T细胞免疫图片
- 格式:pptx
- 大小:2.17 MB
- 文档页数:25

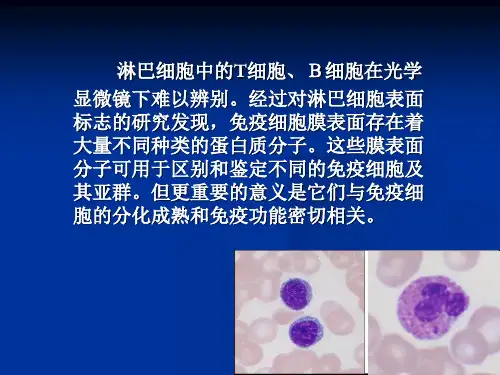



免疫细胞分型常见标志物大串烧(一)——T细胞分型标志物免疫分型(immunophenotyping)是指根据存在于细胞表面、核或细胞质中的标记物或抗原的类型,利用基于抗体的对异质细胞群进行分析,以鉴定多种目标细胞群的存在及比例。
通过使用特异性识别不同细胞类型标记物或抗原的抗体,从而帮助鉴定细胞的谱系。
例如肿瘤标志物的检测、浸润免疫细胞的分析、白血病的诊断等。
免疫分型可以利用组织切片(新鲜或固定组织)通过免疫组化方法以及利用细胞悬液通过流式细胞术进行分析。
流式细胞术可以说是用于探测人类免疫表型的首选技术,特别是由于它可以在诸如血液的复杂混合物中通过测量许多单个细胞上的多个参数,从而鉴定和分析包括稀有亚群在内的许多细胞亚群。
并且由于可用的抗体试剂的扩大和实验方案的不断调整,流式细胞术不仅可用于评估细胞表面蛋白的表达,还可以用于评估胞内蛋白的表达或修饰水平以及细胞因子的水平。
使用细胞表面标记的特定组合对免疫细胞亚群进行定义是一个不断发展的过程,特别是那些高频研究的细胞类型。
这些类型包括调节性T细胞(Treg)、分泌IL-17的T辅助细胞(Th17)、树突状细胞(DC)和自然杀伤细胞(NK)。
尽管新标记的发现和新的细胞子集将在未来一段时间内持续下去,然而,我们还是希望细胞免疫学领域具有足够的成熟程度,以达到对大多数共同研究的免疫细胞亚群的定义达成共识。
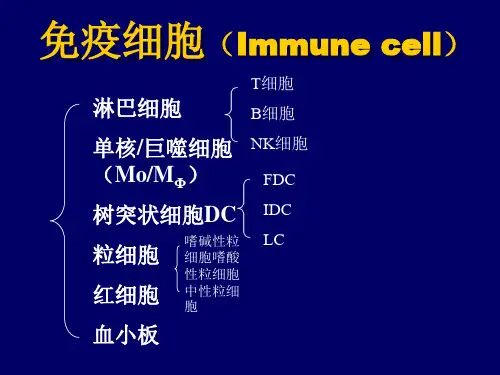
换句话说,应当能够定义一种相对稳定的标记集合,其描绘T细胞、B细胞、NK细胞、单核细胞和DC细胞等主要类型。
因此,我们通过总结文献确定各个细胞类型的关键分化标记,并且总结目前用于人体免疫监测的标准化免疫表型分型组合。
T细胞T淋巴细胞可在CD3表达的基础上被免疫表型化,随后进一步细分(如CD8+杀伤性T细胞和CD4+辅助性T细胞)。
此外,T细胞表型是灵活的,可以随不同的微环境发生变化;表型也可能重叠在多个T细胞群体之间。
此外,T细胞亚群的分类还可能是根据响应某些刺激或磷酸化免疫信号蛋白(如STAT蛋白)而分泌的特定细胞因子水平来确定。

T细胞卡罗林斯卡医学院研究结果表明,与已经产⽣抗体的⼈相⽐,对新冠病毒免疫的所谓T细胞,免疫⼒是其⼤约两倍。
通过对新冠肺炎中重症的患者的检测表明,他们⼤部分⼈表现出强⼤的T细胞防御能⼒和抗体防御能⼒。
卡罗林斯卡医学院在⼀份新闻稿中写道,虽然在症状较轻的⼈中,并不总是能够发现他们是否已产⽣抗体,但是许多患者都有T细胞。
T细胞是⼀种⽩细胞,专门识别被病毒感染的细胞,是我们免疫系统的重要组成部分。
我是T细胞,免疫细胞家族中的杀⼿。
免疫细胞家族中从不缺杀⼿,我的兄弟B细胞,巨噬细胞都是杀⼿。
但我不⼀样,我是家族⾥的“头号杀⼿”。
我能够分清外来的感染源、组织以及⾃⾝的癌细胞将他们直接杀死;有时候忙不过,我也会分泌淋巴激素“叫醒”其他的免疫细胞,帮我⼀起杀敌;杀完了敌⼈,我也能及时收⼿,从不恋战,他们说我拥有“调节或抑制免疫”的能⼒。
正是因为我如此“机智能⼲”,业务能⼒超群,我们免疫家族中⼀半的功勋章都是由我打下的。
不是我吹,如今⼤热的免疫疗法中的两⼤顶梁柱:PD-1/PD-L1抑制剂和CAR-T疗法都是以我为基础⽽诞⽣的。
这不,国内刚刚获批的两⼤“抗癌神药”——Opdivo和Keytruda都是我的孩⼦。
⼀、虽然我瞎,但我有天线.要说我为什么能这么厉害,这还得感谢我的母亲——⾻髓⼲细胞,把我⽣的如此与众不同。
听说我出⽣的那⼀刻,天上下起了太阳⾬,算命先⽣说这可是天⼤的吉兆。
但我们T细胞占尽优势,却唯独有⼀个缺点——视⼒不好。
没有办法靠着⾃⼰的眼睛来发现癌细胞,只有靠着别的细胞找来了癌细胞的信息——特异性抗原,才能顺藤摸⽠的找到犯罪分⼦“癌细胞”,将其⼀击毙命。
所幸的是,虽然我们瞎,但我们每个T细胞出⽣时都会随⾝携带着⼀个信号天线——TCR。
有了它就能做到“⼈在家中坐,知晓天下事”。
同时,我们正是通过信号天线接受到了其他细胞送过来的信息。
正所谓“千⾥送鹅⽑,礼轻情意重”,正是因为我们免疫家族的团结⼀致,互帮互助,才能多年连续蝉联癌症“克星”的美誉,我的成功⾃然也少不了其他兄弟姐妹的帮助和⽀持。

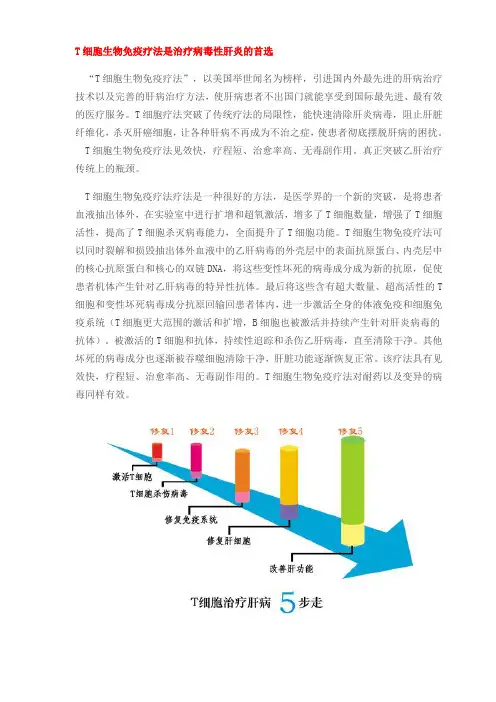
T细胞生物免疫疗法是治疗病毒性肝炎的首选“T细胞生物免疫疗法”,以美国举世闻名为榜样,引进国内外最先进的肝病治疗技术以及完善的肝病治疗方法,使肝病患者不出国门就能享受到国际最先进、最有效的医疗服务。
T细胞疗法突破了传统疗法的局限性,能快速清除肝炎病毒,阻止肝脏纤维化,杀灭肝癌细胞,让各种肝病不再成为不治之症,使患者彻底摆脱肝病的困扰。
T细胞生物免疫疗法见效快,疗程短、治愈率高、无毒副作用。
真正突破乙肝治疗传统上的瓶颈。
T细胞生物免疫疗法疗法是一种很好的方法,是医学界的一个新的突破,是将患者血液抽出体外,在实验室中进行扩增和超氧激活,增多了T细胞数量,增强了T细胞活性,提高了T细胞杀灭病毒能力,全面提升了T细胞功能。
T细胞生物免疫疗法可以同时裂解和损毁抽出体外血液中的乙肝病毒的外壳层中的表面抗原蛋白、内壳层中的核心抗原蛋白和核心的双链DNA,将这些变性坏死的病毒成分成为新的抗原,促使患者机体产生针对乙肝病毒的特异性抗体。
最后将这些含有超大数量、超高活性的T 细胞和变性坏死病毒成分抗原回输回患者体内,进一步激活全身的体液免疫和细胞免疫系统(T细胞更大范围的激活和扩增,B细胞也被激活并持续产生针对肝炎病毒的抗体)。
被激活的T细胞和抗体,持续性追踪和杀伤乙肝病毒,直至清除干净。
其他坏死的病毒成分也逐渐被吞噬细胞清除干净,肝脏功能逐渐恢复正常。
该疗法具有见效快,疗程短、治愈率高、无毒副作用的。
T细胞生物免疫疗法对耐药以及变异的病毒同样有效。
1. T细胞生物免疫疗法治疗肝病的优势:一、有效促进乙肝表面抗原与E抗原转阴:转阴效果明显优于其他乙肝疗法。
二、持续增强机体的抗病毒能力:与药物短暂疗效相比,随着体内的变性坏死病毒数量的增加,这些坏死病毒作为新的抗原,进一步刺激机体免疫系统产生更多活化T细胞和病毒抗体,杀灭更多病毒,直至杀灭干净。
三、有效逆转肝纤维化:对早期肝硬化有良好的治疗效果,具有保肝护肝作用。
四、不需要任何药物,不产生任何副作用:用自体细胞治疗治疗自身的疾病,不需要任何药物,不产生任何毒副作用,是真正意义上的绿色治疗。

•·NGF信号通路(2004-8-16)•·TGF beta 信号转导(2004-8-16)•·细胞凋亡信号(2004-8-16)•·线粒体输入信号(2004-8-16)•·ROS信号(2004-8-16)•·Toll-Like 受体家族(2004-8-16)•·Toll-Like 受体(2004-8-16)•·actin肌丝(2004-8-16)•·Wnt/LRP6 信号(2004-8-16)•·WNT信号转导(2004-8-16)•·West Nile 西尼罗河病毒(2004-8-16)•·Vitamin C 维生素C在大脑中的作用(2004-8-16)•·视觉信号转导(2004-8-16)•·VEGF,低氧(2004-8-16)•·TSP-1诱导细胞凋亡(2004-8-16)•·Trka信号转导(2004-8-16)•·dbpb调节mRNA (2004-8-16)•·CARM1甲基化(2004-8-16)•·CREB转录因子(2004-8-16)•·TPO信号通路(2004-8-16)•·Toll-Like 受体(2004-8-16)•·TNFR2 信号通路(2004-8-16)•·TNFR1信号通路(2004-8-16)•·TNF/Stress相关信号(2004-8-16)•·IGF-1受体(2004-8-16)•·共刺激信号(2004-8-16)•·Th1/Th2 细胞分化(2004-8-16)•·TGF beta 信号转导(2004-8-16)•·端粒、端粒酶与衰老(2004-8-16)•·TACI和BCMA调节B细胞免疫(2004-8-16)•·T辅助细胞的表面受体(2004-8-16)•·T细胞受体信号通路(2004-8-16)•·T细胞受体和CD3复合物(2004-8-16)•·Cardiolipin的合成(2004-8-16)•·Synaptic突触连接中的蛋白(2004-8-16)•·HSP在应激中的调节的作用(2004-8-16)•·Stat3 信号通路(2004-8-16)•·SREBP控制脂质合成(2004-8-16)•·酪氨酸激酶的调节(2004-8-16)•·Sonic Hedgehog (SHH)受体ptc1调节细胞周期(2004-8-16)•·Sonic Hedgehog (Shh) 信号(2004-8-16)•·SODD/TNFR1信号(2004-8-16)•·AKT/mTOR在骨骼肌肥大中的作用(2004-8-16)•·G蛋白信号转导(2004-8-16)•·肝细胞生长因子受体信号(2004-8-16)•·IL1受体信号转导(2004-8-16)•·acetyl从线粒体到胞浆过程(2004-8-16)•·趋化因子chemokine在T细胞极化中的选择性表(2004-8-16)•·SARS冠状病毒蛋白酶(2004-8-16)•·Parkin在泛素-蛋白酶体中的作用(2004-8-16)•·nicotinic acetylcholine受体在凋亡中的作用(2004-8-16)•·线粒体在细胞凋亡中的作用(2004-8-16)•·MEF2D在T细胞凋亡中的作用(2004-8-16)•·Erk5和神经元生存(2004-8-16)•·ERBB2信号转导(2004-8-16)•·GPCRs调节EGF受体(2004-8-16)•·BRCA1调节肿瘤敏感性(2004-8-16)•·Rho细胞运动的信号(2004-8-16)•·Leptin能逆转胰岛素抵抗(2004-8-16)•·转录因子DREAM调节疼敏感(2004-8-16)•·PML调节转录(2004-8-16)•·p27调节细胞周期(2004-8-16)•·MAPK信号调节(2004-8-16)•·细胞因子调节造血细胞分化(2004-8-16)•·eIF4e和p70 S6激酶调节(2004-8-16)•·eIF2调节(2004-8-16)•·谷氨酸受体调节ck1/cdk5 (2004-8-16)•·plk3在细胞周期中的作用(2004-8-1)•·BAD磷酸化调节(2004-8-1)•·Reelin信号通路(2004-8-1)•·RB肿瘤抑制和DNA破坏(2004-8-1)•·NK细胞介导的细胞毒作用(2004-8-1)•·Ras信号通路(2004-8-1)•·Rac 1细胞运动信号(2004-8-1)•·PTEN依赖的细胞生长抑制和细胞凋亡(2004-8-1)•·notch信号通路(2004-8-1)•·蛋白激酶A(PKA)在中心粒中的作用(2004-8-1)•·蛋白酶体Proteasome复合物(2004-8-1)•·Prion朊病毒的信号通路(2004-8-1)•·早老素Presenilin在notch和wnt信号中的作用(2004-8-1)•·mRNA的poly(A)形成(2004-8-1)•·淀粉样蛋白前体信号(2004-8-1)•·PKC抑制myosin磷酸化(2004-8-1)•·磷脂酶C(PLC)信号(2004-8-1)•·巨噬细胞Pertussis toxin不敏感的CCR5信号通(2004-8-1)•·Pelp1调节雌激素受体的活性(2004-8-1)•·PDGF信号通路(2004-8-1)•·p53信号通路(2004-8-1)•·p38MAPK信号通路(2004-8-1)•·Nrf2是氧化应激基本表达的关键基因(2004-8-1)•·OX40信号通路(2004-8-1)•·hTerc转录调节活性图(2004-8-1)•·hTert转录因子的调节作用(2004-8-1)•·AIF在细胞凋亡中的作用(2004-8-1)•·Omega氧化通路(2004-8-1)•·核受体在脂质代谢和毒性中的作用(2004-8-1)•·NK细胞中NO2依赖的IL-12信号通路(2004-8-1)•·TOR信号通路(2004-8-1)•·NO信号通路(2004-8-1)•·NF-kB信号转导通路(2004-8-1)•·NFAT与心肌肥厚的示意图(2004-8-1)•·神经营养素及其表面分子(2004-8-1)•·神经肽VIP和PACAP防止活化T细胞凋亡图(2004-8-1)•·神经生长因子信号图(2004-8-1)•·线虫和哺乳动物的MAPK信号比较(2004-7-17)•·细胞内信号总论(2004-7-17)•·细胞凋亡信号通路(2004-7-17)•·MAPK级联通路(2004-7-17)•·MAPK信号通路图(2004-7-17)•·BCR信号通路(2004-7-17)•·蛋白质乙酰化示意图(2004-7-17)•·wnt信号通路(2004-7-17)•·胰岛素受体信号通路(2004-7-17)•·细胞周期在G2/M期的调控机理图(2004-7-17)•·细胞周期G1/S检查点调控机理图(2004-7-17)•·Jak-STAT关系总表(2004-7-17)•·Jak/STAT 信号(2004-7-17)•·TGFbeta信号(2004-7-17)•·NFkappaB信号(2004-7-17)•·p38 MAPK信号通路(2004-7-17)•·SAPK/JNK 信号级联通路(2004-7-17)•·从G蛋白偶联受体到MAPK (2004-7-17)•·MAPK级联信号图(2004-7-17)•·eIF-4E和p70 S6激酶调控蛋白质翻译(2004-7-17)•·eif2蛋白质翻译(2004-7-17)•·蛋白质翻译示意图(2004-7-17)•·线粒体凋亡通路(2004-7-17)•·死亡受体信号通路(2004-7-17)•·凋亡抑制通路(2004-7-17)•·细胞凋亡综合示意图(2004-7-17)•·Akt/Pkb信号通路(2004-7-17)•·MAPK/ERK信号通路(2004-7-17)•·哺乳动物MAPK信号通路(2004-7-17)•·Pitx2多步调节基因转录(2004-7-17)•·IGF-1R导致BAD磷酸化的多个凋亡路径(2004-7-17)•·多重耐药因子(2004-7-17)•·mTOR信号通路(2004-7-17)•·Msp/Ron受体信号通路(2004-7-17)•·单核细胞和其表面分子(2004-7-17)•·线粒体的肉毒碱转移酶(CPT)系统(2004-7-17)•·METS影响巨噬细胞的分化(2004-7-17)•·Anandamide,内源性大麻醇的代谢(2004-7-17)•·黑色素细胞(Melanocyte)发育和信号(2004-7-17)•·DNA甲基化导致转录抑制的机理图(2004-7-17)•·蛋白质的核输入信号图(2004-7-17)•·PPARa调节过氧化物酶体的增殖(2004-7-17)•·对乙氨基酚(Acetaminophen)的活性和毒性机(2004-7-17)•·mCalpain在细胞运动中的作用(2004-7-17)•·MAPK信号图(2004-7-17)•·MAPK抑制SMRT活化(2004-7-17)•·苹果酸和天门冬酸间的转化(2004-7-17)•·低密度脂蛋白(LDL)在动脉粥样硬化中的作用(2004-7-17)•·LIS1基因在神经细胞的发育和迁移中的作用图(2004-7-17)•·Pyk2与Mapk相连的信号通路(2004-7-17)•·galactose代谢通路(2004-7-17)•·Lectin诱导补体的通路(2004-7-17)•·Lck和Fyn在TCR活化中的作用(2004-7-17)•·乳酸合成图(2004-7-17)•·Keratinocyte分化图(2004-7-17)•·离子通道在心血管内皮细胞中的作用(2004-7-17)•·离子通道和佛波脂(Phorbal Esters)信号(2004-7-17)•·内源性Prothrombin激活通路(2004-7-17)•·Ribosome内化通路(2004-7-17)•·整合素(Integrin)信号通路(2004-7-17)•·胰岛素(Insulin)信号通路(2004-7-17)•·Matrix Metalloproteinases (2004-7-17)•·组氨酸去乙酰化抑制剂抑制Huntington病(2004-7-17)•·Gleevec诱导细胞增殖(2004-7-17)•·Ras和Rho在细胞周期的G1/S转换中的作用(2004-7-17)•·DR3,4,5受体诱导细胞凋亡(2004-7-17)•·AKT调控Gsk3图(2004-7-17)•·IL-7信号转导(2004-7-17)•·IL22可溶性受体信号转导图(2004-7-17)•·IL-2活化T细胞图(2004-7-17)•·IL12和Stat4依赖的TH1细胞发育信号通路(2004-7-17)•·IL-10信号通路(2004-7-17)•·IL 6信号通路(2004-7-17)•·IL 5信号通路(2004-7-17)•·IL 4信号通路(2004-7-17)•·IL 3信号通路(2004-7-17)•·IL 2 信号通路(2004-7-17)•·IL 18信号通路(2004-7-17)•·IL 17信号通路(2004-7-17)•·IGF-1信号通路(2004-7-17)•·IFN gamma信号通路(2004-7-17)•·INF信号通路(2004-7-17)•·低氧诱导因子(HIF)在心血管中的作用(2004-7-17)•·低氧和P53在心血管系统中的作用(2004-7-17)•·人类巨细胞病毒和MAP信号通路(2004-7-17)•·孕酮如何促进卵细胞成熟?(2004-7-17)•·How does salmonella hijack a cell (2004-7-17)•·Hop通路在心脏发育中的作用(2004-7-17)•·HIV-I Nef:负性调节fas和TNF (2004-7-17)•·HIV-1防止宿主细胞耐受的机理(2004-7-17)•·HIV诱导T细胞凋亡图(2004-7-17)•·血红素的伴侣分子(2004-7-17)•·g-Secretase介导ErbB4信号通路(2004-7-17)•·生物激素信号(2004-7-17)•·Granzyme A介导的凋亡信号通路(2004-7-17)•·G蛋白偶联信号需要Tubby支持(2004-7-17)•·糖酵解通路(2004-7-17)•·Ghrelin:食物吸收和能量平衡的调控者(2004-7-17)•·PS1能产生beta淀粉样蛋白导致老年性痴呆(2004-7-17)•·GATA3部分参与TH2细胞因子基因的表达(2004-7-17)•·GABA受体的代谢图(2004-7-17)•·FXR和LXR调节胆固醇代谢(2004-7-17)•·SLRP在骨骼中的作用(2004-7-17)•·自由基诱导细胞凋亡信号(2004-7-17)•·FOSB与药物成瘾(2004-7-17)•·fMLP诱导趋化因子基因表达(2004-7-17)•·Fibrinolysis通路(2004-7-17)•·糖酵解通路(2004-7-17)•·Fc Epsilon Receptor I信号(2004-7-17)•·FAS信号通路(2004-7-17)•·外源性Prothrombin激活通路(2004-7-17)•·真核细胞蛋白质翻译示意图(2004-7-17)•·雌激素反应蛋白EFP控制乳腺癌细胞的细胞周期(2004-7-17)•·EPO介导神经保护作用与NF-kB相关(2004-7-17)•·Erythrocyte分化通路(2004-7-17)•·Erk1/Erk2 Mapk 信号通路(2004-7-17)•·Erk和PI-3K在细胞外间质中的作用(2004-7-17)•·内质网相关的蛋白质降解通路示意图(2004-7-17)•·EPO售转导机制图(2004-7-17)•·血小板凝聚示意图(2004-7-17)•·NDK动力学(2004-7-17)•·线粒体的电子传递链示意图(2004-7-17)•·Eicosanoid代谢(2004-7-17)•·EGF信号通路(2004-7-17)•·calcineurin对Keratinocyte分化的影响(2004-7-17)•·E2F1信号通路(2004-7-17)•·MTA-3在雌激素不敏感性乳腺癌中下调(2004-7-17)•·双链RNA诱导基因表达示意图(2004-7-17)•·Dicer信号通路(RNAi机理)(2004-7-17)•·CDK5在老年性痴呆中的调节作用(2004-7-17)•·树突状细胞调节TH1和TH2发育示意图(2004-7-17)•·RAR和RXR被蛋白酶体降解通路(2004-7-17)•·D4-GDI信号通路示意图(2004-7-17)•·细胞因子和炎症反应示意图(2004-7-9)•·细胞因子网络调控图(2004-7-9)•·CFTR和beta 2肾上腺素受体通路(2004-7-9)•·Cyclin和细胞周期调控图(2004-7-9)•·Ran核质循环转运图(2004-7-9)•·Cyclin E降解通路图(2004-7-9)•·CXCR4信号通路图(2004-7-9)•·CTL介导的免疫反应图(2004-7-9)•·CTCF:第一个多价核因子(2004-7-9)•·皮质激素和心脏保护(2004-7-9)•·骨骼肌的成肌信号图(2004-7-9)•·VitD调控基因表达信号图(2004-7-9)•·补体信号通路(2004-7-9)•·线粒体和过氧化物酶体中β氧化的比较图(2004-7-9)•·经典的补体信号通路图(2004-7-9)•·心律失常的分子机制图(2004-7-9)•·hSWI/SNF ATP依赖的染色体重塑(2004-7-9)•·碳水化合物和cAMP调节ChREBP图(2004-7-9)•·分子伴侣调节干扰素信号图(2004-7-9)•·Ceramide信号图(2004-7-9)•·局部急性感染的细胞与分子信号(2004-7-9)•·细胞与细胞粘附信号(2004-7-9)•·细胞周期G2/M调控点信号调节(2004-7-9)•·细胞周期 G1/S调控点信号图(2004-7-9)•·CDK调节DNA复制(2004-7-9)•·cdc25和chk1在DNA破坏中的作用图(2004-7-9)•·CD40L信号通路图(2004-7-9)•·CCR3信号图(2004-7-9)•·CBL下调EGF受体的信号转导图(2004-7-9)•·一些氨基酸的代谢通路图 3 (2004-7-9)•·一些氨基酸的代谢通路图 2 (2004-7-9)•·一些氨基酸的代谢通路图(2004-7-9)•·Catabolic pathway for asparagine and asp (2004-7-9)•·Caspase 信号级联通路在细胞凋亡中的作用(2004-7-9)•·CARM1和雌激素的信号转导调控(2004-7-9)•·抗氧自由基的心脏保护作用信号转导图(2004-7-9)•·乙肝病毒中的钙信号调控(2004-7-9)•·镉诱导巨噬细胞的DNA合成和增殖(2004-7-9)•·Ca2+/CaM依赖的激活(2004-7-9)•·B细胞活化机理图(2004-6-9)•·BTG家族蛋白和细胞周期的调节(2004-6-9)•·BRCA1作用机理(2004-6-9)•·骨重塑示意图(2004-6-9)•·Botulinum Toxin阻断神经递质释放示意图(2004-6-9)•·缬氨酸的生物合成图(2004-6-9)•·Tryptophan在植物和细菌内的生物合成(2004-6-9)•·苏氨酸和蛋氨酸的体内合成示意图(2004-6-9)•·sphingolipids生物合成(2004-6-9)•·spermidine和spermine生物合成(2004-6-9)•·细菌体内合成脯氨酸的示意图(2004-6-9)•·苯丙氨酸和酪氨酸的生物合成(2004-6-9)•·神经递质的合成示意图(2004-6-9)•·赖氨酸生物合成图(2004-6-9)•·亮氨酸的体内生物合成图(2004-6-9)•·异亮氨酸的生物合成图(2004-6-9)•·甘氨酸和色氨酸的生物合成(2004-6-9)•·Cysteine在哺乳动物中的合成图(2004-6-9)•·Cysteine在细菌和植物内生物合成图(2004-6-9)•·Chorismate在细菌和植物内的生物合成(2004-6-9)•·Arginine在细菌内的生物合成(2004-6-9)•·生物活性肽诱导的通路(2004-6-9)•·脂肪酸的β氧化通路(2004-6-9)•·BCR信号通路示意图(2004-6-9)•·SUMOylation基本机理(2004-6-9)•·PPAR影响基因表达的基本信号机制图(2004-6-9)•·B淋巴细胞表面分子示意图(2004-6-9)•·B细胞生存信号通路(2004-6-5)•·B细胞信号通路的复杂性(2004-6-5)•·GPCR信号的衰减的机理(2004-6-4)•·ATM信号通路(2004-6-4)•·阿斯匹林的抗凝机理(2004-6-4)•·细胞凋亡信号调节DNA片段化(2004-6-4)•·细胞凋亡DNA片段化与组织稳态的机理(2004-6-4)•·反义核酸的作用机理---RNA polymerase III (2004-6-4)•·抗原递呈与处理信号图(2004-6-4)•·Antigen依赖的B细胞激活(2004-6-4)•·Anthrax Toxin Mechanism of Action (2004-6-4)•·血管紧张素转换酶2调节心脏功能(2004-6-4)•·Angiotensin II 介导JNK信号通路的激活(2004-6-4)•·Alternative Complement Pathway (2004-6-4)•·Alpha-synuclein和Parkin在怕金森病中的作用(2004-6-4)•·ALK在心肌细胞中的功能图(2004-6-4)•·AKT信号通路(2004-6-4)•·AKAP95在有丝分裂中的作用图(2004-6-4)•·Ahr信号转导图(2004-6-4)•·Agrin突触后的功能图(2004-6-4)•·ADP-Ribosylation 因子(2004-6-4)•·淋巴细胞粘附分子信号图(2004-6-4)•·Adhesion and Diapedesis of Lymphocytes (2004-6-4)•·Adhesion and Diapedesis of Granulocytes (2004-6-4)•·急性心肌梗死信号转导图(2004-6-4)•·src蛋白质激活图(2004-6-4)•·PKC与G蛋白耦联受体的关系(2004-6-4)•·cAMP依赖的CSK抑制T细胞功能示意图(2004-6-4)•·PKA功能示意图(2004-6-4)•·一氧化氮(NO)在心脏中的功能示意图(2004-6-4)•·RelA 在细胞核内乙酰化和去乙酰化(2004-6-4)actin肌丝Mammalian cell motility requires actin polymerization in the direction of movement to change membrane shape and extend cytoplasm into lamellipodia. The polymerization of actin to drive cell movement also involves branching of actin filaments into a network oriented with the growing ends of the fibers near the cell membrane. Manipulation of this process helps bacteria like Salmonella gain entry into cells they infect. Two of the proteins involved in the formation of Y branches and in cell motility are Arp2 and Arp3, both members of a large multiprotein complex containing several other polypeptides as well. The Arp2/3 complex is localized at the Y branch junction and induces actin polymerization. Activity of this complex is regulated by multiple different cell surface receptor signaling systems, activating WASP, and Arp2/3 in turn to cause changes in cell shape and cell motility. Wasp and its cousin Wave-1 interact with the Arp2/3 complex through the p21 component of the complex. The crystal structure of the Arp2/3 complex has revealed further insights into the nature of how the complex works.Activation by Wave-1, another member of the WASP family, also induces actin alterations in response to Rac1 signals upstream. Wave-1 is held in an inactive complex in the cytosol that is activated to allow Wave-1 to associate with Arp2/3. While WASP is activated by interaction with Cdc42, Wave-1, is activated by interaction with Rac1 and Nck. Wave-1 activation by Rac1 and Nck releases Wave-1 with Hspc300 to activate actin Y branching and polymerization by Arp2/3. Different members of this gene family may produce different actin cytoskeletal architectures. The immunological defects associated with mutation of the WASP gene, theWiskott-Aldrich syndrome for which WASP was named, indicates the importance of this system for normal cellular function.Cory GO, Ridley AJ. Cell motility: braking WAVEs. Nature. 2002 Aug 15;418(6899):732-3. No abstract available.Eden, S., et al. (2002) Mechanism of regulation of WAVE1-induced actin nucleation by Rac1 and Nck. Nature 418(6899), 790-3Falet H, Hoffmeister KM, Neujahr R, Hartwig JH. Normal Arp2/3 complex activation in platelets lacking WASp. Blood. 2002 Sep 15;100(6):2113-22.Kreishman-Deitrick M, Rosen MK, Kreishman-Deltrick M. Ignition of a cellular machine. Nat Cell Biol. 2002 Feb;4(2):E31-3. No abstract available.Machesky, L.M., Insall, R.H. (1998) Scar1 and the related Wiskott-Aldrich syndrome protein, WASP, regulate the actin cytoskeleton through the Arp2/3 complex. Curr Biol 8(25), 1347-56Robinson, R.C. et al. (2001) Crystal structure of Arp2/3 complex. Science 294(5547), 1679-84Weeds A, Yeoh S. Structure. Action at the Y-branch. Science. 2001 Nov 23;294(5547):1660-1. No abstract available.Wnt/LRP6 信号Wnt glycoproteins play a role in diverse processes during embryonic patterning in metazoa through interaction with frizzled-type seven-transmembrane-domain receptors (Frz) to stabilize b-catenin. LDL-receptor-related protein 6 (LRP6), a Wnt co-receptor, is required for this interaction. Dikkopf (dkk) proteins are both positive and negative modulators of this signalingWNT信号转导West Nile 西尼罗河病毒West Nile virus (WNV) is a member of the Flaviviridae, a plus-stranded virus family that includes St. Louis encephalitis virus, Kunjin virus, yellow fever virus, Dengue virus, and Japanese encephalitis virus. WNV was initially isolated in 1937 in the West Nile region of Uganda and has become prevalent in Africa, Asia, and Europe. WNV has rapidly spread across the United States through its insect host and causes neurological symptoms and encephalitis, which can result in paralysis or death. Since 1999 about 3700 cases of West Nile virus (WNV) infection and 200 deaths have been recorded in United States. The viral capsid protein likely contributes to the WNV-associated deadly inflammation via apoptosis induced through the mitochondrial pathway.WNV particles (50 nm in diameter) consist of a dense core (viral protein C encapsidated virus RNA genome)surrounded by a membrane envelope (viral E and M proteins embedded in a lipid bilayer). The virus binds to a specific cell surface protein (not yet identified), an interaction thought to involve E protein with highly sulfated neperan sulfate (HSHS) residues that are present on the surfaces of many cells and enters the cell by a process similar to that of endocytosis. Once inside the cell, the genome RNA is released into the cytoplasm via endosomal release, a fusion process involving acidic pH induced conformation change in the E protein. The RNA genome serves as mRNA and is translated by ribosomes into ten mature viral proteins are produced via proteolytic cleavage, which include three structural components and seven different nonstructural components of the virus. These proteins assemble and transcribe complimentary minus strand RNAs from the genomic RNA. The complimentary minus strand RNA in turns serves as template for the synthesis of positive-stranded genomic RNAs. Once viral E, preM and C proteins have accumulated to sufficient level, they assemble with the genomic RNA to form progeny virions, which migrate to the cell surface where they are surrounded with lipid envelop and released.Vitamin C 维生素C在大脑中的作用Vitamin C (ascorbic acid) was first identified by virtue of the essential role it plays in collagen modification, preventing the nutritional deficiency scurvy. Vitamin C acts as a cofactor for hydroxylase enzymes thatpost-translationally modify collagen to increase the strength and elasticity of tissues. Vitamin C reduces the metal ion prosthetic groups of many enzymes, maintaining activity of enzymes, also acts as an anti-oxidant. Although the prevention of scurvy through modification of collagen may be the most obvious role for vitamin C, it is not necessarily the only role of vitamin C. Svct1 and Svct2 are ascorbate transporters for vitamin C import into tissues and into cells. Both of these transporters specifically transport reduced L-ascorbic acid against a concentration gradient using the intracellular sodium gradient to drive ascorbate transport. Svct1 is expressed in epithelial cells in the intestine, upregulated in cellular models for intestinal epithelium and appears to be responsible for the import of dietary vitamin C from the intestinal lumen. The vitamin C imported from the intestine is present in plasma at approximately 50 uM, almost exclusively in the reduced form, and is transported to tissues to play a variety of roles. Svct2 imports reduced ascorbate from the plasma into veryactive tissues like the brain. Deletion in mice of the gene for Svct2 revealed that ascorbate is required for normal development of the lungs and brain during pregnancy. A high concentration of vitamin C in neurons of the developing brain may help protect the developing brain from free radical damage. The oxidized form of ascorbate, dehydroascorbic acid, is transported into a variety of cells by the glucose transporter Glut-1. Glut-1, Glut-3 and Glut-4 can transport dehydroascorbate, but may not transport significant quantities of ascorbic acid in vivo.视觉信号转导信息来源:本站原创生物谷网站The signal transduction cascade responsible for sensing light in vertebrates is one of the best studied signal transduction processes, and is initiated by rhodopsin in rod cells, a member of the G-protein coupled receptor gene family. Rhodopsin remains the only GPCR whose structure has been resolved at high resolution. Rhodopsinin the discs of rod cells contains a bound 11-cis retinal chromophore, a small molecule derived from Vitamin A that acts as the light sensitive portion of the receptor molecule, absorbing light to initiate the signal transduction cascade. When light strikes 11-cis retinal and is absorbed, it isomerizes to all-trans retinal, changing the shape of the molecule and the receptor it is bound to. This change in rhodopsin抯shape alters its interaction with transducin, the member of the G-protein gene family that is specific in its role in visual signal transduction. Activation of transducin causes its alpha subunit to dissociate from the trimer and exchange bound GDP for GTP, activating in turn a membrane-bound cyclic-GMP specific phosphodiesterase that hydrolyzes cGMP. In the resting rod cell, high levels of cGMP associate with a cyclic-GMP gated sodium channel in the plasma membrane, keeping the channels open and the membrane of the resting rod cells depolarized. This is distinct from synaptic generation of action potentials, in which stimulation induces opening of sodium channels and depolarization. When cGMP gated channels in rod cells open, both sodium and calcium ions enter the cell, hyperpolarizing the membrane and initiating the electrochemical impulse responsible for conveying the signal from the sensory neuron to the CNS. The rod cell in the resting state releases high levels of the inhibitory neurotransmitter glutamate, while the release of glutamate is repressed by the hyperpolarization in the presence of light to trigger a downstream action potential by ganglion cells that convey signals to the brain. The calcium which enters the cell also activates GCAP, which activates guanylate cyclase (GC-1 and GC-2) to rapidly produce more cGMP, ending the hyperpolarization and returning the cell to its resting depolarized state. A protein called recoverin helps mediate the inactivation of the signal transduction cascade, returning rhodopsin to its preactivated state, along with the rhodopsin kinase Grk1. Phosphorylation of rhodopsin by Grkl causes arrestin to bind, helping to terminate the receptor activation signal. Dissociation and reassociation of retinal, dephosphorylation of rhodopsin and release of arrestin all return rhodopsin to its ready state, prepared once again to respond to light.VEGF,低氧信息来源:本站原创生物谷网站Vascular endothelial growth factor (VEGF) plays a key role in physiological blood vessel formation and pathological angiogenesis such as tumor growth and ischemic diseases. Hypoxia is a potent inducer of VEGF in vitro. The increase in secreted biologically active VEGF protein from cells exposed to hypoxia is partly because of an increased transcription rate, mediated by binding of hypoxia-inducible factor-1 (HIF1) to a hypoxia responsive element in the 5'-flanking region of the VEGF gene. bHLH-PAS transcription factor that interacts with the Ah receptor nuclear translocator (Arnt), and its predicted amino acid sequence exhibits significant similarity to the hypoxia-inducible factor 1alpha (HIF1a) product. HLF mRNA expression is closely correlated with that of VEGF mRNA.. The high expression level of HLF mRNA in the O2 delivery system of developing embryos and adult organs suggests that in a normoxic state, HLF regulates gene expression of VEGF, various glycolytic enzymes, and others driven by the HRE sequence, and may be involved in development of blood vessels and the tubularsystem of lung. VEGF expression is dramatically induced by hypoxia due in large part to an increase in the stability of its mRNA. HuR binds with high affinity and specificity to the VRS element that regulates VEGF mRNA stability by hypoxia. In addition, an internal ribosome entry site (IRES) ensures efficient translation of VEGF mRNA even under hypoxia. The VHL tumor suppressor (von Hippel-Lindau) regulates also VEGF expression at a post-transcriptional level. The secreted VEGF is a major angiogenic factor that regulates multiple endothelial cell functions, including mitogenesis. Cellular and circulating levels of VEGF are elevated in hematologic malignancies and are adversely associated with prognosis. Angiogenesis is a very complex, tightly regulated, multistep process, the targeting of which may well prove useful in the creation of novel therapeutic agents. Current approaches being investigated include the inhibition of angiogenesis stimulants (e.g., VEGF), or their receptors, blockade of endothelial cell activation, inhibition of matrix metalloproteinases, and inhibition of tumor vasculature. Preclinical, phase I, and phase II studies of both monoclonal antibodies to VEGF and blockers of the VEGF receptor tyrosine kinase pathway indicate that these agents are safe and offer potential clinical utility in patients with hematologic malignancies.TSP-1诱导细胞凋亡信息来源:本站原创生物谷网站As tissues grow they require angiogenesis to occur if they are to be supplied with blood vessels and survive. Factors that inhibit angiogenesis might act as cancer therapeutics by blocking vessel formation in tumors and starving cancer cells. Thrombospondin-1 (TSP-1) is a protein that inhibits angiogenesis and slows tumor growth, apparently by inducing apoptosis of microvascular endothelial cells that line blood vessels. TSP-1 appears to produce this response by activating a signaling pathway that begins with its receptor CD36 at the cell surface of the microvascular endothelial cell. The non-receptor tyrosine kinase fyn is activated by TSP-1 through CD36, activating the apoptosis inducing proteases like caspase-3 and p38 protein kinases. p38 is a mitogen-activated kinase that also induces apoptosis in some conditions, perhaps through AP-1 activation and the activation of genes that lead to apoptosis.Trka信号转导信息来源:本站原创生物谷网站Nerve growth factor (NGF) is a neurotrophic factor that stimulates neuronal survival and growth through TrkA, a member of the trk family of tyrosine kinase receptors that also includes TrkB and TrkC. Some NGF responses are also mediated or modified by p75LNTR, a low affinity neurotrophin receptor. Binding of NGF to TrkA stimulates neuronal survival, and also proliferation. Pathways coupled to these responses are linked to TrkAthrough association of signaling factors with specific amino acids in the TrkA cytoplasmic domain. Cell survival through inhibition of apoptosis is signaled through activation of PI3-kinase and AKT. Ras-mediated signaling and phospholipase C both activate the MAP kinase pathway to stimulate proliferation.dbpb调节mRNA信息来源:本站原创生物谷网站Endothelial cells respond to treatment with the protease thrombin with increased secretion of the PDGF B-chain. This activation occurs at the transcriptional level and a thrombin response element was identified in the promoter of the PDGF B-chain gene. A transcription factor called the DNA-binding protein B (dbpB) mediates the activation of PDGF B-chain transcription in response to thrombin treatment. DbpB is a member of the Y box family of transcription factors and binds to both RNA and DNA. In the absence of thrombin, endothelial cells contain a 50 kD form of dbpB that binds RNA in the cytoplasm and may play a role as a chaperone for mRNA. The 50 kD version of dbpB also binds DNA to regulate genes containing Y box elements in their promoters. Thrombin activation results in the cleavage of dbpB to a 30 kD form. The proteolytic cleavage releases dbpB from RNA in the nucleus, allowing it to enter the nucleus and binds to a regulatory element distinct from the site recognized by the full length 50 kD dbpB. The genes activated by cleaved dbpB include the PDGF B chain. Dephosphorylation of dbpB also regulates nuclear entry and transcriptional activation.RNA digestion in vitro can release dbpB in its active form, suggesting that the protease responsible for dbpB may be closely associated in a complex. Identification of the protease that cleaves dbpB, the mechanisms of phosphorylation and dephosphorylation, and elucidation of the signaling path by which thrombin induces dbpB will provide greater understanding of this novel signaling pathway.CARM1甲基化信息来源:本站原创生物谷网站Several forms of post-translational modification regulate protein activities. Recently, protein methylation by CARM1 (coactivator-associated arginine methyltransferase 1) has been observed to play a key role in transcriptional regulation. CARM1 associates with the p160 class of transcriptional coactivators involved in gene activation by steroid hormone family receptors. CARM1 also interacts with CBP/p300 transcriptional coactivators involved in gene activation by a large variety of transcription factors, including steroid hormone receptors and CEBP. One target of CARM1 is the core histones H3 and H4, which are also targets of the histone acetylase activity of CBP/p300 coactivators. Recruitment of CARM1 to the promoter region by binding to coactivators increases histone methylation and makes promoter regions more accessible for transcription. Another target of CARM1 methylation is a coactivator it interacts with, CBP. Methylation of CBP by CARM1 blocks。