鸟氨酸、瓜氨酸和精氨酸发酵
- 格式:doc
- 大小:387.00 KB
- 文档页数:8

第十二章鸟氨酸、瓜氨酸和精氨酸发酵L-精氨酸(L-Arginine, L-Arg)是具有胍基的碱性氨基酸,是合成蛋白质和肌酸的重要原料。
精氨酸是人体和动物体的半必需氨酸,在医药和食品工业上具有广泛的用途。
L-鸟氨酸(L-Ornithine, L-Orn)和L-瓜氨酸(L-Citrulline, L-Cit)是精氨酸生物合成的前体物质。
L-精氨酸是生物体尿素循环中的一种重要中间代谢产物,临床上除作为复合氨基酸输液的主要成分之一外,L-精氨酸及其盐类广泛用作氨中毒性肝昏迷的解毒剂和肝功能促进剂。
对病毒性肝炎疗效显著,对肠道溃疡、血栓形成、神经衰弱和男性无精病等症都有治疗效果。
它也是配制营养支持用或特殊治疗用要素膳的重要原料。
第一节生物合成途径和代谢调节机制一、鸟氨酸、瓜氨酸、精氨酸的相互转化图12-l 鸟氨酸、瓜氨酸、精氨酸的结构从结构(见图12-1)上看,鸟氨酸虽与谷氨酸都是五碳酸,但是鸟氨酸却是一羧基二氨基的氨基酸。
如果在鸟氨酸末端氨基的氮上接上氨甲酰基,则生成瓜氨酸。
瓜氨酸经过精氨琥珀酸,将瓜氨酸的酮基转换成亚氨基,则成为精氨酸。
当然,精氨酸放出尿素,就转变为鸟氨酸。
因此,鸟氨酸、瓜氨酸和精氨酸的生物合成,可认为是从谷氨酸出发,逐步合成鸟氨酸、瓜氨酸和精氨酸,从而组成以精氨酸为最终产物的不分支代谢途径。
但是,如果精氨酸发生分解,放出尿素,则生成鸟氨酸。
这样在代谢途径上,就使所谓的终产物精氨酸与鸟氨酸相衔接,形成一个循环,即尿素循环或鸟氨酸环(图12-2)。
图12-2 尿素循环二、生物合成途径和代谢调节机制从谷氨酸经鸟氨酸、瓜氨酸生物合成L-精氨酸,由八个酶催化的反应组成(见图12-3),第一步和第五步反应因微生物的种类不同而不同。
谷氨酸N-乙酰谷氨乙酰谷氨酸磷酸乙酰谷氨酸半醛N-乙酰鸟氨鸟氨酸精氨酰琥珀NADPHNADP×酸α-酮瓜氨门冬氨×氨基甲酰磷酸××⑴⑶⑸⑹精氨酸反丁烯二酸I型大肠杆菌、枯草杆菌×谷氨酸N-乙酰谷氨酸N-乙酰谷氨酸磷酸N-乙酰谷氨酸半醛N-乙酰鸟氨酸精氨酰琥珀酸精氨酸××××⑴⑵⑶⑷鸟氨酸瓜氨酸×⑸⑹⑺⑻×反馈抑制×阻遏×可能存在的阻遏II型谷氨酸产生菌、酵母菌×图12-3 L-精氨酸生物合成途径及调节机制大肠杆菌、枯草杆菌等微生物由图12-3 I的途径合成L-精氨酸,称为I型途径。


《氨基酸工艺学》教学大纲一、课程基本信息二、课程性质和目的氨基酸工艺学是一门新型发酵的技术科学,以探讨氨基酸发酵工厂的生产技术为主要目的。
学习”氨基酸工艺学”的目的是使学生能运用已学过的微生物学、生物化学、化工原理和分析化学等基础知识,进一步深化与提高,来认识与解决氨基酸发酵工业生产中的具体问题。
三、课程教学的基本要求通过本课程的学习是学生掌握选育氨基酸生产菌的基本原理,了解氨基酸代谢与代谢控制发酵的基本理论、发酵控制的关键及分离精制氨基酸的一般原理与方法,从而使学生初步具有选育新菌种、探求新工艺、新装备和从事氨基酸发酵研究的能力。
四、课程教学基本内容1. 绪论⑴概述⑵氨基酸发酵的历史与发展动向⑶我国氨基酸行业存在的问题和解决措施2. 淀粉水解糖的制备⑴淀粉的组成及其特性⑵淀粉水解糖的制备方法⑶双酶法制糖工艺3. 谷氨酸发酵机制⑴谷氨酸生产菌、谷氨酸生物合成的调节机制⑵谷氨酸发酵过程中细胞渗透性的控制4. 谷氨酸生产菌的特征、育种及扩大培养⑴谷氨酸生产菌的主要特征、国内谷氨酸生产菌及其比较、谷氨酸生产菌及其比较、⑵谷氨酸发酵的代谢控制育种策略、生物工程新技术育种、菌种的扩大培养及种子的质量要求5. 谷氨酸发酵过程控制⑴发酵培养基、培养条件对谷氨酸发酵的影响⑵发酵过程中主要变化及中间代谢控制,泡沫的消除⑶糖蜜原料强制发酵工艺、低糖流加工艺及后期补糖工艺⑷提高发酵产率的主要措施6. 噬菌体与杂菌的防治⑴谷氨酸发酵中的噬菌体的污染与防治⑵杂菌的污染与防治7. 谷氨酸的提取⑴概论⑵谷氨酸发酵液的性质⑶等电点法提取谷氨酸、离子交换法提取谷氨酸、等电点-离子交换法提取谷氨酸、深缩连续等电点法提取谷氨酸⑷谷氨酸发酵液的综合利用8. 谷氨酸制味精⑴味精的性质⑵谷氨酸制味精的工艺流程⑶谷氨酸的中和与除铁,谷氨酸中和液的脱色、浓缩、结晶⑷味精生产中异常现象及其处理9. 谷氨酸清洁生产工艺⑴谷氨酸清洁生产工艺概述⑵提取谷氨酸闭路循环工艺⑶发酵废母液提取菌体蛋白工艺、生产饲料酵母工艺、发酵母液浓缩生产复合有机肥工艺、发酵废水生物处理工艺10. 氨基酸发酵机制⑴概述⑵酶活性的调控、酶合成的调控⑶氨基酸生物合成的调节机制11. 氨基酸产生菌的选育与发酵技术⑴氨基酸产生菌的选育与定向育种策略、用细胞内基因重组手段选育氨基酸产生菌、用重组DNA技术构建氨基酸工程菌、氨基酸产生菌的稳定化⑵发酵条件对氨基酸发酵的影响⑶氨基酸的提取与精制12. 天冬氨酸族氨基酸发酵⑴天冬氨酸族氨基酸生物合成途径⑵天冬氨酸族氨基酸生产菌的选育及发酵机制⑶影响天冬氨酸族氨基酸发酵的主要因素⑷赖氨酸发酵、苏氨酸发酵、蛋氨酸发酵、高丝氨酸发酵、天冬氨酸发酵13. 鸟氨酸、瓜氨酸和精氨酸发酵14. 异亮氨酸、亮氨酸与缬氨酸发酵15. 色氨酸、苯丙氨酸和酪氨酸发酵16. 丙氨酸、脯氨酸、谷氨酸胺和组氨酸发酵五、学时分配表六、建议教材与教学参考书1.《氨基酸工艺学》,陈宁,中国轻工业出版社,2007年1月七、课程考核方式与成绩评定本课程采取的考核方式如:考试、考查、论文等。

鸟氨酸循环记忆口诀稿子一嘿,亲爱的小伙伴们!今天咱们来聊聊鸟氨酸循环这个有点小复杂,但超重要的知识点。
先来说说这个循环的步骤吧。
鸟氨酸循环,从鸟氨酸开始,就像一场奇妙的旅行。
氨和二氧化碳是它的小伙伴,一起加入了这场大冒险。
鸟氨酸先和氨还有二氧化碳,合成了瓜氨酸,这就像是旅程的第一站。
然后瓜氨酸又和天冬氨酸结伴,变成了精氨酸代琥珀酸。
这一步就像是走到了中途的一个小景点,有点精彩哦。
接着,精氨酸代琥珀酸裂解,了精氨酸。
这感觉就像到达了一个重要的转折点。
精氨酸水解,又变回了鸟氨酸,同时释放出尿素。
哇塞,这一轮循环就圆满结束啦!那怎么记住这个过程呢?我有个小口诀要分享给大家:“鸟氨携碳合瓜氨,瓜氨遇酸成代琥,代琥裂解出精氨,精氨水解回鸟氨,尿素伴随把家还。
”是不是感觉一下子就清晰多啦?小伙伴们,记住这个口诀,鸟氨酸循环就再也难不倒咱们啦!加油哦!稿子二哈喽呀,友友们!今天来唠唠鸟氨酸循环,别怕,不难的哟!咱们就把鸟氨酸循环想象成一个有趣的故事。
鸟氨酸是主角,它带着氨和二氧化碳这两个小伙伴出发啦。
一开始,它们仨一见面,就变成了瓜氨酸,这是故事的开头。
接着,瓜氨酸碰到了天冬氨酸,哎呀,它们一组合,就成了精氨酸代琥珀酸,这情节是不是越来越有意思啦?然后呢,精氨酸代琥珀酸一拆分,精氨酸就出现了。
就好像是隐藏的角色登场啦。
呀,精氨酸被水解,鸟氨酸又回来了,还顺便把尿素给带出来了。
为了记住这个好玩的过程,咱们有个口诀哟:“鸟氨带着俩伙伴,合成瓜氨第一步。
瓜氨遇见天冬酸,变成代琥接着走。
代琥分裂出精氨,精氨水解鸟氨归,尿素也来凑热闹。
”怎么样,友友们,有了这个口诀,是不是觉得鸟氨酸循环变得简单又有趣啦?咱们一起多念叨念叨,把它牢牢记住!。
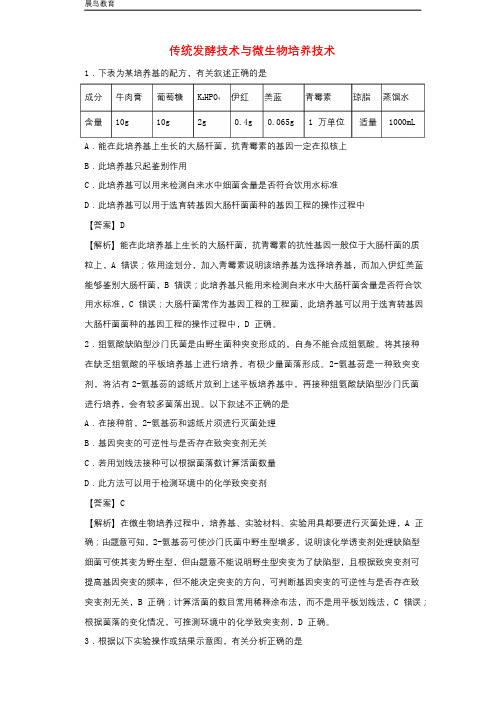
传统发酵技术与微生物培养技术1.下表为某培养基的配方,有关叙述正确的是成分牛肉膏葡萄糖K2HPO4 伊红美蓝青霉素琼脂蒸馏水含量10g 10g 2g 0.4g 0.065g 1万单位适量1000mL A.能在此培养基上生长的大肠杆菌,抗青霉素的基因一定在拟核上B.此培养基只起鉴别作用C.此培养基可以用来检测自来水中细菌含量是否符合饮用水标准D.此培养基可以用于选育转基因大肠杆菌菌种的基因工程的操作过程中【答案】D【解析】能在此培养基上生长的大肠杆菌,抗青霉素的抗性基因一般位于大肠杆菌的质粒上,A错误;依用途划分,加入青霉素说明该培养基为选择培养基,而加入伊红美蓝能够鉴别大肠杆菌,B错误;此培养基只能用来检测自来水中大肠杆菌含量是否符合饮用水标准,C错误;大肠杆菌常作为基因工程的工程菌,此培养基可以用于选育转基因大肠杆菌菌种的基因工程的操作过程中,D正确。
2.组氨酸缺陷型沙门氏菌是由野生菌种突变形成的,自身不能合成组氨酸。
将其接种在缺乏组氨酸的平板培养基上进行培养,有极少量菌落形成。
2-氨基芴是一种致突变剂,将沾有2-氨基芴的滤纸片放到上述平板培养基中,再接种组氨酸缺陷型沙门氏菌进行培养,会有较多菌落出现。
以下叙述不正确的是A.在接种前,2-氨基芴和滤纸片须进行灭菌处理B.基因突变的可逆性与是否存在致突变剂无关C.若用划线法接种可以根据菌落数计算活菌数量D.此方法可以用于检测环境中的化学致突变剂【答案】C【解析】在微生物培养过程中,培养基、实验材料、实验用具都要进行灭菌处理,A正确;由题意可知,2-氨基芴可使沙门氏菌中野生型增多,说明该化学诱变剂处理缺陷型细菌可使其变为野生型,但由题意不能说明野生型突变为了缺陷型,且根据致突变剂可提高基因突变的频率,但不能决定突变的方向,可判断基因突变的可逆性与是否存在致突变剂无关,B正确;计算活菌的数目常用稀释涂布法,而不是用平板划线法,C错误;根据菌落的变化情况,可推测环境中的化学致突变剂,D正确。

第二节提高初级代谢产物产量的方法我们知道,初级和次级代谢产物在遗传控制、合成途径等方面存在差异的,因而获得发酵过量生产的方法也不同。
由于次级代谢产物的合成远离初级代谢的主要途径,微生物细胞对其合成控制较弱,因此,改变环境条件易于影响其表达,基因型改变后的产量变异幅度也较大,而初级代谢产物则与此相反。
这在选择提高代谢产物方法时应予考虑。
提高初级代谢产物产量的方法主要有以下几种:1.使用诱导物与糖类和蛋白质降解有关的水解酶类大都属诱导酶类,因此向培养基中加入诱导物就会增加胞外酶的产量。
如加入槐糖(1,2-β-D-葡二糖)诱导木霉菌的纤维素酶的生成,木糖诱导半纤维素酶和葡萄糖异构酶的生成等。
但诱导物的价格往往比较贵,经济上未必合算。
加入廉价的含有诱导物的原料,如槐豆荚等某些种籽皮中含有槐糖,玉米芯富含木聚糖,培养过程中可陆续被水解产生槐糖、木糖,这就是经常采用的方法。
但是,玉米芯等这类不溶性聚合物的分解过程缓慢,以其唯一碳源时,培养周期比较长,产品的体积生产率仍难大幅度提高。
可考虑先使微生物在廉价的可溶性碳源中迅速生长,形成大量菌体后,再加入诱导物诱导水解酶类生成的方法。
诱导物的浓度过高及能被迅速利用时,也会发生酶合成的阻遏,这在纤维二糖对纤维素酶的产生,对二糖对半纤维素酶生产中都已观察到,这也是使用诱导物时应予注意的。
2.除去诱导物——选育组成型产生菌在发酵工业中,要选择到一种廉价、高效的诱导物是不容易的,分批限量加入诱导物在工艺上也多不便,更为有效的方法是改变菌株的遗传特性,除去对诱导物的需要,即选育组成型突变株。
通过诱变处理,使调节基因发生突变,不产生有活性的阻遏蛋白,或者操纵基因发生突变不再能与阻遏物相结合,都可达到此目的。
迄今尚未见由于结构基因发生改变而得到组成型的报道。
已设计出多种选育组成型突变株的方法,其主要原则是创造一种利于组成型菌株生长而不利于诱导型菌株生长的培养条件,造成对组成型的选择优势以及适当的识别两类菌落的方法,从而把产生的组成型突变株选择出来。
第十二章鸟氨酸、瓜氨酸和精氨酸发酵L-精氨酸(L-Arginine, L-Arg)是具有胍基的碱性氨基酸,是合成蛋白质和肌酸的重要原料。
精氨酸是人体和动物体的半必需氨酸,在医药和食品工业上具有广泛的用途。
L-鸟氨酸(L-Ornithine, L-Orn)和L-瓜氨酸(L-Citrulline, L-Cit)是精氨酸生物合成的前体物质。
L-精氨酸是生物体尿素循环中的一种重要中间代谢产物,临床上除作为复合氨基酸输液的主要成分之一外,L-精氨酸及其盐类广泛用作氨中毒性肝昏迷的解毒剂和肝功能促进剂。
对病毒性肝炎疗效显著,对肠道溃疡、血栓形成、神经衰弱和男性无精病等症都有治疗效果。
它也是配制营养支持用或特殊治疗用要素膳的重要原料。
第一节生物合成途径和代谢调节机制一、鸟氨酸、瓜氨酸、精氨酸的相互转化图12-l 鸟氨酸、瓜氨酸、精氨酸的结构从结构(见图12-1)上看,鸟氨酸虽与谷氨酸都是五碳酸,但是鸟氨酸却是一羧基二氨基的氨基酸。
如果在鸟氨酸末端氨基的氮上接上氨甲酰基,则生成瓜氨酸。
瓜氨酸经过精氨琥珀酸,将瓜氨酸的酮基转换成亚氨基,则成为精氨酸。
当然,精氨酸放出尿素,就转变为鸟氨酸。
因此,鸟氨酸、瓜氨酸和精氨酸的生物合成,可认为是从谷氨酸出发,逐步合成鸟氨酸、瓜氨酸和精氨酸,从而组成以精氨酸为最终产物的不分支代谢途径。
但是,如果精氨酸发生分解,放出尿素,则生成鸟氨酸。
这样在代谢途径上,就使所谓的终产物精氨酸与鸟氨酸相衔接,形成一个循环,即尿素循环或鸟氨酸环(图12-2)。
图12-2 尿素循环二、生物合成途径和代谢调节机制从谷氨酸经鸟氨酸、瓜氨酸生物合成L-精氨酸,由八个酶催化的反应组成(见图12-3),第一步和第五步反应因微生物的种类不同而不同。
谷氨酸N-乙酰谷氨乙酰谷氨酸磷酸乙酰谷氨酸半醛N-乙酰鸟氨鸟氨酸精氨酰琥珀NADPHNADP×酸α-酮瓜氨门冬氨×氨基甲酰磷酸××⑴⑶⑸⑹精氨酸反丁烯二酸I型大肠杆菌、枯草杆菌×谷氨酸N-乙酰谷氨酸N-乙酰谷氨酸磷酸N-乙酰谷氨酸半醛N-乙酰鸟氨酸精氨酰琥珀酸精氨酸××××⑴⑵⑶⑷鸟氨酸瓜氨酸×⑸⑹⑺⑻×反馈抑制×阻遏×可能存在的阻遏II型谷氨酸产生菌、酵母菌×图12-3 L-精氨酸生物合成途径及调节机制大肠杆菌、枯草杆菌等微生物由图12-3 I的途径合成L-精氨酸,称为I型途径。
谷氨酸产生菌、酵母菌等微生物由图12-3 II的途径合成L-精氨酸,称为II途径。
两种途径的区别在于I型途径由谷氨酸生成N-乙酰谷氨酸,由N-乙酰鸟氨酸生成鸟氨酸,分别由N-乙酰谷氨酸合成酶和N-乙酰鸟氨酸酶催化;而II型途径由谷氨酸合成N-乙酰谷氨酸,由N-乙酰鸟氨酸生成鸟氨酸,是由N-乙酰谷氨酸和乙酰鸟氨酸乙酰基转移酶共轭反应催化,即II型途径中有一个从谷氨酸到鸟氨酸的循环反应。
由于微生物合成L-精氨酸的途径不同,故其合成的调节机制也不同。
具有I型合成途径的大肠杆菌等,其生物合成途径的第一个酶,即N-乙酰谷氨酸合成酶受到精氨酸的反馈抑制,其途径上催化八步反应的酶的合成都受到精氨酸的阻遏。
具有II型途径的谷氨酸产生菌等,其生物合成的第二个酶,即N-乙酰谷氨酸激酶受到精氨酸的反馈抑制,其合成途径中各步酶的合成,大部分受到精氨酸的阻遏。
第二节鸟氨酸和瓜氨酸发酵一、鸟氨酸发酵1.由谷氨酸产生菌发酵生产鸟氨酸S. Udaka等(1957年)把谷氨酸棒杆菌的瓜氨酸缺陷(Cit-)突变株,在限量精氨酸的培养基中培养,成功地积累了大量的鸟氨酸,其产量高达36%(对糖分子收率)。
该突变株缺失鸟氨酸氨甲酰基转移酶,阻断了瓜氨酸、精氨酸的生成,这就解除了精氨酸对N-乙酰谷氨酸激酶的反馈抑制,也就使鸟氨酸氨甲酰基转移酶的底物——鸟氨酸大量积累。
Kinoshita等发现由谷氨酸棒杆菌选育精氨酸营养缺陷突变株,可高产L-鸟氨酸。
当L-精氨酸亚适量时,10%葡萄糖可产L-鸟氨酸25g/L。
陈宁等以谷氨酸棒杆菌为出发菌株,经硫酸二乙酯和紫外线多次诱变,定向选育出一株鸟氨酸生产菌A 1157,其遗传标记为Arg-+D-Arg r+SG r,并能耐受高浓度糖。
该菌株切断了由鸟氨酸向精氨酸的代谢途径,并解除了精氨酸对鸟氨酸合成酶系的反馈调控,从而可以大量积累鸟氨酸。
采用乳糖发酵短杆菌、柠檬酸节杆菌等的精氨酸缺陷突变株,均可在限量供给精氨酸的培养基中大量积累L-鸟氨酸(见表12-1)。
表12-1 L-鸟氨酸产生菌遗传标记及产量菌株遗传标记底物L-鸟氨酸(g·L-1)乳糖发酵短杆菌柠檬节杆菌枯草杆菌谷氨酸小球菌Arg-Arg-ArgHx r、TRA rArg-、ArgHx r葡萄糖葡萄糖葡萄糖葡萄糖62403548注:ArgHx—精氨酸氧肟酸,TRA—1,2,4-三唑-3-丙氨酸2.由其它菌株的突变株生产鸟氨酸枯草杆菌、大肠杆菌、产气气杆菌、变形杆菌(Proteus reftgeri)的精氨酸缺陷型(Arg —)菌株,在适当培养条件下,可由葡萄糖发酵生产鸟氨酸。
另外,解烃棒杆菌(Corynebacterium hydrocarbodastus)和石蜡节杆菌(Arthrobacter paraffineus)的Arg—菌株,可以在以正石蜡为主要碳源的培养基中生产鸟氨酸。
洁白链霉菌(Streptomyoes virginiae)的赖氨酸和硫胺素双重缺陷型菌株,在适当条件下,可由糖质生产10g/L的鸟氨酸。
3.发酵条件S. Udaka等在研究谷氨酸棒杆菌Cit—或Arg—菌株的鸟氨酸发酵时发现,为了使鸟氨酸产生菌充分生长,必需在培养基中适当添加菌株要求的物质精氨酸。
如果精氨酸的添加量不断增加,则菌体增殖达到顶点,但是其所生产的氨基酸种类却发生改变。
如表12-2所示,当精氨酸添加量达到300mg/L以上时,鸟氨酸生成量减少,相反地,谷氨酸生成量却增大了。
表12-2 由营养源浓度引起的生成氨基酸的变化营养源添加生成氨基酸*(g/L)精氨酸(mg/L)生物素(μg/L)鸟氨酸谷氨酸200 50052.5562.50.518注:*把谷氨酸棒杆菌(生物素缺陷型)的Cit-菌株,培养在含有萄葡糖10%,尿素0.3%,NH4Cl 1.0%的无机合成培养基申,培养3天的结果。
二、瓜氨酸发酵瓜氨酸和鸟氨酸一样,是精氨酸生物合成的中间体,所以可采用精氨酸营养缺陷型菌株进行瓜氨酸发酵。
从提高生产性和便于发酵管理出发,也可以采用精氨酸营养缺陷型、精氨酸结构类似物抗性及嘧啶抗性的营养缺陷和多重抗性突变株生产瓜氨酸。
1.由精氨酸营养缺陷型菌株生产瓜氨酸奥村等最先报告了大量积累瓜氨酸的研究,把枯草杆菌K的精氨酸营养缺陷型菌株培养在含有葡萄糖13%的发酵培养基中,发酵3天,生成瓜氨酸l6.5g/L。
在瓜氨酸发酵中,精氨酸是精氨酸营养缺陷型菌株的要求物质。
只有当精氨酸亚适量时,瓜氨酸积累量才最大。
瓜氨酸发酵的最佳精氨酸添加量为0.5g/L左右。
NH4C1是良好的氮源,宜在培养过程中分次添加。
瓜氨酸在菌体生长进入恒定期时,急速生成。
瓜氨酸的对糖分子收率最高为13.7%。
荻野等报告了谷氨酸棒杆菌精氨酸营养缺陷型菌株的瓜氨酸发酵。
该菌株生产瓜氨酸的精氨酸最佳添加量为0.125mg/L左右。
培养基中的磷盐浓度,对瓜氨酸生成有较大的影响。
所添加的磷盐(磷酸二氢钾和磷酸氢二钾合计)浓度为0.05%时最佳。
如果磷酸钾的浓度是前述浓度数倍时,瓜氨酸生成量下降到原来的一半以下。
例如,在磷盐浓度达0.1%左右时,谷氨酸生成明显增加。
这说明磷盐对这些物质的生物合成的控制具有重要作用。
该精氨酸营养缺陷型菌株,可由10%葡萄糖生成瓜氨酸达10.7g/L,对糖分子收率为11%。
另外,如果在发酵培养基中添加鸟氨酸、谷氨酸或其衍生物(乙酰谷氨酸等),则可以增加瓜氨酸的生成量。
这表明,添加的这类瓜氨酸前体物质,可在发酵中转变为瓜氨酸。
Okumura等由枯草杆菌K选育精氨酸缺陷型突变株,在含有13%葡萄糖的发酵培养基中发酵3天可积累17g/L L-瓜氨酸。
Nakamura等由谷氨酸棒杆菌选育出精氨酸缺陷突变株,在10%葡萄糖的发酵培养基中可积累11g/L的L-瓜氨酸。
2.由营养缺陷型及结构类似物抗性多重突变株发酵生产瓜氨酸Kato等在枯草杆菌的精氨酸及嘧啶双重抗性突变株的基础上,使其再带上L-精氨酸缺陷标记。
该突变株产酸率不受培养基中L-精氨酸浓度的影响,可产L-瓜氨酸26g/L。
为了进一步提高瓜氨酸产量,对黄色短杆菌的精氨酸缺陷突变株赋予磺胺胍抗性,则在11%葡萄糖的发酵培养基中,可生成瓜氨酸30g/L,而在含有3%葡萄糖、0.5%乙酸铵的初始培养基中,流加乙酸和乙酸铵混合液进行培养,则最终可生产瓜氨酸47g/L。
表12-3 L-瓜氨酸产生菌遗传标记及产量菌株遗传标记底物L-瓜氨酸(g·L-1)枯草杆菌谷氨酸小球菌枯草杆菌枯草杆菌黄色短杆菌黄色短杆菌Arg-Arg-Arg-、Arg r、Pyr r6-AU rArg-、SG rArg-、SG r葡萄糖葡萄糖葡萄糖葡萄糖葡萄糖葡萄糖/乙酸铵17112631.83047注:Pyr—嘧啶;6-AU—6-氮杂尿嘧啶;SG—磺胺胍3.酶法生产据报道,可以采用酶法,即采用恶臭假单胞菌之类微生物生产的精氨酸分解酶、L-精氨酸脱亚氨基酶,或采用固定化菌体等,由精氨酸生产瓜氨酸。
第三节精氨酸发酵精氨酸是精氨酸生物合成途径的最终产物。
精氨酸自身是其合成代谢的调节因子,而且精氨酸生物合成途径没有分支,所以精氨酸发酵不能用阻断代谢流,也就是不能用像发酵生产鸟氨酸和瓜氨酸那样,用营养缺陷型来进行。
一、精氨酸发酵的代谢控制育种策略根据精氨酸的生物合成途径及代谢调节机制,L-精氨酸高产菌的育种要点如下:1.解除菌体自身的反馈调节精氨酸的生物合成受精氨酸自身的反馈抑制和反馈阻遏,采用抗反馈调节突变株,以解除精氨酸自身的反馈调节,使精氨酸得以积累。
选育营养缺陷型的回复突变株也可以解除菌体自身的反馈调节,如选育有N-乙酰谷氨酸激酶缺陷的回复突变株。
选育D-精氨酸、精氨酸氧肟酸、2-噻唑丙氨酸(2-TA)、6-氮尿苷、6-巯基嘌呤、8-氮鸟嘌呤、磺胺胍、刀豆氨酸、2-甲基蛋氨酸、6-氮杂尿嘧啶等抗性突变株,均可提高精氨酸的产量。
2.增加前体物的合成由图12-3可以看出,谷氨酸是精氨酸生物合成的前体物,因此,选育氟乙酸、氟柠檬酸、重氮丝氨酸、狭霉素C、德夸菌素、酮基丙二酸、缬氨霉素、寡霉素、对羟基肉桂酸、2,4-二硝基酚、亚砷酸等抗性及氟丙酮酸、脱氢赖氨酸、萘啶酮酸、棕榈酰谷氨酸等敏感突变株,可增加精氨酸前体物的合成,从而有利于精氨酸产量的提高。