第二章 分子克隆工具酶
- 格式:pdf
- 大小:681.92 KB
- 文档页数:114

第二章分子克隆工具酶基因工程工具酶⏹限制性内切酶——主要用于DNA分子的特异切割⏹DNA甲基化酶——用于DNA分子的甲基化⏹核酸酶——用于DNA和RNA的非特异性切割⏹核酸聚合酶——用于DNA和RNA的合成⏹核酸连接酶——用于DNA和RNA的连接⏹核酸末端修饰酶——用于DNA和RNA的末端修饰⏹其它酶类——用于生物细胞的破壁,转化,核酸纯化,检测等。
第一节限制性核酸内切酶(restriction enzyme)一、限制性内切酶的发现历史核酸酶:催化核酸酯键的水解核酸外切酶:作用于核酸链的末端(3′或5 ′),逐个水解核苷酸。
核酸内切酶:从核酸分子内部切断3′,5′磷酸二酯键。
限制性核酸内切酶:在细菌细胞内存在的一类能识别并水解外源双链DNA的核酸内切酶。
1、细菌的限制——修饰现象细菌的限制——修饰作用发现细菌的限制(restriction)现象:寄主控制的专一性phageλKR-M 系统的作用:R-M 系统是细菌安内御外的积极措施。
保护自身的DNA不受制;破坏外源DNA使之迅速降解细菌R-M 系统的限制酶可以降解DNA,为避免自身DNA 的降解,细菌可以修饰(甲基化酶)自身DNA,未被修饰的外来DNA 则会被降解。
个别噬菌体在被降解之前已经发生了修饰,则可免予被降解。
限制性内切酶本是微生物细胞中用于专门水解外源DNA的一类酶,其功能是避免外源DNA的干扰或噬菌体的感染,是细胞中的一种防御机制。
由于R/M现象的发现使得核酸内切酶成为基因工程重要的工具酶。
2、限制性核酸内切酶的发现限制性核酸内切酶(restriction endonuelease)是一类能识别双链DNA中特殊核苷酸序列,并使每条链的一个磷酸二酯键断开的内脱氧核糖核酸酶(endo-deoxyribonuclease)。
它是可以将外来的DNA切断,即能够限制异源DNA的侵入并使之失去活力,但对自己的DNA却无损害作用,这样可以保护细胞原有的遗传信息。





第二章分子克隆工具酶题目第二章分子克隆工具酶题目第二章分子克隆工具酶一、选择题1.有关基因工程的描述恰当的就是()a.限制性内乌酶只在赢得目的基因时才用b.重组质粒的构成在细胞内顺利完成c.质粒都可为载运体d.蛋白质的结构可为合成目的基因提供资料2.限制性内乌酶ecorⅰ在野生型的λ噬菌体dna中存有5个切点,hindⅲ存有7个切点。
调整这些酶切位点的数量,主要通过()。
a.体内变异b.全然酶乌后相连接c.部分酶切d.先用甲基化酶修饰后再酶切二、填空题1.如果用限制性内乌核酸酶研磨双链dna产生5?注重的黏性末端,则可以用__________展开3?末端标记。
如果用限制性内乌核酸酶研磨dna产生的就是3?注重的黏性末端,可以用__________________展开3?末端标记。
2.可用t4dna聚合酶进行平末端的dna标记,因为这种酶具有____________和_______的活性。
三、判断题1.pbr327就是在pbr322的基础上换成了1089bp的片段,遗留下了ampr和tetr的完备片段。
pbr327比pbr322存有更高的拷贝,每个细胞中所含30-45个拷贝。
同时pbr327具备切割能力,能直接转入其他细胞()四、名词解释1.dna聚合酶2.限制性内乌酶3.甲基化酶4.t4-dna连接酶5.核酸酶6.klenow酶7.t4dna聚合酶8.taqdna聚合酶9.逆转录酶10.同裂酶11.核酶五、简答题1.限制性内切酶分为哪几大类,各有什么特点?2.dna修饰酶主要有哪些,各有什么作用?3.简述klenow聚合酶的用途。
4.比较e.colidna连接酶和taqdna连接酶的促进作用特点。
5.限制性内切酶i的识别序列和切点是―g?gatcc―,限制性内切酶ii的识别序列和切点是―?gatc―。
在质粒上有酶i的1个切点,在目的基因的两侧各有l个酶ii的切点。
用上述两种酶分别切割质粒和含有目的基因的dna。

第二章分子克隆工具酶DNA 重组技术中对核酸的“精雕细刻”主要用酶作为工具。
60年代末,70年代初科学家们陆续发现了DNA 连接酶、T4DNA 连接酶、Ⅱ型限制性核酸内切酶Hin fI 和Eco RI、反转录酶等,才使他们能对DNA 分子进行剪切、连接等基因操作,故称这些酶为工具酶。
到目前为止,世界上发现的工具酶种类和数量繁多,现将重组DNA 分子技术中常的工具酶简介如下。
限制性核酸内切酶(restriction endonuclease)在重组DNA 技术中有重要地位,在此较详细介绍。
第一节限制性核酸内切酶核酸酶可分为两类:核酸外切酶(exonuclease)是从核酸的一端开始,一个接一个把核苷酸水解下来;核酸内切酶(endonuclease则从核酸链中间水解3’,5’磷酸二酯键,将核酸链切断。
很多细菌和细胞中都能识别外来的核酸并将其分解,1962年发现这是因为细菌中含有特异的核酸内切酶,能识别特定的核酸序列而将核酸切断;同时又伴随有特定的核酸修饰酶,最常见的是甲基化酶,能使细胞自身核酸特定的序列上碱基甲基化,从而避免受内切酶水解,外来核酸没有这种特异的甲基化修饰,就会被细胞的核酸酶所水解.这样细胞就构成了限制一修饰体系,其功能就是保护自身的DNA,分解外来的DNA,以保护和维持自身遗传信息的稳定,这对细菌的生存和繁衍具有重要意义。
这就是限制性核酸内切酶名称中“限制”二字概念的由来。
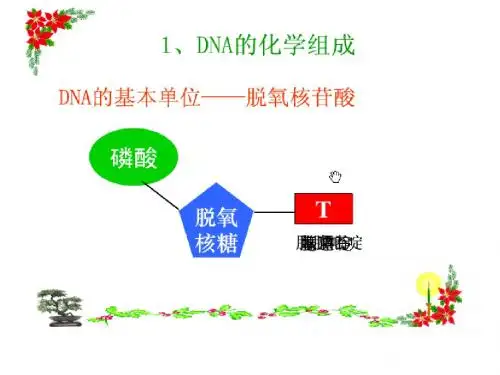
1.1 限制性核酸内切酶(restriction endonuclease1.1.1 限制性核酸内切酶的概念是一类能够识别双链DNA 分子中的某种特定核苷酸序列,并由此切割DNA 双链结构的核酸水解酶。
它们主要是从原核生物中分离纯化出来的。
到目前为止,已经分离出可识别230种不同DNA 序列的Ⅱ型限制性核酸内切酶达2300种以上。
在限制性核酸内切酶作用下,侵入细菌的“外源”DNA分子便会被切割成不同大小的片段,而细菌自己固有的DNA 碱基甲基化,在此修饰酶的保护下,则可免受限制性核酸内切酶的降解。

2 分子克隆工具酶(4学时)概述限制性内切核酸酶甲基化酶(Methylase)DNA聚合酶(DNA polymerase Ⅰ)其他分子克隆工具酶工具酶的定义o在生物技术中常用的各种工具酶系指能用于DNA和RNA分子的切割、连接、聚合、反转录等有关的各种酶系统称为工具酶。
o基因工程的操作,是在分子水平上的操作,是依赖一些酶(如限制性核酸内切酶,连接酶,DNA聚合酶等)作为工具对基因进行人工切割,拼接和扩增等操作。
所以把这些酶称之为“工具酶”。
o工具酶是对野生菌株(或真核生物如酵母)进行改造、优化、而产生的生物工程产品。
o自然界的许多微生物体内存在着一些具有特异功能的酶类。
这些酶类参与微生物的核酸代谢,在核酸复制和修复等反应中具有重要作用,有的酶还作为微生物区别自己和非己的DNA 进而降解非己DNA的防御工具。
o本章主要介绍用于基因工程的各种工具酶。
常用的工具酶2.1 限制性内切酶(restriction enzyme)o限制与修饰(Restriction and modification)o限制酶识别的序列o限制酶产生的末端o DNA末端长度对限制酶切割的影响o位点偏爱(Site preference)o酶切反应条件o星星活性(star activity)o单链DNA 的切割o酶切位点的引入o影响酶活性的因素o酶切位点在基因组中分布的不均一性E.coli k 感染频率下降许多E .coli B 【E .coli B 限制λ(k )】1. 限制与修饰现象①50年代后Luria 和Human(1952), Bertani 和Weigle (1953) 发现细菌的“限制”现象:2.1.1 限制与修饰(Restriction and modification )E.coli B 修饰λ(k)*λ(k)感染E.coli(B)细菌如何对λ噬菌体进行限制和修饰?E.coli B对这些λ(K)进行了修饰。
表2-1 λ噬菌体不同感染株对E.coli 不同菌株的感染率111E.coli C 10-4110-4E.coli B10-410-41E.coli KλC λB λK λ噬菌体感染率E.coli 菌株细菌的限制—修饰系统①1962年W.Arber(瑞士)发现,寄主对噬菌体的修饰是在λPhage的DNA上。
②1965年,Arber发现修饰与λ的降解有关,提出:细胞中存在位点特异性限制酶和特异性甲基化酶,即细胞中有限制—修饰系统(R-MRestriction-modification system)。
甲基化是常见的修饰作用,可使腺嘌呤A 成为N 6甲基-腺膘呤,胞嘧啶C 成为5‘甲基胞嘧啶。
R-M系统是细菌安内御外的积极措施。
2.限制性内切酶的发现•1968年,Meselson从E.coli K株中分离出了第一个限制酶Eco K,同年Linn和Aeber从E.coli B株中分离到限制酶Eco B。
•1970年,Smith和Wilcox从流感嗜血杆菌中分离到一种限制性酶Hin dⅡ,这是第一个分离到的Ⅱ类限制性内切核酸酶。
•HindⅡ限制性内切酶位点和切割位点如下:5 '…GTPy↓PuAC…3 '3 '…CAPu↑PyTG…5 '由宿主控制的限制与修饰作用限制性内切酶的命名o限制性酶采用三字母的命名原则,即属名+ 种名+ 株名的1个首字母,再加上序号,将限制性内切核酸酶的命名要点列于表。
表限制性内切核酸酶的命名要点Escherichia Coli Ry13E c o RⅠ属名种类株系编号3.限制与修饰系统的种类o根据酶的亚单位组成、识别序列的种类和是否需要辅助因子,限制与修饰系统至少可分为四类。
o在已纯化分类的3000多种限制性内切酶中,已发现了超过250种的特异识别序列。
表2-2 各种限制与修饰系统的比较YesYes No 限制作用是否需用ATP 同时竞争互斥分开的反应限制反应与甲基化反应在识别位点下游24-26bp 无特异性,至少在识别位点外1000bp 在识别位点中或靠近识别位点切割位点5-7bp 非对称非对称4-6bp, 大多数为回文对称结构识别位点二亚基双功能酶三亚基双功能酶(R ,M ,S 亚基) 内切酶与甲基化酶分子不在一起酶分子ⅢⅠⅡⅡ型(typeⅡ)限制与修饰系统o限制酶:一般是同源二聚体(homodimer),由两个彼此按相反方向结合在一起的相同亚单位组成,每个亚单位作用在DNA 链的两个互补位点上。
o修饰酶:单体,修饰作用一般由两个甲基转移酶来完成,分别作用于其中一条链,但甲基化的碱基在两条链上是不同的。
o它们识别回文对称序列(palindrome sequence),在回文序列内部或附近切割DNA,产生带3'-羟基和5'-磷酸基团的DNA 产物,需Mg2+,相应的修饰酶只需SAM 。
识别序列主要为4-6bp ,或更长且呈二重对称的特殊序列,但有少数酶识别更长的序列或简并序列,切割位置因酶而异。
oⅡs 型(type Ⅱs)限制与修饰系统,占5% ,与Ⅱ型具有相似的辅因子要求,但识别位点是非对称,也是非间断的,长度为4-7bp ,切割位点可能在识别位点一侧的20bp 范围内。
Ⅰ型(typeⅠ)限制与修饰系统o其限制酶和甲基化酶(即R亚基和M亚基)各作为一个亚基存在于酶分子中,另外还有负责识别DNA序列的S亚基,分别由hsdR、hsdM和hsdS基因编码,属于同一操纵子(转录单位)。
如Eco K编码基因的结构为R2M2S,Eco B 编码基因的结构为R2M4S2 。
o Eco B 酶的识别位点如下,其中两条链中的A *为甲基化位点,N表示任意碱基。
TGA*(N)8TGCTo Eco K 酶的识别位点如下,其中两条链中的A°为可能的甲基化位点。
AA°C(N)6GTGCo但是Eco B 酶和Eco K 酶的切割位点在识别位点1000bp 以外,且无特异性。
hsd: host specificity for DNA restrictionI类酶的作用方式(引自Lewin,1997)o在一定序列上识别DNA分子, 并能同DNA分子作用, 因其识别DNA后, 要朝一个方向或两个方向移动一段距离(通常为1000个碱基左右),并且要形成一个环才能切割DNA, 所以识别位点和切割位点不一致,产生的片段较大。
Ⅲ型(type Ⅲ)限制与修饰系统o种类更少,所占比例不到1% ,如Eco P1 和Eco P15 。
它们的识别位点分别是AGACC 和CAGCAG ,切割位点则在下游24-26bp 处。
o Eco P1是由两个亚基组成,一个亚基(M亚基)负责位点识别和修饰。
另一个亚基(R亚基)具有核酸酶的活性。
切割DNA时需要ATP,Mg2+,也能被SAM激活,但并非必需。
oⅢ类限制性内切核酸酶(引自Lewin,1997)2.1.2 限制酶识别的序列1.限制酶识别序列的长度o限制酶识别序列的长度一般为4-8个碱基,最常见的为6个碱基。
o当识别序列为4个和6个碱基时,它们可识别的序列在完全随机的情况下,平均每256个和4096个碱基中会出现一个识别位点(44=256,46=4096)。
o注意:识别位点存在的概率不同!2.限制酶识别序列的结构o限制酶识别的序列大多数为回文对称结构,切割位点在DNA两条链相对称的位置。
o特例:(见下页举例)识别序列不是对称的识别多种序列识别的序列呈间断对称,对称序列之间含有若干个任意碱基。
Eco RⅠG↓AATTC Hin dⅢA↓AGCTTCTTAA↑G TTCGA↑A识别序列不是对称的Acc BSⅠCCG↓CTC Bss SⅠC↓TCGTGGGC↑GAG GAGCA↑C识别多种序列AccⅠGT↓MKAC,M=A或C, K=G或T识别的序列呈间断对称Alw NⅠCAGNNNC↓TG DdeⅠC↓TNAG GT↑CNNNGAC GANT↑C3.限制酶切割的位置o限制酶对DNA的切割位置大多数在内部,但也有在外部的。
在外部的,又有两端、两侧和单侧之别。
2.1.3 限制酶产生的末端1.限制酶产生匹配粘端(matched end)o识别位点为回文对称结构的序列经限制酶切割后,产生的末端为匹配黏端,亦即黏性末端(cohesive end)。
o若在对称轴5′侧切割底物,DNA双链交错断开产生5′突出黏性末端;o若在3′侧切割,则产生3′突出黏性末端。
NNG↓AATTCNN NNCTTAA↑GNN Eco RⅠNNG+NNCTTAAAATTCNNGNN5’----NCTGCA↓GN----3’PstI5’----NCTGCA 3’----NG↑ACGTCN----5’3’----NGGN----3’ACGTCN----5’2.限制酶产生平末端(Blunt end)o在回文对称轴上同时切割DNA的两条链,则产生平末端,如HaeⅢ(GG↓CC)和Eco R V(GAT↓ATC)。
5’----NCCC ↓GGGN----3’SmaI5’----NCCC 3’----NGGG ↑CCCN----5’3’----NGGG +CCCN----5’GGGN----3’平末端o产生平末端的DNA可任意连接,但连接效率较黏性末端低。
3.限制酶产生非对称突出末端o当识别序列为非对称序列时,切割的DNA产物的末端是不同的,如Bbv CⅠ,它的识别切割位点如下:CC↓TCAGCGGAGT↑CGo有些限制酶识别简并序列,其识别的序列中有几种是非对称的。
如AccⅠ,它的识别切割位点如下:GT↓AT/CGACCATA/GC↑TGo有些限制酶识别间隔序列,间隔区域的序列是任意的,如DraⅢ,识别切割位点是CAC↑NNN↓GTG。
4.同裂酶(isoschizomer)o识别相同序列的限制酶称同裂酶,但它们的切割位点可能不同。
①同序同切酶识别序列和切割位置都相同,如HpaⅡ与MapⅠ识别切割位点为C↓CGG。
②同序异切酶KpnⅠ和Acc65Ⅰ识别的序列是相同的,但切割位点不同,分别为GGTAC↓C和G↓GTACC。
③“同功多位”许多识别简并序列的限制酶包含了另一种限制酶的功能。
如Eco RⅠ为G↓AATTC,ApoⅠ为R↓AATTY,后者可识别前者的序列。
④其他有些限制酶识别的序列有交叉,如在pUC系列质粒的多克隆位点(multiple cloning site, MCS)中有一个SalⅠ位点(G↓TCGAC),该位点也可被AccⅠ(GT↓MKAC)和HincⅡ(GTY↓RAC)切割。
R=A,G Y=C,T5.同尾酶o许多不同的限制酶切割DNA产生的末端是相同的,且是对称的,即它们可产生相同的黏性突出末端。
这些酶统称为同尾酶(isocaudarner)。