真核生物的基因结构
- 格式:doc
- 大小:111.00 KB
- 文档页数:3






真核生物染色体基因组的结构和功能真核生物的基因组一般比较庞大,例如人的单倍体基因组由3×106bp硷基组成,但人细胞中所含基因总数大概会超过3万个。
这就说明在人细胞基因组中有许多DN A序列并不转录成mR NA用于指导蛋白质的合成。
研究发现这些非编码区往往都是一些大量的重复序列,这些重复序列或集中成簇,或分散在基因之间。
在基因内部也有许多能转录但不翻译的间隔序列(内含子)。
因此,在人细胞的整个基因组当中只有很少一部份(约占2-3%)的DNA序列用以编码蛋白质。
真核生物基因组有以下特点。
1.真核生物基因组DNA与蛋白质结合形成染色体,储存于细胞核内,除配子细胞外,体细胞内的基因的基因组是双份的(即双倍体,diploi d),即有两份同源的基因组。
2.真核细胞基因转录产物为单顺反子。
一个结构基因经过转录和翻译生成一个mRNA分子和一条多肽链。
3.存在重复序列,重复次数可达百万次以上。
4.基因组中不编码的区域多于编码区域。
5.大部分基因含有内含子,因此,基因是不连续的。
6.基因组远远大于原核生物的基因组,具有许多复制起点,而每个复制子的长度较小。
高度重复序列:高度重复序列在基因组中重复频率高,可达百万(106)以上。
在基因组中所占比例随种属而异,约占10-60%,在人基因组中约占20%。
高度重复顺序又按其结构特点分为三种(1)反向重复序列这种重复顺序约占人基因组的5%。
反向重复序列由两个相同顺序的互补拷贝在同一DNA链上反向排列而成。
变性后再复性时,同一条链内的互补的拷贝可以形成链内碱基配对,形成发夹式或“+”字形结构。
反向重复间可有一到几个核苷酸的间隔,也可以没有间隔。
没有间隔的又称回文结构,这种结构约占所有反向重复的三分之一。

原核、真核生物基因及表达调控引言现代生物学中“基因”一词甚为流行,细胞学、遗传学、生物化学等,以及各种生物学课本中,都涉及到“基因”一词。
甚至象典型的宏观生物学科——生态学,也把一片森林称为一个“基因库”[1]。
现代生物学已经完全证明,DNA 分子是由称为核普酸的有机分子线性聚合而成。
基因就是核普酸按一定顺序排列而成的DNA分子片段,它携带着遗传信息。
基因表达(gene expression)是指细胞在生命过程中,把储存在DNA顺序中遗传信息经过转录和翻译,转变成具有生物活性的蛋白质分子。
其实质就是遗传信息的转录和翻译。
在个体生长发育过程中,生物遗传信息的表达按一定的时序发生变化(时序调节),并随着内外环境的变化而不断加以修正(环境调控)[2]。
原核生物和真核生物的基因及表达过程有着差异。
随着世界分子生物学研究不断深入,基因表达技术有了很大的提高。
迄今为止,人们已经研究开发出多种原核和真核表达系统用以生产重组蛋白[3]。
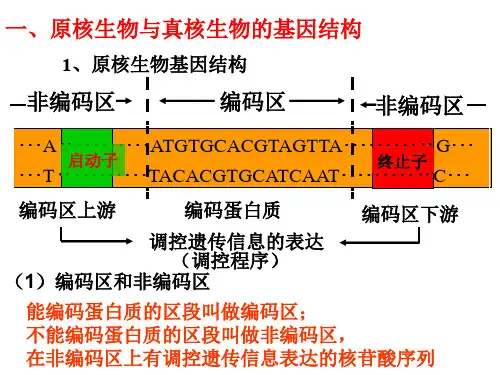
一.原核、真核生物基因结构原核生物基因分为编码区与非编码区,所谓的编码区就是能转录为相应的信使RNA,进而指导蛋白质的合成,非编码区位于编码区的上游及下游。
[4]在调控遗传信息表达的核苷酸序列中最重要的是位于编码区上游的RNA聚合酶结合位点。
RNA聚合酶是催化DNA转录为RNA,能识别调控序列中的结合位点,并与其结合。
真核生物基因结构见图1:图1 真核生物基因结构二.原核、真核生物基因结构的区别最主要的在于真核基因是不连续的,而原核基因是连续的。
所谓真核基因的不连续,即一个基因的编码序列也叫外显子,被一个或多个非编码序列,又叫内含子所间隔。
[5]这些内含子和外显子同属一个转录单位,转录形成前体。
经过转录的加工,即切去内含子,重新连按外显子,从而得到成熟。
而绝大多数的原核基因是连续的,没有内含子的间隔,转录产生成熟。
不仅如此,而且凡在代谢途径上功能有关的多个基因可能紧密相联,与它们的调控基因一起组成一个操纵子,转录到一条链。

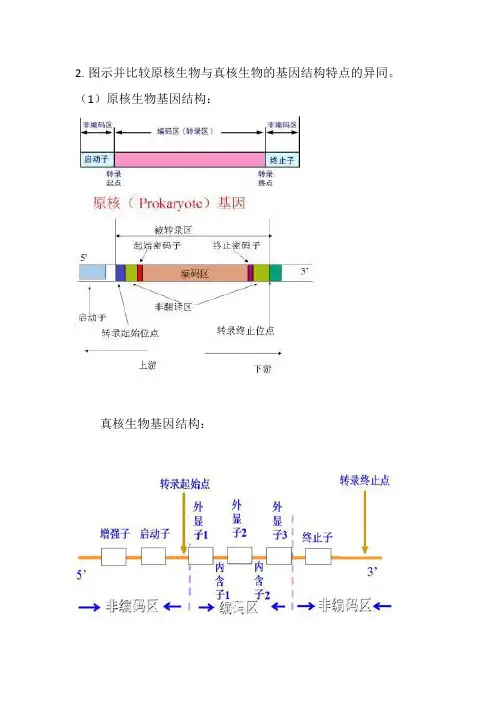
2.图示并比较原核生物与真核生物的基因结构特点的异同。
(1)原核生物基因结构:
真核生物基因结构:
(2)原核生物与真核生物基因结构不同点:
a.原核生物基因编码区是连续的,不断面的。
真核生物
基因编码区是间隔的,不连续的,断面的。
b.原核生物基因编码区不含外显子与内含子,而真核生
物基因编码区则有外显子与内含子。
c.原核生物基因结构较为简单,真核生物基因结构较复
杂且真核生物基因为断裂基因。
d.原核生物基因只有一个复制起点,真核基因为多复制
起点
e.原核生物基因结构含有操纵子,而真核基因一般没有
操纵子
f.原核生物基因为单拷贝基因,而真核基因为多拷贝的。
g.原核生物基因结构一般为环状,而真核基因为链状。
原核生物与真核生物基因结构相同点:
a.均含有编码区和非编码区。
b.均含有转录起点和终点。
c.在非编码区都含有调控遗传信息表达的核苷酸序列,在编码区上游都含有与RNA聚合酶结合的启动子位点。

简述真核生物基因的结构特点真核生物基因的结构特点包括以下几个方面:1. 真核生物基因位于染色体上,是真核生物细胞中的核心结构。
染色体是由 DNA 和蛋白质组成的复合物,是在细胞分裂时传递遗传信息的基本单位。
真核生物基因组的 DNA 与蛋白质结合形成染色体,储存于细胞核内。
除配子细胞外,体细胞内的基因组是双份的 (即双倍体,diploid),即有两份同源的基因组。
2. 真核细胞基因转录产物为单顺反子 (monocistron),即一个结构基因转录、翻译成一个 mRNA 分子,一条多肽链。
真核生物的基因转录是在 DNA 模板上以 RNA 为模板进行转录,产生的 mRNA 是单链,在细胞质中由核糖体(ribosome) 进行翻译。
3. 真核生物基因组中存在大量重复序列,包括高度重复序列和中度重复序列。
高度重复序列重复频率可达 106 次,包括卫星 DNA、反向重复序列和较复杂的重复单位组成的重复序列;中度重复序列可达 103~104 次,如为数众多的Alu 家族序列,KpnI 家族,Hinf 家族序列,以及一些编码区序列如 rRNA 基因、tRNA 基因、组蛋白基因等。
4. 真核生物基因是不连续的,在真核生物结构基因的内部存在许多不编码蛋白质的间隔序列 (interveningsequences),称为内含子 (intron),编码区则称为外显子 (exon)。
内含子与外显子相间排列,转录时一起被转录下来,然后RNA 中的内含子被切掉,外显子连接在一起成为成熟的 mRNA,作为指导蛋白质合成的模板。
5. 真核生物基因组远大于原核生物的基因组,具有许多复制起点,而每个复制子的长度较小。
真核生物基因组 DNA 与蛋白质结合形成染色体,储存于细胞核内。
除配子细胞外,体细胞内的基因组是双份的 (即双倍体,diploid),即有两份同源的基因组。
真核生物的基因结构
真核生物的基因结构包括编码区和非编码区。
编码区其实是断裂基因结构,也就是不连续基因。
具有蛋白编码功能的不连续DNA 序列称为外显子,
外显子之间的非编码序列为内含子。
每个外显子和内含子接头区都有一段高度保守的一致序列,即内含子5'末端大多数是GT 开始,3'末端大多是AG 结束,称为GT-AG 法则,是普遍存在于真核基因中RNA 剪接的识别信号。
第一个外显子首端和最后一个外显子末端,分别为翻译蛋白的起始密码子和终止密码子。
============================================================= 首位和末位外显子两侧的区域为非编码区,也可以叫做侧翼序列,侧翼序列中包含一些调控元件,比如启动子、终止子,还可能有增强子。
上游侧翼序列包含启动子区域,启动子区域包含:
5’端TSS 上游约20~30个核苷酸的位置,有TATA 框(TATA box),碱基序
列为TATAATAAT,是RNA聚合酶的重要的接触点,它能够使酶准确地识别转录的起始点并开始转录,影响着转录开始的位点。
真核生物结构基因真核生物是指细胞内有核和其他细胞器的生物,包括动物、植物、真菌和原生动植物等。
结构基因是指编码蛋白质的基因。
在真核生物中,结构基因是生命活动的重要组成部分,控制着细胞的生长和分化等重要生物过程。
真核生物的细胞结构真核生物的细胞结构主要包括细胞核、线粒体、内质网、高尔基体、溶酶体等。
其中,细胞核是真核细胞的重要特征,包含了细胞的遗传信息和控制蛋白质合成的核糖核酸。
线粒体是细胞内的能量中心,参与了细胞的呼吸作用。
内质网则参与了蛋白质的合成和运输等活动。
高尔基体则负责将蛋白质及其他分子送达其它细胞器或细胞外。
溶酶体则起到了消化细胞外物质和细胞内垃圾的作用。
真核生物的基因结构真核生物的基因结构主要包括启动子、外显子和内含子等。
启动子是基因的起始点,其中包含有调控基因转录的序列。
外显子则编码了蛋白质所需要的信息。
内含子则是指在基因转录过程中,需要被剪切掉的无用序列。
另外,真核生物的基因还包括了转录因子和RNA剪接因子等,这些因子参与了基因的表达和调控过程。
真核生物的基因表达调控真核生物的基因表达调控非常复杂,包括转录起始、RNA加工、转运和翻译等多个环节。
其中,转录因子的作用非常重要,它们可以结合到启动子上,激活或抑制基因的转录过程。
此外,RNA剪接也是基因表达的重要调控机制,它可以选择性地剪切内含子,从而调节外显子的组合方式,影响蛋白质的翻译过程。
真核生物的基因编辑技术近年来,CRISPR/Cas9等基因编辑技术的出现,使得真核生物的基因编辑变得更加方便和高效。
通过这些技术,可以对真核生物的基因进行精准的编辑和修改,从而实现对基因表达和调控的精确控制。
这些技术的出现为真核生物研究提供了新的思路和手段,也为人类疾病的治疗和基因工程的发展提供了新的可能性。
真核生物结构基因是生命活动的重要组成部分,对于真核生物的生长、分化和遗传等方面都有着重要的作用。
随着基因编辑技术的不断发展,相信我们对于真核生物结构基因的了解和应用将会更加深入和广泛。
真核生物的基因结构
真核生物的基因结构包括编码区和非编码区。
编码区其实是断裂基因结构,也就是不连续基因。
具有蛋白编码功能的不连续DNA 序列称为外显子,
外显子之间的非编码序列为内含子。
每个外显子和内含子接头区都有一段高度保守的一致序列,即内含子5’末端大多数是GT 开始,3’末端大多是AG 结束,称为GT-AG 法则,是普遍存在于真核基因中RNA 剪接的识别信号。
第一个外显子首端和最后一个外显子末端,分别为翻译蛋白的起始密码子和终止密码子。
====================================================== =======
首位和末位外显子两侧的区域为非编码区,也可以叫做侧翼序列,侧翼序列中包含一些调控元件,比如启动子、终止子,还可能有增强子。
上游侧翼序列包含启动子区域,启动子区域包含:
列为TATAATAAT,是RNA聚合酶的重要的接触点,它能够使酶准确地识别转录
止密码子之间区域,不编码蛋白质。
miRNA 经常结合于3‘UTR,从而引起mRNA。