序公司进行测序。
本研究采用双向测序,最终序列由正反向两条序列拼接而成。
6.数据分析首先对原始数据进行序列质量、序列长度和序列分布情况的统计。
进行数据预处理后,对高质量序列进行有效序列统计并做物种分类。
7.系统进化分析贝叶斯法建树(BEAST):使用贝叶斯法(Bayesian method)构建系统进化树。
由于贝叶斯法对替换模型很敏感,所以先使用软件jModelTest并根据分析结果选择HKY作为替换模型。
使用软件BEAUti和BEAST进行系统发育分析,Site Heterogeneity Model选择None,即假定物种的进化保持相同的速率。
使用软件TreeAnnotator寻找最大可信树(Maximum clade credibility tree),老化(burn-in)数据设定为1000,即运行的马尔可夫链最初的1000世代。
结果本研究所构建的系统发育树见图1。
图1 使用16s rRNA基因对11种青鳉属和2种花鳉科物种构建的系统发育树Figure 1 Phylogenetic tree of 11 Oryzias and 2 Poeciliidae species by 16s rDNA根据采集序列样本的地理位置以构建的系统发育树为模型建立样本地图见图2.图2 展示图1系统发育树分支的样本地图:花斑青鳉(蓝),深青鳉(蓝),印尼青鳉(蓝),苏拉威西青鳉(蓝),西里伯斯青鳉(蓝),恒河青鳉(粉),爪哇青鳉(粉),弓背青鳉(黄),吕宋青鳉(黄),青鳉(黄),湄公河青鳉(黄)。
Figure 2. Sample map showing lineage of phylogenetic tree in figure 1: Oryzias marmoratus(blue), Oryzias profundicola(blue), Oryzias matanensis(blue), Oryzias sarasinorum(blue), Oryziascelebensis(blue), Oryzias dancena(pink),Oryzias javanicus(pink), Oryzias curvinotus(yellow), Oryzias luzonensis(yellow), Oryzias latipes(yellow), Oryzias mekongensis(yellow).讨论通过匹配最优的核苷酸替代模型,选择剑尾鱼和孔雀鱼为外群确定进化树的根,最终获得进化树上青鳉属的各个单系群。
本研究样本主要从东亚及东南亚地区采集,通过构建图2样本地图进一步验证了图1系统发育树的结果可靠性。
并且可以获知东亚大陆地区和东南亚群岛上的青鳉物种基本上可以认为是起源于同一祖先的两个姐妹群;隔海相望的越南地区的弓背青鳉和菲律宾的吕宋青鳉亲缘关系较近;印度尼西亚中部苏拉威西岛上的五种青鳉的亲缘关系亲疏与地理分布距离密切相关。
参考文献[1] Medakafish as a model system for vertebrate developmental genetics. BioEssays 22, 487–495. Ishikawa, Y., 2000、[2] Asian medaka fishes offer new models for studying mechanisms of seawater adaptation.Inoue, K., Takei, Y., 2003. Biochem.Physiol. B 136, 635–645.[3] The development of cellular immune defence in marine medaka Oryzias melastigma. Seemann, F ; Peterson, DR ; Chiang, MWL; Au, DWT. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology V olume 199, September 2017, Pages 81-89[4] Co-option of Sox3 as the male-determining factor on the Y chromosome in the fish Oyrzias dancena. Yusuke Takehana, Masaru Matsuda, Taijun Myosho, Maximiliano L. Suster, Koichi Kawakami, Tadasu Shin-I, Yuji Kohara, Yoko Kuroki, Atsushi Toyoda, Asao Fujiyama, Satoshi Hamaguchi, Mitsuru Sakaizumi & Kiyoshi Naruse. Nature Communications 5, Article number: 4157 (2014)、[5]Molecular phylogeny of the medaka fishes genus Oryzias (Beloniformes: Adrianichthyidae) based on nuclear and mitochondrial DNA sequences. Yusuke Takehana a,*, Kiyoshi Naruse b , Mitsuru Sakaizumi .Molecular Phylogenetics and Evolution 36 (2005) 417–428、[6]Coenye T, Vandamme P.在已测序细菌基因组中的多种16S rRNA操纵子的基因内接合的不均一性. FEMS Microbiol. Lett. November 2003, 228 (1): 45–9. PMID 14612235. doi:10.1016/S0378-1097(03)00717-1、[7]系统发育学研究中对16S rDNA扩增的运用W G Weisburg, S M Barns, D A Pelletier and D J Lane; J Bacteriol. 1991 January; 173(2): 697-703、[8] Woese, C. R.; G. E. Fox. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences. 1977-11-01, 74 (11): 5088–5090. Bibcode:1977PNAS...74.5088W. ISSN 0027-8424. PMC 432104. PMID 270744. doi:10.1073/pnas.74.11.5088、。