线粒体
- 格式:doc
- 大小:19.02 KB
- 文档页数:6

线粒体第一节形态结构一、线粒体的形状、大小、数目和分布♦形状多样,多为卵圆形或棒状。
♦大小直径0.5-1.0μm,长1.5-3.0μm。
♦数目不同细胞内差异大,通常含1000-2000个,新陈代谢旺盛的细胞内含量较多。
♦分布多分布于生理功能旺盛的区域和需要能量较多的部位。
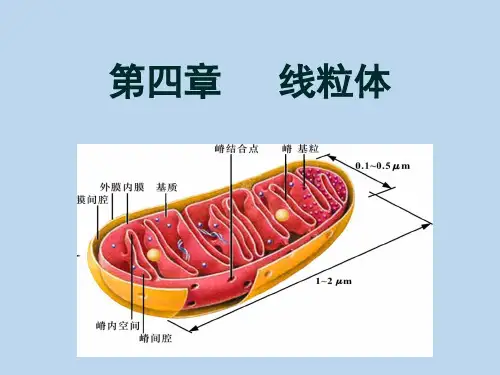
二、线粒体的超微结构在电镜下,线粒体是由两层单位膜套叠而成的封闭性囊状结构,主要由外膜、内膜、膜间隙和基质腔组成。
外膜♦外膜膜厚5-7nm。
磷酸钨复染,外膜有排列整齐的筒状圆柱体。
中间有孔径为2nm的孔,称为孔蛋白。
♦分子量5000以内的物质可以自由通过。
内膜♦平均厚度约4.5nm,通透性很小,分子量大于150的物质无法通过,具有高度选择性。
♦内膜向线粒体内室突出形成嵴。
嵴♦线粒体中形态学变化最大的结构。
♦特征性。
♦主要有两种类型:♦板层状(大多数高等动物细胞中线粒体的嵴);♦小管状(原生动物和其它一些较低等的动物细胞中线粒体的嵴)。
基粒♦内膜和嵴的基质面有许多带柄的小体称为基粒,也称为ATP酶复合体。
♦每个线粒体含有104-105个基粒。
♦形似棒糖,分头、柄和基部三个部分。
♦头部可溶性A TP酶,与基粒柄部合称偶联因子F1;含有ATP酶复合体抑制多肽。
♦柄对寡霉素敏感的蛋白,控制离子通道。
♦基部疏水蛋白,又称偶联因子F0;质子通道。
基质♦在内膜和嵴围成的腔隙中充满的较致密的低电子密度物质。
♦内含大量蛋白质和脂类,包括♦大量重要酶系三羧酸循环酶系、脂肪酸氧化酶系、蛋白质和核酸合成的酶系等。
♦DNA、核糖核蛋白体等。
♦基质颗粒。
第二节化学组成和酶的分布一、线粒体的化学组成♦主要由蛋白质、脂类和水组成。
♦蛋白质65-70%,内膜中含量较多;♦可溶性蛋白:基质中的酶和膜的外周蛋白。
♦不溶性蛋白:镶嵌蛋白、结构蛋白和酶蛋白。
♦脂类25-30%,磷脂为主,内外膜组成不完全相同。
♦与其它膜性结构区别——丰富心磷脂和较少的胆固醇。
二、酶的分布约120种,包括氧化还原酶(37%)、连接酶(10%)、水解酶(<9%)。




线粒体线粒体(mitochondrion)是一种存在于大多数细胞中的由两层膜包被的细胞器,是细胞中制造能量的结构,是细胞进行有氧呼吸的主要场所,被称为"powerhouse"。
其直径在0.5到10微米左右。
除了溶组织内阿米巴、篮氏贾第鞭毛虫以及几种微孢子虫外,大多数真核细胞或多或少都拥有线粒体,但它们各自拥有的线粒体在大小、数量及外观等方面上都有所不同。
线粒体拥有自身的遗传物质和遗传体系,但其基因组大小有限,是一种半自主细胞器。
除了为细胞供能外,线粒体还参与诸如细胞分化、细胞信息传递和细胞凋亡等过程,并拥有调控细胞生长和细胞周期的能力。
大小线粒体是一些大小不一的球状、棒状或细丝状颗粒,一般为0.5-1.0μm,长1-2μm,在光学显微镜下,需用特殊的染色,才能加以辨别。
在动物细胞中,线粒体大小受细胞代谢水平限制。
不同组织在不同条件下可能产生体积异常膨大的线粒体,称为“巨线粒体”(megamitochondria):胰脏外分泌细胞中可长达10-20μm;神经元胞体中的线粒体尺寸差异很大,有的也可能长达10μm;人类成纤维细胞的线粒体则更长,可达40μm。
有研究表明在低氧气分压的环境中,某些如烟草的植物的线粒体能可逆地变为巨线粒体,长度可达80μm,并形成网络。
形状线粒体一般呈短棒状或圆球状,但因生物种类和生理状态而异,还可呈环状、线状、哑铃状、分杈状、扁盘状或其它形状。
成型蛋白(shape-formingprotein)介导线粒体以不同方式与周围的细胞骨架接触或在线粒体的两层膜间形成不同的连接可能是线粒体在不同细胞中呈现出不同形态的原因。
数量不同生物的不同组织中线粒体数量的差异是巨大的。
有许多细胞只拥有多达数千个的线粒体(如肝脏细胞中有1000-2000个线粒体),而一些细胞则只有一个线粒体(如酵母菌细胞的大型分支线粒体)。
大多数哺乳动物的成熟红细胞不具有线粒体。
一般来说,细胞中线粒体数量取决于该细胞的代谢水平,代谢活动越旺盛的细胞线粒体越多。



线粒体名词解释线粒体是细胞内的一种细胞器,存在于几乎所有真核生物的细胞内。
线粒体是细胞的能量合成和供应中心,其主要功能是参与细胞的呼吸作用,通过氧化磷酸化反应产生ATP分子来提供细胞所需的能量。
线粒体还参与合成一些重要的细胞代谢产物,如氨基酸、脂类和胆固醇。
线粒体的结构线粒体呈椭圆形或长圆形,具有双层膜结构,外层膜相对光滑,内层膜有发达的折叠系统,形成许多棒状结构,称为内膜棒。
内膜棒上有许多鳃状突起,称为线粒体旨(cristae),它们增加了线粒体内膜的表面积,提高了呼吸作用和氧化磷酸化的效率。
线粒体内膜与内质网(ER)的外膜相连,形成线粒体-内质网联系。
线粒体的呼吸作用线粒体的呼吸作用是指将生物有机物(如葡萄糖、脂肪酸和氨基酸)氧化分解为二氧化碳和水,释放出大量的能量。
呼吸作用分为有氧呼吸和无氧呼吸两种形式。
有氧呼吸是指在氧气存在的条件下,通过线粒体内的氧化磷酸化过程,将生物有机物完全氧化为二氧化碳和水,并产生ATP分子。
有氧呼吸分为三个阶段:糖解过程、Krebs循环和氧化磷酸化。
糖解过程将葡萄糖分解为丙酮酸,Krebs循环将丙酮酸进一步氧化为二氧化碳,并释放出能量。
氧化磷酸化过程通过电子传递链,将氧化过程释放的能量转化为化学能,合成ATP分子。
无氧呼吸是指在没有氧气的条件下,通过线粒体内的乳酸发酵和酒精发酵过程,将生物有机物氧化为乳酸或乙醇,并释放出一部分能量。
无氧呼吸是在有氧呼吸受限的条件下,细胞为了维持一定的ATP供应而采取的一种代谢途径。
线粒体的其他功能除了参与细胞的呼吸作用,线粒体还具有其他重要功能。
首先,线粒体参与合成一些重要的细胞代谢产物,如氨基酸、脂类和胆固醇。
其次,线粒体参与细胞的离子平衡调节,特别是钙离子的存储和释放。
线粒体内膜上存在有大量的Ca2+通道和Na+/Ca2+交换蛋白,调节细胞内钙离子浓度。
此外,线粒体还参与调节细胞的凋亡(细胞自我死亡)过程,通过释放细胞凋亡信号分子,触发细胞凋亡的级联反应。
![[细胞生物学]线粒体](https://uimg.taocdn.com/3b68c27b5901020206409c00.webp)

医学生物学_线粒体mitochondrion知识介绍线粒体(Mitochondrion)是细胞内的一个重要的细胞器,被称为“细胞的发电厂”,它是细胞内能量代谢的中心。
线粒体含有自己独立的DNA,以及一系列与能量产生和调控相关的酶和蛋白质。
本文将从结构、功能和研究进展三个方面对线粒体进行介绍。
一、线粒体的结构线粒体是椭圆形的细胞器,通常大小约为1至10微米。
它由两层膜组成,外膜是平滑的而内膜形成了许多纵向的褶皱,称为线粒体内膜嵴(cristae)。
线粒体外膜和内膜之间的空间称为间腔,而线粒体内膜和内粒质之间的空间称为内膜间隙。
线粒体内膜上悬有许多小囊泡,称为线粒体间隙质(matrix),其中含有线粒体DNA和许多酶和蛋白质。
二、线粒体的功能线粒体是细胞内能量代谢的中心,其主要功能是通过细胞呼吸产生大部分的细胞能量(ATP)。
线粒体通过氧化磷酸化的过程将食物中的化学能转化为细胞能量。
这个过程包括三个主要的步骤:糖酵解、三羧酸循环和氧化磷酸化。
糖酵解产生的乳酸和氧化磷酸化产生的乙酸和CO2都可以进入线粒体进行进一步的代谢。
另外,线粒体还参与许多其他生物学过程,如细胞凋亡、钙离子调节、脂质代谢和胆固醇合成等。
此外,线粒体还与细胞的免疫应答、衰老过程和许多疾病,例如癌症和神经系统疾病等有关。
三、线粒体的研究进展线粒体的研究涉及多个领域,如结构生物学、生物化学、分子生物学和疾病研究等。
近年来,随着对线粒体的研究的深入,我们对其结构和功能有了更深入的了解,并取得了一些重要的发现。
首先,线粒体的结构被认为是动态可塑的。
线粒体的形状和数量可以根据细胞的需求进行调节。
例如,在细胞凋亡过程中,线粒体会发生形态改变,并释放一系列的细胞凋亡蛋白质。
此外,线粒体还通过与其他细胞器如内质网和高尔基体的相互作用来调节细胞功能。
其次,线粒体在维持细胞稳态和健康方面起到重要作用。
线粒体功能的损害与多种疾病的发生和发展密切相关。
例如,线粒体DNA突变会导致一些遗传性疾病的发生,如线粒体脑肌病和线粒体糖尿病等。
线粒体——世界的幕后统治者线粒体是细胞内微小的细胞器,以ATP的形式生产我们几乎所有的能量。
平均每个细胞里有300-400个线粒体,整个人体里有1亿亿个。
本质上所有的复杂细胞里都有线粒体。
线粒体看上去像细菌,这外观并非伪装:它们从前是自由生活的细菌,后来大约在20亿年前适应了寄生在大细胞里的生活。
它们还保留了基因组的一个碎片,作为曾经独立存在的印记。
它们与宿主细胞之间纠结的关系织成了生命所有的经纬,从能量、性和繁殖,到细胞自杀、衰老和死亡。
线粒体是一个欲盖弥彰的秘密。
许多人都出于各式各样的原因听说过它。
报纸和一些教科书简单地把它描述成生命的“发电厂”——活细胞里微小的发电机,生产我们赖以生存的几乎全部能量。
一个细胞内部有几百或几千个线粒体,它们利用氧来燃烧食物。
线粒体是如此微小,以至于一粒沙里可以轻易地容纳10亿个。
线粒体的进化给生命装上了涡轮发动机,蓄势待发,随时可以启动。
所有动物体内都有线粒体,包括最懒惰的在内。
连不能移动的植物和藻类也要利用线粒体,在光合作用中放大太阳能那无声的轰鸣。
有些人更熟悉“线粒体夏娃”这个词,按照推测,她是所有当代人最晚近的共同祖先——如果我们沿母系血统追踪遗传特征,从女儿到母亲再到外祖母,直至上溯到远古的迷雾中。
线粒体夏娃是所有母亲的母亲,她被认为大约生活在17万年前的非洲,又称“非洲夏娃”。
我们之所以能通过这样的方式追踪遗传上的祖先,是因为所有线粒体都保有小小的一份自己的基因,这些基因仅通过卵子传递给下一代,不通过精子传递。
这意味着,线粒体基因起着母系姓氏的作用,使我们可以沿母系血统追溯祖先,就像有些家族努力沿父系血统把家世追溯到征服者威廉、诺亚乃至穆罕默德。
近来,这其中的某些观念受到挑战,但大体上的理论仍然成立。
当然,这项技术不仅可以使我们知道谁是我们的祖先,也可帮助澄清谁不是我们的祖先。
根据线粒体分析,尼安德特人并未与现代智人杂交,而是在欧洲的边缘被排挤到灭绝。
线粒体还因为它们在法医学上的运用而成为新闻热点。
通过线粒体分析可以确定人或尸体的真实身份,有几个著名的案子运用了这一点。
末代沙皇尼古拉二世的身份,就是通过将其线粒体与亲属的进行比较而得到确认。
第一次世界大战末期,一个17岁女孩从柏林的一条河里被救起,她自称是沙皇失踪的女儿安娜斯塔西娅,随后她被送往一家精神病院接受治疗。
经过70年的纷争,她的说法终于在她于1984年去世后被线粒体分析否认。
更近一些的事例是,世贸中心劫后那些无法辨认的遇难者遗骸是由线粒体基因识别的。
将“正版”萨达姆·侯赛因与他的众多替身之一区分开来,也是靠这种技术。
线粒体基因之所以如此有用,部分是因为它们大量存在。
每个线粒体含有5至10份基因副本,一个细胞里通常有数以百计的线粒体,也就有成千上万份同样的基因,而细胞核(细胞的控制中心)里的基因只有2份副本存在。
因此,完全无法提取任何线粒体基因的情况是很少见的。
一旦线粒体基因被提取出现,基于我们与母亲和母系亲属拥有相同线粒体基因的事实,通常就可以确认或否定设想中的亲属关系。
有一个理论叫做“衰老的线粒体理论”,说的是衰老和许多与此有关的疾病是由在正常细胞呼吸中从线粒体里泄漏出来的活跃分子——自由基导致的。
线粒体并不能完全“防火花”,它们在利用氧燃烧食物时,自由基的火花会逃逸出来,损害邻近的结构,包括线粒体基因本身,以及远处的细胞核基因。
我们细胞里的基因每天要受到1万至10万次自由基攻击,实际上每秒就有一次。
大部分这类损伤很快就会得到修复,不会造成别的麻烦,但偶尔有些攻击会导致无法逆转的变异——基因序列发生持久的改变——这些变异会在一生中累积起来。
受破坏更严重的细胞会死亡,稳定的细胞损耗是衰老和退行性疾病的基础。
许多令人痛苦的遗传疾病也与自由基攻击线粒体基因产生的变异有关。
这些疾病通常有着奇异的遗传模式,其严重性在各世代中会有所不同,但总的来说它们都会随着衰老而趋于恶化。
线粒体疾病通常影响新陈代谢活跃的组织如肌肉和脑,导致癫痫、部分运动失调、失明、耳聋和肌肉退化。
还有一些人熟悉线粒体是因为它是一种具有争议的不育症治疗手段。
从健康女性供体的卵子(卵母细胞)中提取线粒体,移植到患不育症的女性的卵子中,这种技术称为“卵胞质移植”。
它在媒体上首次露面是在一家英国报纸上,报道的标题用彩色字写道“二母一父的婴儿诞生“。
这篇典型的媒体产品并非完全错误——细胞核里的所有基因来自”真正的“母亲,而部分线粒体基因来自”供体“母亲,所以这些婴儿确实从两个不同的母亲那里遗传了一部分基因。
尽管这项技术诞育了30多名显然很健康的婴儿,但它后来在英国和美国都被宣布为非法。
线粒体甚至在电影《星球大战》中登场,冒充对著名的、愿它与你同在的原力的科学解释,这让一些星战影迷非常恼火。
在前期的电影中,原力即使不算宗教也是唯灵的,但在后来的一部电影中被解释成“迷地原虫”(midichlorians)的产物。
一位乐于助人的绝地武士说,迷地原虫是“栖居在所有活细胞里的微小生命形式,我们和它们是共生体,为共同的利益生活在一起。
没有迷地原虫,生命就不可能存在,我们也无从了解原力。
”迷地原虫在名字和行为上与线粒体的相似之处令人无法错认,这是设定者有意为之。
线粒体的祖先是细菌,它们作为共生体(与其它生物存在互利关系的生物)生活在我们的细胞里。
与迷地原虫一样,线粒体也有许多神秘的特征,甚至可以形成有许多分枝的网络,能够相互通信。
Lynn Margulis 在20世纪70年代使这个一度存在争议的理论变得非常出名,到现在,线粒体的细菌起源已经成为被生物学家所接受的事实。
线粒体的这些方面已经通过报纸和大众文学为许多人所熟知,其它方面在过去的一二十年里在科学界广为人知,但对大众来说也许还较为神秘。
其中最重要的一点是凋亡,即细胞的程序性死亡,在这一过程中,细胞个体为了大局利益——以身体为整体——而自杀。
大约从20世纪90年代中期开始,研究人员发现凋亡并不是像从前认为的那样受细胞核里的基因控制,而是受线粒体控制。
这一发现对医学研究有着重要意义,因为细胞无法在需要时凋亡是癌症的根源。
许多研究人员现在尝试通过某种方式操纵线粒体,而不以细胞核里的基因为靶标。
但这其中还有着更深远的意义。
在癌症中,细胞个体寻求自由,摆脱了为有机体整体负责的桎梏。
在早期进化中,把这种桎梏加在细胞上想必是很困难的:一个有独立生存潜力的细胞,在还可以选择离开群体独自生活的时候,凭什么要为了生活在细胞群体里的利益而接受死刑?没有程序性死亡,将细胞联结起来形成复杂多细胞生物的纽带也许永远也不会进化出来。
由于程序性死亡依赖于线粒体,也许可以说没有线粒体就没有多细胞生物。
为了这免得听起来太古怪,需要说明的是所有多细胞植物和动物确实含有线粒体。
线粒体占有突出地位的另一个领域是真核细胞的起源。
真核细胞是有着细胞核的复杂细胞,所有的植物、动物、藻类和真菌都是由真核细胞构成的。
“真核”(eukaryotic)一词源自希腊语“真正的核”,指细胞内部基因的所在地。
但这个名字是有明显缺陷的。
事实上,真核细胞除细胞核之外还包含许多其它部件,包括著名的线粒体。
这类复杂细胞怎样进化而来,是一个热点话题。
人们一般认为,它们逐步进化,直到有一天某个原始的真核细胞吞噬了一个细菌,后者在经过许多世代的奴役之后,最终变得完全寄人篱下,进化成了线粒体。
这个理论预言,某些不包含线粒体的、籍籍无名的单细胞真核生物将被发现是我们所有人的祖先,它们是从线粒体被“捕获”并投入使用之前的那些岁月存留下来的孑遗。
但在经过了十来年详细的遗传分析之后,人们现在发现似乎所有的已知真核细胞都拥有或者曾经拥有(后来丢失了)线粒体。
这意味着复杂细胞的起源与线粒体的起源不可分割,它们是同一个事件。
如果这是真的,那么不仅多细胞生物的进化需要线粒体,构成多细胞生物的部件——真核细胞的进化也需要线粒体。
而如果这是真的,那么如果没有线粒体,地球生物不会进化成细菌以外的东西。
线粒体的另一个秘密领域与两性的区别有关,事实上它是两性存在的必要条件。
性是一个著名的谜:有性生殖需要父母双方来产生一个后代,无性生殖或孤雌生殖则只需要母亲一方,父亲的存在不仅多余,而且是对空间和资源的浪费。
更糟糕的是,两性的存在意味着我们只能在人口的一半中寻找配偶,至少在我们把性当作生殖手段时是这样。
不管是不是为了生殖,如果所有的人都是同性,或者性别多到近乎无限,情况会好得多:两性是所有可能的局面中最糟糕的一种。
这个谜题的答案之一与线粒体有关,该理论于20世纪70年代晚期出现,现在已被科学界广泛接受,也许公众对其了解相对较少。
该理论认为,我们必须有两种性别,是因为一种性别必须专门负责通过卵子把线粒体传递下去,而另一种性别必须专门地通过精子不把线粒体传递下去。
本书第6部分将详细阐述这一点。
所有这些研究领域使线粒体重新取得了它在20世纪50年代的鼎盛时期过后再也不曾拥有的重要地位,当年人们首次证实线粒体是细胞的动力来源,生产我们所需的几乎所有能量。
顶尖学术杂志《科学》在1999年充分认识到这一点,把一期封面和相当大的篇幅给了线粒体,标题为“线粒体又回来了”。
这种忽视有两个主要原因。
其一是生物能量学——研究线粒体中能量产生过程的科学——被认为是一个艰难而且模糊的领域,有一句曾在各种学术报告厅的窃窃私语中流传的保证对此作了漂亮的总结:“别担心,谁都听不懂线粒体学家们(mithchondriacs)在说什么。
”第二个原因与20世纪下半叶分子遗传学的起源有关。
就像著名的线粒体学家Immo Scheffler说的那样:“分子生物学家们忽视线粒体的原因,可能是他们没有立即认识到线粒体基因这一发现的深远意义及应用前景。
需要很长时间来积累一个范围足够大、内容足够多的数据库,解决与人类学、生物起源、疾病、进化及其他问题有关的诸多挑战。
”我在前面说了,线粒体是一个欲盖弥彰的秘密。
尽管最近享有盛名,但它仍然是个谜。
许多深奥的进化问题人们几乎没有提出过,更不用说经常在学术杂志上讨论。
围绕着线粒体发展起来的不同领域往往实质上局限在自己的圈子里。
例如,线粒体产生能量的机制——将离子泵过膜,称为渗透作用——在所有形式的生命中都存在,包括最原始的细菌,这是非常奇怪的。
用一位评论者的话来说,“自达尔文以来,生物学还没有提过出像爱因斯坦、海森堡和薛定谔的理论那样违反直觉的看法。
”但这个理论被证明是正确的,并使Peter Mitchell 在1978年获得诺贝尔奖。
但人们很少提出这样一个问题:为什么这样一种特定的能量产生方式成为如此多种不同生命的核心?我们将会看到,这个问题的答案将解释生命起源本身。
还有一个非常有意思的问题很少被提及,那就是线粒体基因的持久存在。
学术论文将我们的家谱追溯到线粒体夏娃,甚至利用线粒体基因重建出不同物种之间的关系,但很少问及线粒体基因到底为什么存在,仅仅假定它们是细菌起源的遗迹。