条件性基因敲除与敲入
- 格式:pptx
- 大小:476.57 KB
- 文档页数:38

传统ES打靶基因敲除敲入小鼠技能传统ES 打靶基因敲除/敲入小鼠技术技术原理传统的基因打靶技术制备基因敲除(KO )/敲入(KI )基因打靶技术是建立在DNA同源重组与胚胎干细胞等技术基础上的分子生物学技术。
同源重组是指当外源DNA 片段与宿主基因组片段同源性高时,同源DNA 区部分可与宿主DNA 的相应片段发生交换(即同源重组)。
基因打靶就是通过同源重组技术将外源基因定点整合入靶细胞基因组上某一确定的位点,以达到定点修饰改造染色体上某一基因的目的。
基因打靶技术目前已被广泛认为是一种理想的特定修饰与改造生物体遗传物质的最佳方法。
尤其是条件性和诱导性基因打靶系统的建立,使得对基因在时间和空间上的靶位修饰更加明确、效果更加精确可靠,该技术的发展已经为发育生物学、分子遗传学、免疫学及医学等学科提供了一个全新的、强有力的研究和治疗手段,并已显示出巨大的应用前景及商业价值。
服务流程和周期、管路敷设技术通过管线不仅可以解决吊顶层配置不规范高中资料试卷问题,而且可保障各类管路习题到位。
在管路敷设过程中,要加强看护关于管路高中资料试卷连接管口处理高中资料试卷弯扁度固定盒位置保护层防腐跨接地线弯曲半径标高等,要求技术交底。
管线敷设技术中包含线槽、管架等多项方式,为解决高中语文电气课件中管壁薄、接口不严等问题,合理利用管线敷设技术。
线缆敷设原则:在分线盒处,当不同电压回路交叉时,应采用金属隔板进行隔开处理;同一线槽内,强电回路须同时切断习题电源,线缆敷设完毕,要进行检查和检测处理。
、电气课件中调试对全部高中资料试卷电气设备,在安装过程中以及安装结束后进行高中资料试卷调整试验;通电检查所有设备高中资料试卷相互作用与相互关系,根据生产工艺高中资料试卷要求,对电气设备进行空载与带负荷下高中资料试卷调控试验;对设备进行调整使其在正常工况下与过度工作下都可以正常工作;对于继电保护进行整核对定值,审核与校对图纸,编写复杂设备与装置高中资料试卷调试方案,编写重要设备高中资料试卷试验方案以及系统启动方案;对整套启动过程中高中资料试卷电气设备进行调试工作并且进行过关运行高中资料试卷技术指导。

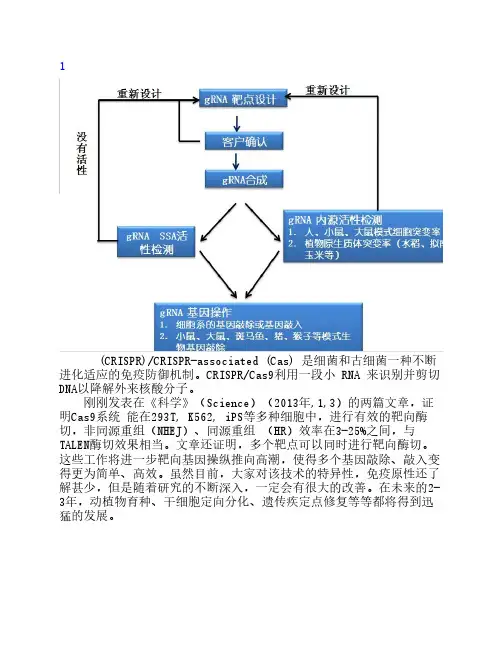
1(CRISPR)/CRISPR-associated (Cas) 是细菌和古细菌一种不断进化适应的免疫防御机制。
CRISPR/Cas9利用一段小 RNA 来识别并剪切DNA以降解外来核酸分子。
刚刚发表在《科学》(Science)(2013年,1,3)的两篇文章,证明Cas9系统 能在293T, K562, iPS等多种细胞中,进行有效的靶向酶切,非同源重组(NHEJ)、同源重组 (HR)效率在3-25%之间,与TALEN酶切效果相当。
文章还证明,多个靶点可以同时进行靶向酶切。
这些工作将进一步靶向基因操纵推向高潮,使得多个基因敲除、敲入变得更为简单、高效。
虽然目前,大家对该技术的特异性,免疫原性还了解甚少,但是随着研究的不断深入,一定会有很大的改善。
在未来的2-3年,动植物育种、干细胞定向分化、遗传疾定点修复等等都将得到迅猛的发展。
图1. RNA指导的CRISPR/Cas9基因剪切系统唯尚立德已经利用CRISPR/Cas9在斑马鱼,哺乳动物细胞上成功实现基因敲除和基因敲入,现推出如下技术服务:1. 应用CRISPR /Cas9提供稳定细胞系靶向基因敲除、敲入技术服务;2. 应用CRISPR /Cas9提供模式生物靶向基因技术服务,包括小鼠,大鼠,斑马鱼等;近几十年来,随着全基因组测序技术的不断成熟,我们在各种细菌和古细菌(archaea)中也陆续发现了很多成簇的、规律间隔的短回文重复序列(clustered regularly interspaced short palindromic repeat sequences,即CRISPR序列,这就是二十多年前日本科学家发现的那个序列)和CRISPR相关基因(CRISPR-associated genes, Cas gene)。
研究发现,这些CRISPR序列与很多病毒或者质粒的DNA序列是互补的,说明这套CRISPR–Cas系统很有可能是生物体抵御病毒等外来入侵者的一套特异性防御机制,就好像是另外一套适应性免疫反应系统(adaptive immune system)。

基因打靶基因打靶包括:胚胎干细胞的获得和培养、打靶载体的构建、重组ES细胞的筛选、嵌合体小鼠的制备、基因敲除小鼠的建立、Cre-loxP系统、FLP/FRT系统和条件性基因敲除、基因敲入和大规模ES细胞突变库的建立。
基本概念:1.基因打靶:是利用同源重组技术来定点改变物种的基因组顺序和结构,从而在突变的个体内来研究基因及基因组的功能。
2.基因敲除:是使用基因组中某个/某几个基因或基因的顺式元件产生缺陷,从而在突变体内。
3.丧生正常的功能,来推测这些基因或元件原来在体内的功能。
基因敲入:在个体基因组中定点加入某个/某几个基因或顺式元件,使之表达或发挥作用,从而研究该基因或顺式元件在体内的功能。
4.基因打靶技术是一种定向改变生物活体遗传信息的实验手段。
它的产生和发展建立在胚胎干细胞技术和同源重组技术成就的基础之上,并促进了相关技术的进一步发展。
自1987年早期胚胎干细胞技术建立及第一例基因剔除小鼠诞生以来,基因打靶的研究进展迅速,给现代生物学和医学研究带来了革命性的变化,并直接引发了现代生物学和医学研究各个领域中许多突破性的进展,成为后基因组时代研究基因功能最直接和最有效的方法之一。
一、胚胎干细胞的获得和培养基因打靶中用的小鼠ES细胞系有:D3、E14、R1、J1、CCE,均来源于129小鼠品系和其杂交品系(因为这类小鼠具有自发突变形成畸胎瘤和畸胎肉瘤的倾向,是基因敲除的理想实验动物)。
ES 623和B6-IIIES细胞系,来源于C57BL/6小鼠品系。
BALB/c-I,来源于BALB/c小鼠品系。
常用的饲养层细胞为PMEF(小鼠原代胚成纤维细胞。
PMEF需6 Gy的X 射线照射或丝裂霉素C处理细胞抑制生长后才能用作饲养细胞)。
建立ES细胞的过程中,最好采用只传了2-3代的原代小鼠胚胎成纤维细胞作为饲养细胞,所取得的ICM(内细胞团)只有10%-30%的几率建立ES细胞系。
一旦ES 克隆被确定,接下来应该考虑检查ES细胞的核型。

四环素诱导基因敲除敲入技术服务详解Cre重组酶是一种位点特异性重组酶,能介导两个LoxP位点(序列)之间的特异性重组,使LoxP位点间的基因序列被删除或重组。
LoxP序列是由两个13bp反向重复序列和中间间隔的8bp序列共同组成,8bp的间隔序列同时也确定了LoxP的方向。
Cre在催化DNA 链交换过程中与DNA共价结合,13bp的反向重复序列是Cre酶的结合域。
Cre重组酶介导两个LoxP位点间的重组是一个动态、可逆的过程,可以分成三种情况:1.如果两个LoxP位点位于一条DNA链上,且方向相同,Cre重组酶能有效切除两个LoxP位点间的序列;2.如果两个LoxP位点位于一条DNA链上,但方向相反,Cre重组酶能导致两个LoxP位点间的序列倒位;3.如果两个LoxP位点分别位于两条不同的DNA链或染色体上,Cre酶能介导两条DNA链的交换或染色体易位。
另外,Cre不仅可以识别LoxP的2个13bp的反向重复序列和8bp的间隔区域,而且当一个13bp的反向重复序列或者8bp的间隔区发生改变时仍能识别并发生重组。
利用这一特点,人们在构建载体时可以根据需要改造LoxP位点序列,以用于特定的基因突变或修复,增加了该系统的应用范围。
诱导性基因敲除以Cre/loxp系统为基础,但却是利用控制Cre表达的启动子的活性或所表达的Cre酶活性具有可诱导的特点,通过对诱导剂给予时间的控制或利用Cre基因定位表达系统中载体的宿主细胞特异性和将该表达系统转移到动物体内的过程在时间上的可控性,从而在loxP动物的一定发育阶段和一定组织细胞中实现对特定基因进行遗传修饰之目的的基因敲除技术。
常见的几种诱导性类型如下:四环素诱导型;干扰素诱导型;激素诱导型;腺病毒介导型。
Cre-LoxP图:该系统包含两个互补系统,分别为tTA依赖和rtTA依赖的基因敲除系统,现在又被称为Tet―Off (tTA依赖)系统和T et―On (rtTA依赖)系统。

转基因、基因敲入/敲除动物技术已经成为现代生命科学基础研究和药物研发领域不可或缺的重要技术,该技术从上世纪七八十年代诞生以来,已有近四十年的历史,经典技术如DNA原核显微注射、胚胎干细胞显微注射技术一直以来经久不衰,并逐渐从基础研究实验室转向商业模式,成为一项高度标准化的新兴产业一、技术介绍与研究进展转基因、基因敲入/敲除动物技术已经成为现代生命科学基础研究和药物研发领域不可或缺的重要技术,该技术从上世纪七八十年代诞生以来,至今已有近四十年的历史,经典技术如DNA原核显微注射、胚胎干细胞显微注射技术一直以来经久不衰,在小鼠模型构建方面日趋完善,并且如同剪切酶和抗体等常规分子生物学试剂的制备技术一样,逐渐从基础研究实验室转向商业模式,成为一项高度标准化的新兴产业,催生了数以百计的创新药物和数以千计的优秀文章。
尽管如此,传统技术仍然存在一些难以克服的缺陷,如步骤繁琐、周期漫长、成功率低、费用高昂等,而ZFN和TALEN等新技术的出现,或有可能将这一局面彻底改变。
二、同源重组技术原理基因敲除鼠技术是上世纪80年代中后期基于DNA同源重组的原理发展起来的,Capecchi和Smithies在1987年根据同源重组(homologous recombination)的原理,首次实现了ES的外源基因的定点整合(targeted integration),这一技术称为"基因打靶"(gene targeting)或"基因敲除"(gene knockout),利用这种ES的显微注射就可以制作出基因敲出小鼠(KO Mice: knockout mice);由于这一工作,Capecchi和Smithies于2007年与Evans分享了诺贝尔医学奖。
同源重组(homologous recombination)定义:是指发生在姐妹染色单体(sister chromatin) 之间或同一染色体上含有同源序列的DNA分子之间或分子之内的重新组合。

条件性基因敲除的基本原理Cre/loxP重组系统条件性基因敲除的基本原理 Cre / loxP 重组系统条件性基因敲除主要是通过Cre/10xP或者Ftp/FRT重组系统来实现的。
这两个系统都是位点特异性重组酶系统,已发展成为在体内、外进行遗传操作的有力工具。
这两个系统的应用,可以使靶基因的表达或缺失发生在试验动物发育的某一阶段或某一特定的组织器官。
此外,若与控制Cre或Flp表达的其他诱导系统相结合,还可以对某一基因同时实现时空两方面的调控。
1.Cre/loxP系统的原理Cre/loxP系统来源于F1噬菌体,可以介导位点特异的DNA重组。
该系统含有两种成分:①一段长34bp的DNA序列,含有两个13 bp的反向重复序列和一个8 bp的核心序列。
这段34bp序列是重组酶识别的位点,被称为loxP位点(10cus of X―over in P1)。
②Cre重组酶(cyclizationrecombination),它是一种由343个氨基酸组成的单体蛋白,可以引发loxP位点的DNA重组。
任何序列的DNA,当其位于两个loxP位点之间的时候,在Cre重组酶的作用下要么被缺失(两个loxP位点的方向相同),要么方向发生倒转(两loxP位点的方向相反),如图所示。
Cre/loxP系统的作用机制2.Cre/loxP系统优点Cre/10xP系统之所以在基因敲除中获得了非常广泛的应用,是由该系统的诸多优点决定的:①Cre重组酶与具有loxP位点的DNA片断形成复合物后,可以提供足够的能量引发之后的DNA重组过程,因此该系统不需要细胞或者生物体提供其他的辅助因子;②loxP位点是一段较短的DNA序列,因此非常容易合成;③Cre重组酶是一种比较稳定的蛋白质,因此可以在生物体不同的组织、不同的生理条件下发挥作用;④Cre重组酶的编码基因可以置于任何一种启动子的调控之下,从而使这种重组酶在生物体不同的细胞、组织、器官,以及不同的发育阶段或不同的生理条件下产生,进而发挥作用,这一点也是该系统在应用过程中最为重要的一点。

.概述:基因敲除是自80年代末以来发展起来的一种新型分子生物学技术,是通过一定的途径使机体特定的基因失活或缺失的技术。
通常意义上的基因敲除主要是应用DNA同源重组原理,用设计的同源片段替代靶基因片段,从而达到基因敲除的目的。
随着基因敲除技术的发展,除了同源重组外,新的原理和技术也逐渐被应用,比较成功的有基因的插入突变和iRNA,它们同样可以达到基因敲除的目的。
2.实现基因敲除的多种原理和方法:2.1.利用基因同源重组进行基因敲除基因敲除是80年代后半期应用DNA同源重组原理发展起来的。
80年代初,胚胎干细胞<ES细胞)分离和体外培养的成功奠定了基因敲除的技术基础。
1985年,首次证实的哺乳动物细胞中同源重组的存在奠定了基因敲除的理论基础。
到1987年,Thompsson首次建立了完整的ES细胞基因敲除的小鼠模型[1]。
直到现在,运用基因同源重组进行基因敲除依然是构建基因敲除动物模型中最普遍的使用方法。
2.1.1利用同源重组构建基因敲除动物模型的基本步骤(图1>:a.基因载体的构建:把目的基因和与细胞内靶基因特异片段同源的DNA 分子都重组到带有标记基因(如neo 基因,TK 基因等>的载体上,成为重组载体。
基因敲除是为了使某一基因失去其生理功能,所以一般设计为替换型载体。
b.ES 细胞的获得:现在基因敲除一般采用是胚胎干细胞,最常用的是鼠,而兔,猪,鸡等的胚胎干细胞也有使用。
常用的鼠的种系是129及其杂合体,因为这类小鼠具有自发突变形成畸胎瘤和畸胎肉瘤的倾向,是基因敲除的理想实验动物。
而其他遗传背景的胚胎干细胞系也逐渐被发展应用。
[2,3] c.同源重组:将重组载体通过一定的方式(电穿孔法或显微注射>导入同源的胚胎干细胞(ES cell>中,使外源DNA与胚胎干细胞基因组中相应部分发生同源重组,将重组载体中的DNA序列整合到内源基因组中,从而得以表达。
一般地,显微注射命中率较高,但技术难度较大,电穿孔命中率比显微注射低,但便于使用。

基因敲除技术(组图)一.概述:基因敲除是自80年代末以来发展起来的一种新型分子生物学技术,是通过一定的途径使机体特定的基因失活或缺失的技术。
通常意义上的基因敲除主要是应用DNA 同源重组原理,用设计的同源片段替代靶基因片段,从而达到基因敲除的目的。
随着基因敲除技术的发展,除了同源重组外,新的原理和技术也逐渐被应用,比较成功的有基因的插入突变和iRNA ,它们同样可以达到基因敲除的目的。
二.实现基因敲除的多种原理和方法:1.利用基因同源重组进行基因敲除基因敲除是80年代后半期应用DNA 同源重组原理发展起来的。
80年代初,胚胎干细胞(ES细胞)分离和体外培养的成功奠定了基因敲除的技术基础。
1985年,首次证实的哺乳动物细胞中同源重组的存在奠定了基因敲除的理论基础。
到1987年,Thompsson首次建立了完整的ES细胞基因敲除的小鼠模型[1]。
直到现在,运用基因同源重组进行基因敲除依然是构建基因敲除动物模型中最普遍的使用方法。
(1)利用同源重组构建基因敲除动物模型的基本步骤(图1):①.基因载体的构建:把目的基因和与细胞内靶基因特异片段同源的DNA 分子都重组到带有标记基因(如neo 基因,TK 基因等)的载体上,成为重组载体。
基因敲除是为了使某一基因失去其生理功能,所以一般设计为替换型载体。
②.ES 细胞的获得:现在基因敲除一般采用是胚胎干细胞,最常用的是鼠,而兔,猪,鸡等的胚胎干细胞也有使用。
常用的鼠的种系是129及其杂合体,因为这类小鼠具有自发突变形成畸胎瘤和畸胎肉瘤的倾向,是基因敲除的理想实验动物。
而其他遗传背景的胚胎干细胞系也逐渐被发展应用。
[2,3]③.同源重组:将重组载体通过一定的方式(电穿孔法或显微注射)导入同源的胚胎干细胞(ES cell)中,使外源DNA 与胚胎干细胞基因组中相应部分发生同源重组,将重组载体中的DNA 序列整合到内源基因组中,从而得以表达。
一般地,显微注射命中率较高,但技术难度较大,电穿孔命中率比显微注射低,但便于使用。

如何正确选择基因敲除敲⼊质粒?研究基因功能最有效的⽅法之⼀是⽤实验室设计的DNA⽚段取代或破坏基因,使其失活或“敲除”。
构建特殊的质粒可以通过同源重组替代酵母、⼩⿏或果蝇中的基因。
其原理很简单:将⼀个带有⽬标序列修复模板的质粒递送到细胞,该细胞将模板与内源性基因重组。
在这⾥,我们将主要探讨⽤于灭活哺乳动物细胞中特定基因的技术和质粒。
尽管CRISPR的敲除/敲⼊系统⾮常流⾏,但这个系统仍然很有价值,特别是在CRISPR不能使⽤的情况下(例如,附近没有合适的PAM序列,或者你感兴趣的基因很难⽤gRNA特异性靶向)。
在选择敲除策略时,⼀定要记住这些技巧!敲除质粒1.同源重组是⼀种准确修复破坏性的双链断裂的机制,核苷酸序列在两个相似或相同的DNA分⼦之间进⾏交换。
基因靶向则是利⽤这⼀⾃然过程,运⽤⼀种特殊设计的含有同源序列的载体,将⽬标基因位点替换为同源序列。
为了更了解这个过程,我们将介绍⼀个旨在敲除给定基因的第2外显⼦的实验。
设计你的⽬标结构。
为了在细胞中重组,⾄少需要2 kb的同源序列,但6到14 kb的同源序列是靶向结构的典型特征。
在图1所⽰的例⼦中,与⽬标基因外显⼦1和外显⼦3相对应的序列已克隆到载体抗⽣素抗性基因的任意⼀侧。
为了避免选择那些结构随机整合到基因组中的细胞,像HSV 胸苷激酶(HSV-tk)这样的阴性选择标记被包含在⼀个同源臂之外。
当我们稍后选择细胞时,我们将⾸先对抗⽣素抗性基因进⾏阳性选择,然后对阴性选择标记进⾏反选择——这后⼀步将杀死许多随机整合了全部或⼤部分质粒的细胞。
2.呈递质粒给宿主细胞。
重组后,⽬标基因的第2外显⼦将从染⾊体上移除,并被抗性基因取代。
这样基因就被破坏或敲除了。
3.利⽤正负选择确定正确的重组细胞。
重组是⼀个偶然事件,所以你必须选择⽽不是筛选已经发⽣重组的细胞。
新霉素、嘌呤霉素和潮霉素耐药基因通常⽤于阳性选择。
正选择标记对重组进⾏选择,⽽负选择标记对不适当的、随机的到不同位点的重组进⾏选择。

基因敲除⼩⿏饲养繁殖技巧科研狗但凡要做动物实验的,哪个不需要伺候⼩⿏?每天好吃的好喝地伺候着,定期更换垫料,还要操⼼它们的⼈⽣⼤事,给它分配对象,⽣了娃还要给帮它们做“亲⼦鉴定”,看见⼩⿏脱发了,要想办法,母⿏不⽣了,要想办法。
回头再看看⾃⼰,没头发,还没对象,天天愁⽂章,担⼼毕不了业,全指望⼩⿏们加油,让⾃⼰早⽇出成果,所以可不得好吃好喝的伺候它们嘛!尤其是基因敲除⼩⿏,⾝价昂贵,从接到它的那⼀刻起,整个⼈都⼜兴奋⼜紧张,⽣怕怠慢了这些⼩家伙,那么收到基因敲除⼩⿏后要如何制定计划进⾏扩群繁殖以满⾜实验需要呢?除了了解所选品系的⽣活习性外,我们还要考虑⾃⼰需要的基因编辑⿏可能出现哪些问题,有没有解决或者是改善的⽅法,这些我们可以通过查阅⽂献看有没有关于该基因敲除⼩⿏的相关信息并作出⼀个初步判断。
1.背景品系的特点这个不⽤多说,需要做动物实验的同学,谁还没养过⼏只⼩⿏呀,养之前肯定已经把它的⽣活习性、繁殖特点、饲养要求等了解的很清楚了,举个栗⼦,⽐如说C57BL/6J,该品系的雄⿏好⽃,尤其是刚断奶后不同窝的雄⿏不宜放⼀笼饲养。
2.饲养环境的要求应尽可能地给⼩⿏提供适宜的⽣活环境和营养,这⽅⾯可以参照国家标准来进⾏饲养管理。
⽐如,⼩⿏对于噪⾳、光照⽐较敏感,所以我们应给⼩⿏提供安静的环境,尽量选在⼈员⾛动少的地⽅,也不要频繁查看⼩⿏情况,这样⼩⿏会有压⼒的。
母⿏有筑巢的天性,可以向笼中放柔软的棉垫,⿎励其筑巢,有助于提⾼产量。
3.基因敲除⼩⿏饲养管理应注意哪些问题⼤多数基因敲除⼩⿏相对野⽣型⼩⿏⽽⾔,在某些⽅⾯会存在或⼤或⼩的问题,所以我们需要查阅相关的⽂献,看看有没有该基因敲除⼩⿏的⽂献报道,该基因被修饰后会出现哪些问题?这些问题有没有解决或者是改善的⽅法。
如果是某种基因被修饰后会导致⼩⿏不能合成某种物质,那么可以给⼩⿏额外补充该物质;若是基因敲除⼩⿏的成活率低,问题有可能出现在着床期⾄成年期的任意时间段,通过代乳和改善饲养条件也许能得以解决;如果基因敲除⼩⿏的⽣殖⼒低。
基因敲除基因敲除(knockout)是指一种遗传工程技术,针对某个序列已知但功能未知的序列,改变生物的遗传基因,令特定的基因功能丧失作用,从而使部分功能被屏蔽,并可进一步对生物体造成影响,进而推测出该基因的生物学功能。
指外源DNA与受体细胞基因组中序列相同或相近的基因发生同源重组,从而代替受体细胞基因组中的相同/相似的基因序列,整合入受体细胞的基因组中。
此法可产生精确的基因突变,也可正确纠正机体的基因突变。
基因嵌入又称基因置换,它是利用内源基因序列两侧或外面的断裂点,用同源序列的目的基因整个置换内源基因。
用于基因敲除和基因嵌入的技术有Cre/Lox P系统、FLPI系统等。
步骤:获得干细胞基因敲除一般应用于鼠,而最常用的鼠的种系是129及其杂合体,因为这类小鼠具有自发突变形成畸胎瘤和畸胎肉瘤的倾向,是基因敲除的理想实验动物。
而其他遗传背景的胚胎干细胞系逐渐被发展应用,最来自于C57BL/6×CBN/JNCrjF1小鼠的胚胎干细胞系成功地用于基因敲除。
由于这些远交系遗传背景复杂,所得到的模式小鼠往往不能得到重复性好的实验结果,所以也需要在C57BL/6等近交系小鼠上做回交。
另一方面,因为回交次数不一样,也会造成实验结果重复性差。
这对生物科研,尤其是医药企业,安评中心,药检部门等,是一个很大的缺点。
这对开发制作标准模式动物也是一个很大的缺陷。
所以,如果能够直接用C57BL/6 ES细胞进行基因打靶,就将直接获得C57BL/6品系的模式小鼠。
c57BL/6小鼠种系等已经广泛的应用于免疫学,神经学,癌症,等几乎所有研究领域。
已经有一些公司或科研机构已经开始用C57BL/6遗传背景的胚胎干细胞进行基因打靶。
载体构建把目的基因和与细胞内靶基因特异片段同源的DNA分子都重组到带有标记基因(如neo基因,TK基因等)的载体上,此重组载体即为打靶载体。
因基因打靶的目的不同,此载体有不同的设计方法,可分为替换性载体和插入型载体。
敲除小鼠常见问题与回答一、什么是ES细胞显微注射?答:胚胎干细胞显微注射是制作基因敲除小鼠的一个最常用的方法。
主要过程是将携带目的基因的胚胎干细胞注射到小鼠的囊胚腔中获得嵌合体小鼠。
所得嵌合体小鼠的组织,将同时含有来源于囊胚的细胞和胚胎干细胞。
嵌合体小鼠必须和野生型小鼠配种以决定遗传改变的生殖系能否传递,这样可能获得转基因或打靶基因(来源于胚胎干细胞)稳定的生殖系传递的小鼠。
一般情况下,大约能获得50%继承了目的基因的后代。
二、嵌合体遗传学上用以指不同遗传性状嵌合或混杂表现的个体。
免疫学上的涵义则指一个机体身上有两种或两种以上染色体组成不同的细胞系同时存在,彼此能够耐受,不产生排斥反应,相互间处在嵌合状态。
在基因敲除鼠中指通过向囊胚注射被外源基因转化了的胚胎干细胞,使得发育成为的个体中含有不同基因型的细胞,产生的个体也叫嵌合体,即该生物体中嵌合了两种不同遗传结构的细胞(一种是基因型被改变了的细胞,另一种是原来的基因型的细胞)。
三、条件性敲除的原理?答:Cre-LoxP系统是源于P1噬菌体的一个DNA重组体系,由Cre酶和相应的LoxP位点组成,它能导致重组发生在特定的DNA序列处(LoxP位点),该系统可以将外源基因定点整合到染色体上或将特定DNA片段删除。
基于Cre-LoxP的基因打靶要分两步来进行。
首先要在胚胎干细胞的基因组中引入LoxP序列,这一步可以通过打靶载体的设计和对同源重组子的筛选来实现。
下一步通过Cre介导的重组来实现靶基因的遗传修饰或改变。
Cre-LoxP系统既可以在细胞水平上用Cre重组酶表达质粒转染中靶细胞,通过识别LoxP位点将抗性标记基因切除,又可以在个体水平上将重组杂合子小鼠与Cre转基因小鼠杂交,筛选子代小鼠就可得到删除外源标记基因的条件性敲除小鼠。
四、如何鉴定和挑选嵌合体?答:动物只有部分组织细胞整合有外源基因,则称为嵌合体动物。
它的鉴定主要根据毛色去鉴定。
注射的ES和囊胚来源不同的小鼠品系。
基因敲除和敲入技术的发展和应用随着科技的不断进步,人类对于基因的探究也日渐深入。
基因敲除和敲入技术作为其中的重要探究手段,则受到越来越多的关注和应用。
本文将从技术原理、研究进展、应用前景等方面阐述基因敲除和敲入技术。
一、技术原理基因敲除和敲入技术是利用DNA重组技术将人造的基因片段有选择地插入到细胞或某个生物体的特定位置,从而改变这个生物体的某种性状或者产生新的性状。
所谓基因敲除,就是将某个基因的序列刻意人为删除;而基因敲入,则是将人造基因片段有选择地插入到细胞或生物体中。
这两种技术都属于基因工程领域内的基础技术,也为其他生命科学领域提供了良好的研究工具。
二、研究进展基因敲除和敲入技术的研究始于20世纪80年代,最早是借助于整合子(Transposon)等修饰体来达到基因敲入的目的。
然而其限制因素太多,因此研究人员开始寻找其他方法。
1996年,Nobuyoshi Shimizu和John Witthuhn在细菌中利用锂盐法将基因敲除策略应用到实践中,为基因敲除技术的研发打下了基础。
此后的20多年里,随着不断的技术进步和生物技术产业的蓬勃发展,基因敲除和敲入技术也得到了飞速发展。
最近几年,基因敲除和敲入技术的进步尤其引人注目。
CRISPR-Cas9技术以其高效易用和精确性受到广泛关注,成为现在最主流的基因敲除和敲入技术之一。
除此之外,ZFN(zinc finger nuclease)和TALEN(TAL effector nuclease)等技术也在持续发展中,不断完善。
随着技术的不断创新和完善,基因敲除和敲入技术的应用范围也越来越广泛。
三、应用前景基因敲除和敲入技术得到广泛应用的趋势也日益明显,涵盖了从基础科学到临床医学的多个应用领域。
在基础科学领域,基因敲除和敲入技术可以被广泛应用于基础遗传学、生理学、细胞生物学、分子生物学等各个领域的研究。
以生物缺陷基因的研究为例,利用这两种技术可以实现对缺陷基因的破坏或修复,有助于进一步认识缺陷基因对生命活动的影响机制等。
生物节律基因检测方法生物节律是生物体内部生理和行为上的周期性变化,这些变化由基因控制,并与地球的昼夜节律密切相关。
随着科学技术的发展,生物节律基因检测方法已逐渐成为研究的热点。
本文将为您详细介绍生物节律基因检测的相关方法。
一、概述生物节律基因检测旨在揭示生物体内部节律性变化的遗传机制,为生物节律调控、疾病预防与治疗提供科学依据。
目前,研究人员已开发出多种生物节律基因检测方法,主要包括以下几种:1.基因敲除与基因敲入技术2.基因表达谱分析3.基因突变检测4.蛋白质组学分析5.生物信息学方法二、具体检测方法1.基因敲除与基因敲入技术基因敲除技术通过破坏目标基因,观察生物体节律性变化,从而揭示基因在生物节律调控中的作用。
基因敲入技术则是在生物体内引入外源基因,研究其功能。
这两种技术均可用于生物节律基因的研究。
2.基因表达谱分析基因表达谱分析是通过检测生物体在不同时间点的基因表达水平,分析基因表达与生物节律的关系。
这种方法的优点是能同时研究多个基因的表达变化,有助于揭示生物节律调控网络的复杂性。
3.基因突变检测基因突变检测是针对已知生物节律基因的突变进行检测,分析突变对生物节律的影响。
这种方法有助于发现新的生物节律基因及突变类型。
4.蛋白质组学分析蛋白质组学分析是通过研究生物体在不同时间点的蛋白质表达变化,揭示生物节律调控的分子机制。
这种方法的优点是能直接反映蛋白质水平的变化,有助于发现新的生物节律调控因子。
5.生物信息学方法生物信息学方法是通过构建生物节律基因调控网络,预测生物节律基因的功能和相互作用。
这种方法可以在大规模基因组数据中挖掘生物节律基因,为实验研究提供线索。
三、总结生物节律基因检测方法的研究为揭示生物节律的遗传机制提供了有力手段。
随着科学技术的不断发展,更多高效、准确的检测方法将被开发出来,为生物节律调控及相关疾病的研究提供有力支持。