乳糖操纵子
- 格式:pdf
- 大小:2.58 MB
- 文档页数:24

乳糖操纵子复习题
乳糖操纵子是分子生物学中一个重要的概念,它涉及到基因表达的调
控机制。
以下是关于乳糖操纵子的复习题内容:
1. 定义:请简述乳糖操纵子是什么,并解释其在细胞中的作用。
2. 组成:描述乳糖操纵子的基本组成部分,包括启动子、操纵基因、
结构基因等。
3. 调控机制:解释乳糖操纵子的正调控和负调控机制是如何工作的。
4. 诱导:阐述乳糖如何作为诱导剂激活乳糖操纵子的表达。
5. 抑制:描述在没有乳糖的情况下,乳糖操纵子是如何被抑制的。
6. cAMP-CAP复合物:解释cAMP-CAP复合物在乳糖操纵子调控中的作用。
7. 乳糖操纵子的发现:简述乳糖操纵子是如何被发现的,以及这一发
现对分子生物学的意义。
8. 应用:讨论乳糖操纵子在现代生物技术中的应用,特别是在基因工
程和基因治疗中的作用。
9. 比较:将乳糖操纵子与其他类型的操纵子(如色氨酸操纵子)进行
比较,指出它们的异同点。
10. 实验研究:列举一些实验方法,用于研究乳糖操纵子的调控机制。
11. 问题解决:提出一些可能在研究乳糖操纵子时遇到的问题,并给出可能的解决方案。
12. 未来方向:探讨乳糖操纵子研究的未来方向,以及这些研究可能对医学和生物技术带来的影响。
通过这些问题的复习,可以加深对乳糖操纵子及其调控机制的理解,为进一步的学习和研究打下坚实的基础。

乳糖操纵子名词解释乳糖操纵子(lactose operation)能合成和分泌乳糖的一类重要细胞,它们分布在不同类型的细胞内。
乳糖操纵子中的酶系有的是糖苷酶,有的是羧酸酯酶,还有一些是复合酶。
目前已发现的操纵子有七种类型,但只有两个编码,不论是催化水解乳糖还是释放乳糖的酶均是如此。
乳糖操纵子分布在所有高等动物组织中,哺乳类有两种:insulin- like autoantibody-抗胰岛素样蛋白4;一种乳糖操纵子,由四个区域组成,分别编码降血糖蛋白4(hypoglycemic-like autoantibody-抗胰岛素样蛋白4),降血糖蛋白5(hypoglycemic-like albumin-抗胰淀粉样蛋白)和乳糖操纵子自身。
此外尚有由insulin- like autoantibody-抗胰岛素样蛋白4(抗-4)与血浆蛋白G、铁蛋白、转铁蛋白结合的复合体,即乳糖操纵子复合体(oligosaccharide-like autoantibody complex-球蛋白操纵子复合体),也可能存在于不同细胞。
其中抗-4分子量为50万,具有与抗-4同源的抗胰岛素样蛋白4抗原决定簇,不受胰岛素影响,当它和其它球蛋白合成后,会结合于巨噬细胞膜上,并被膜内的锌粒子中和,再与巨噬细胞内的受体结合,从而阻断胰岛素与受体结合,进入细胞内的胰岛素失去降血糖作用。
至今只发现一种能降低血糖的操纵子,此种操纵子也称为受体型操纵子。
此种操纵子是位于酪氨酸磷酸酶基因上游,酪氨酸激酶基因下游,有关的其他基因几乎均已克隆。
当激活后,位于上游的酪氨酸磷酸酶基因激活使细胞内游离的酪氨酸浓度增加,酪氨酸水解成磷酸肌醇和磷酸胆碱释放入血液循环中,这将使血糖降低;而酪氨酸磷酸酶则与血清白蛋白结合,阻止白蛋白转运氨基酸,抑制氨基酸通过白蛋白进入血液,也可以抑制外周组织对氨基酸的利用。
该操纵子中的受体称为“受体酪氨酸磷酸酶”。
该操纵子也可能参与葡萄糖和脂肪酸的代谢。


乳糖操纵子乳糖操纵子是参与乳糖分解的一个基因群,由乳糖系统的阻遏物和操纵序列组成,使得一组与乳糖代谢相关的基因受到同步的调控。
1961年雅各布(F.Jacob)和莫诺德(J.Monod)根据对该系统的研究而提出了著名的操纵子学说。
在大肠杆菌的乳糖系统操纵子中,β-半乳糖苷酶,半乳糖苷渗透酶,半乳糖苷转酰酶的结构基因以LacZ(z),Lac Y(y),Lac A(a)的顺序分别排列在染色体上,在z的上游有操纵序列Lac O(o),更前面有启动子Lac P(p),这就是操纵子(乳糖操纵子)的结构模式。
编码乳糖操纵系统中阻遏物的调节基因Lac I(i)位于和p上游的临近位置。
细菌相关功能的结构基因常连在一起,形成一个基因簇。
它们编码同一个代谢途径中的不同的酶。
一个基因簇受到同一的调控,一开俱开,一闭俱闭。
也就是说它们形成了一个被调控的单位,其它的相关功能的基因也包括在这个调控单位中,例如编码透过酶的基因,虽它的产物不直接参与催化代谢,但它可以使小分子底物转运到细胞中。
乳糖分解代谢相关的三个基因,lacZ、Y、A就是很典型的是上述基因簇。
它们的产物可催化乳糖的分解,产生葡萄糖和半乳糖。
它们具有顺式作用调节元件和与之对应的反式作用调节因子。
三个结构基因图的功能是:lacZ编码β-半乳糖苷酶(β-galactosidase),此酶由500kd的四聚体构成,它可以切断乳糖的半乳糖苷键,而产生半乳糖和葡萄糖lacY编码β一半乳糖苷透性酶(galactoside permease),这种酶是一种分子量为30kDd膜结合蛋白,它构成转运系统,将半乳糖苷运入到细胞中。
lacA编码β-硫代半乳糖苷转乙酰基酶(thiogalactosidetransacetylase),其功能只将乙酰-辅酶A上的乙酰基转移到β-半乳糖苷上。
无论是lacZ发生突变还是lacY发生突变却可以产生lac-型表型,这种lac-表型的细胞不能利用乳糖。

乳糖操纵子的原理
乳糖操纵子是真核生物基因中一段编码乳糖酶的 DNA序列。
该基因的发现,为研究在人体内存在的、与乳糖代谢有关的酶提供了线索。
我们知道,牛奶是由各种不同类型的乳糖组成,即半乳糖和葡萄糖。
牛奶中除了这两种成分外,还含有其他一些成分,如钙、磷、铁等。
我们人体不能合成这些成分,必须由食物来补充。
乳糖是一种简单的碳水化合物,在人类和哺乳动物体内都存在。
人和动物食用牛奶后,小肠中的乳糖酶就会将其分解成葡萄糖和半乳糖。
在这一过程中,半乳糖苷被水解成单糖,进入大肠中与细菌产生的细菌素结合,细菌素进一步分解成酸和二氧化碳,经肠道排出体外。
这种由碳水化合物分解成单糖和二氧化碳的过程称为“分解代谢”。
乳糖是哺乳动物乳汁中最重要的碳水化合物成分之一。
在婴儿出生后3~6个月内,主要是靠母乳来提供能量。
在哺乳期内,由于母亲体内乳糖酶活力下降或缺乏,以及婴儿消化道尚未发育成熟等原因,母乳中的乳糖酶活性很低或缺乏。
—— 1 —1 —。



乳糖操纵子名词解释乳糖操纵子:乳糖操纵子也称为乳糖运输蛋白,是一种催化乳糖分解的酶。
表1乳糖操纵子名词解释 1。
催化酶。
由于其构成的蛋白质分子中有一个特殊序列(N),即所谓“不对称区”,在这个区域内有两种催化结构的酶同时起作用,因此对绝大多数天然底物有高度选择性。
乳糖是一种低聚糖,它与人体所需要的葡萄糖、半乳糖、阿拉伯糖等都属于单糖,在人体内具有特殊的功能,是糖类的主要成分之一,被吸收后参与新陈代谢。
因此,乳糖可作为食品添加剂应用到食品中去,作为提高人们生活水平的功能食品。
在食品工业上主要用来制造无糖糕点和饼干、奶粉、冰淇淋等,并将乳糖经发酵变成乳酸,以提高甜度,改善风味。
2。
乳糖操纵子。
存在于胞质溶胶内,又称胞质溶胶。
分布于质膜下,含有一个多肽复合物,由100多个氨基酸残基组成,不完全相同的数目超过10万个,该区域为不对称的α螺旋形结构。
3。
表位( mRNA)。
存在于胞质溶胶的不对称区,由RNA聚合酶在转录前的间隔期间转录。
表位是一种特殊的RNA,除了与操纵子有关外,还在RNA结构和进化中扮演着重要角色。
4。
基因家族。
位于基因簇,位于多个细胞中,组织中或系统中。
5。
编码区( coding region)。
基因组DNA上特定位置,在大多数哺乳动物和鸟类的基因中,在前驱物导入时,必须改变。
6。
操纵子( modulator)。
在一个基因簇上控制转录的一段DNA序列,有时可以是RNA。
这些序列有几个特征,例如与位于转录起始位置的操纵元结合,与酶有关联的氨基酸序列。
7。
基因簇( locus)。
在不同种或同种不同个体间,共享一段相同的核苷酸序列的核苷酸序列集合,包括基因。
8。
微卫星( Microsatellite)。
在染色体的DNA序列中,短的重复序列,常常出现的两个重复序列之间的间隙,是连续的DNA序列中的一段。
9。
小卫星( mini satellite)。
在染色体的DNA序列中,长的重复序列,常常出现的三个重复序列之间的间隙,是连续的DNA序列中的一段。

原核生物中,乳糖操纵子是一种在乳糖存在时调控基因表达的元件。
这种调控机制广泛存在于大肠杆菌等细菌中,它允许细菌在环境中检测到乳糖的存在并调整相关基因的表达。
以下是原核生物中乳糖操纵子基因表达调控的基本原理:
1. 乳糖操纵子的组成:
- 乳糖操纵子包括两个基本部分,一个是操纵子的操作元件(operator),另一个是调控基因的操纵子结合蛋白(repressor protein)。
2. 操作元件(Operator):
- 操纵子的操作元件是一个DNA序列,位于被调控的基因的上游区域。
- 操纵子的操作元件是乳糖操纵子的结合位点,调控蛋白可以与其结合。
3. 调控基因的操纵子结合蛋白:
- 调控基因的操纵子结合蛋白通常是一个负调控因子,即在没有乳糖的情况下,它会结合到操作元件上,阻止RNA聚合酶的结合,从而抑制基因的转录。
4. 乳糖的作用:
- 当细菌环境中存在乳糖时,乳糖分子会与调控基因的操纵子结合蛋白发生结合。
- 乳糖结合到操纵子结合蛋白后,导致蛋白的构象发生变化,无法再结合到操纵子的操作元件上。
5. 操纵子的操作元件的解离:
- 由于操纵子结合蛋白不能再结合到操作元件上,RNA聚合酶得以在操作元件上结合并启动被调控基因的转录。
6. 基因的表达:
- 乳糖操纵子的解离使RNA聚合酶能够转录下游基因,从而启动基因的表达,产生相关的蛋白质。
通过这个机制,原核生物能够根据环境中乳糖的存在与否,灵活地调控基因的表达,以适应不同的代谢和生存需求。
这种调控机制是一种典型的负调控,其中乳糖的存在解除了负调控因子对基因的抑制。

乳糖操纵子引言乳糖操纵子是一种能够操纵乳糖代谢的物质或机制的称谓。
乳糖是一种存在于奶制品中的碳水化合物,而对乳糖的消化和代谢需要人体内的乳糖酶来完成。
然而,有些人的体内缺乏乳糖酶,导致乳糖无法被消化和代谢,从而引发乳糖不耐受症状。
乳糖操纵子的出现为乳糖不耐受症患者提供了新的治疗和饮食选择。
本文将介绍乳糖操纵子的定义、分类以及应用等相关内容。
乳糖操纵子的定义乳糖操纵子是一种能够通过调节乳糖代谢的物质或机制。
乳糖操纵子可以分为两类:一类是能够增强乳糖酶活性的物质或机制,另一类是能够减弱或抑制乳糖酶活性的物质或机制。
通过作用于乳糖酶或其他相关代谢途径,乳糖操纵子可以改变人体对乳糖的代谢过程,从而影响乳糖不耐受症状的发生和程度。
乳糖操纵子的分类根据其对乳糖酶活性的影响,乳糖操纵子可以分为以下几类:1.增强型乳糖操纵子:这类乳糖操纵子能够增加乳糖酶的活性,提高乳糖的代谢速度。
常见的增强型乳糖操纵子包括乳糖酶替代物以及对乳糖酶具有促进作用的物质。
2.抑制型乳糖操纵子:这类乳糖操纵子能够减弱或抑制乳糖酶的活性,降低乳糖的代谢速度。
常见的抑制型乳糖操纵子包括乳糖酶抑制剂以及对乳糖酶具有抑制作用的物质。
乳糖操纵子的应用随着对乳糖不耐受症的研究深入,乳糖操纵子逐渐被应用在乳糖不耐受症的治疗和饮食调节中。
下面是乳糖操纵子在不同应用场景中的具体应用:1.药物治疗:某些乳糖操纵子可以作为药物用于治疗乳糖不耐受症。
例如,一些乳糖酶替代物可以在人体内代替缺乏的乳糖酶,帮助消化和代谢乳糖。
2.膳食调节:乳糖操纵子还可以通过膳食调节的方式应用于乳糖不耐受症的患者。
例如,饮食中可以添加含有增强型乳糖操纵子的食品,增加乳糖酶的活性,从而促进乳糖的代谢。
3.乳制品加工:乳糖操纵子的应用还可以扩展到乳制品的加工过程中。
例如,在酸奶的生产中可以添加抑制型乳糖操纵子,降低乳糖的含量,从而适应乳糖不耐受症患者的需求。
总结乳糖操纵子作为一种能够调节乳糖代谢的物质或机制,为乳糖不耐受症的患者提供了新的治疗和饮食选择。
乳糖操纵子的转录调控原理介绍乳糖操纵子(lac operon)是一种在大肠杆菌中广泛存在的基因调控系统。
它对乳糖的利用起到重要作用,并在转录水平上对乳糖降解相关基因进行调节。
本文将详细探讨乳糖操纵子的转录调控原理。
乳糖操纵子的组成乳糖操纵子由三个主要部分组成:乳糖酶基因(lacZ)、乳糖转运蛋白基因(lacY)和乳糖重pressor基因(lacI)。
下面将对每个部分进行详细介绍。
lacZ基因lacZ基因编码乳糖酶(β-galactosidase),它能将乳糖分解为葡萄糖和半乳糖两种单糖。
乳糖酶的产生对于细菌能够利用乳糖作为碳源至关重要。
lacY基因lacY基因编码乳糖转运蛋白(lactose permease),它能将乳糖从细胞外转运到细胞内。
乳糖转运蛋白的存在使得细菌能够主动吸收外源性乳糖。
lacI基因lacI基因编码乳糖重pressor(lactose repressor),它是乳糖操纵子的主要调控因子。
乳糖重pressor能够结合到乳糖操纵子上的操纵子区域,从而抑制lacZ和lacY基因的转录。
转录调控的原理乳糖操纵子的转录调控主要通过乳糖重pressor和乳糖的存在与否来实现。
下面将分别介绍两种情况下的转录调控原理。
乳糖存在时的转录调控当细菌培养基中存在乳糖时,乳糖分子能够结合到乳糖重pressor上,从而改变其构象,使其无法结合到操纵子区域上。
这样一来,lacZ和lacY基因的转录将不再受到抑制,从而使乳糖酶和乳糖转运蛋白的产生得以增加。
乳糖缺失时的转录调控当细菌培养基中缺乏乳糖时,乳糖重pressor无乳糖结合,能够结合到操纵子区域上,从而阻止lacZ和lacY基因的转录。
这种抑制作用是通过乳糖重pressor与RNA聚合酶的相互作用来实现的。
转录调控的细节机制乳糖操纵子的转录调控不仅仅是简单的开关机制,其中还涉及到一些细节的调控机制。
下面将介绍其中的几个重要细节。
CAP-cAMP复合物的作用CAP(catabolite activator protein)是一种转录激活蛋白,与cAMP(cyclic AMP)结合后形成CAP-cAMP复合物。
乳糖操纵子名词解释乳糖操纵子( lac- G cluster),指能够对半乳糖等六种乳糖分子进行跨膜转运的转运体。
相关酶存在于高尔基体上的乳糖操纵基因( lac- G)调控亚单位(操纵子)内。
通常一个亚基由一个DNA 分子(操纵子)及若干个操纵蛋白(亚基)所组成,蛋白质为单顺反子。
当前的研究表明,乳糖操纵子可分为三个基本组件:操纵基因、蛋白转运亚基及转运蛋白。
在每一亚基中,均含有一对核酸( DNA或RNA)和蛋白质。
蛋白质的氨基酸序列和其相应的顺反子序列都是保守的,而与这些保守的氨基酸序列相互作用的核苷酸序列则因亚基而异。
操纵基因可以有若干个,通常是整合到基因组上;有些细菌的转运体可包括四个基因。
在不同的细菌间,同一乳糖操纵子也存在很大的差异,甚至同一菌属不同菌株间也存在着相当大的差异。
各种转运体之间有极其复杂的交叉现象,导致乳糖操纵子具有非常高的多样性。
乳糖操纵子的结构十分保守,对于鉴定遗传标记具有重要意义。
同一乳糖操纵子存在不同的亚基,其生理功能也有所差异。
一般来说,乳糖操纵子各亚基的基本功能是不同的,其主要区别有以下几点:( 1)转运亚基的形态和位置,取决于该亚基所携带的乳糖分子的大小和电荷。
此外,一个亚基往往含有两种或两种以上的乳糖分子,从而使该亚基的活性受到不同程度的影响。
( 2)转运蛋白的空间结构与转运性质。
转运蛋白以一种特定的方式结合在内质网上的特殊位点上,在转运过程中对于转运方向起重要作用。
人类及其它哺乳动物细胞中有多种转运体,这些转运体并不编码一种乳糖分子,但都有类似的结构,并且能将外源性乳糖分子从胞外转运至胞内。
不同细胞中乳糖操纵子的亚基数目可有很大差别,例如,分泌性胃肠道的Lac- G cluster包括一个编码前导蛋白的操纵基因,一个与受体蛋白结合的转运蛋白,还有两个负责ATP生成的转运蛋白亚基。
在乳糖操纵子中,有一部分的Lac- G亚基是不编码蛋白质的,这些亚基称为操纵基因。
乳糖操纵子的基本调控过程1.引言1.1 概述乳糖操纵子是一种广泛存在于真核生物中的基因调控系统,它在乳糖代谢过程中发挥重要作用。
乳糖是一种双糖,由葡萄糖和半乳糖组成,可被一些生物利用。
在某些条件下,乳糖操纵子可以调控与乳糖相关的基因的表达水平。
乳糖操纵子通常由两个组分组成:乳糖操纵子启动子和乳糖操纵子结合蛋白。
乳糖操纵子启动子位于被调控基因的上游区域,其中包含着与乳糖操纵子结合蛋白相互作用的DNA序列。
乳糖操纵子结合蛋白则通过与乳糖操纵子启动子结合,调控被调控基因的转录水平。
乳糖操纵子的基本调控过程涉及到乳糖操纵子结合蛋白的结合和解离,以及与乳糖的结合。
当环境中存在乳糖时,乳糖操纵子结合蛋白会与乳糖结合,形成复合物,进而与乳糖操纵子启动子结合。
这个复合物会招募其他转录因子和RNA聚合酶,促使被调控基因的转录。
而在乳糖缺乏的环境下,乳糖操纵子结合蛋白会解离,并不再与乳糖操纵子启动子结合,阻碍被调控基因的转录。
乳糖操纵子的基本调控过程对于真核生物的生存和生长具有重要意义。
它可以帮助真核生物适应环境中乳糖的变化,从而合理利用有限的营养资源。
同时,乳糖操纵子的基本调控过程也为我们研究基因调控的机制提供了一个良好的模型系统。
通过深入研究乳糖操纵子调控过程的细节,我们可以更好地理解基因的表达调控网络,为疾病的诊断和治疗提供新的途径。
在本文中,我们将详细讨论乳糖操纵子的基本调控过程的要点,以及其对真核生物的重要意义。
通过对其相关机制的探究,我们希望能够增进对基因调控的理解,并为进一步的研究提供启示。
同时,我们也将展望未来在这一领域的研究方向,为乳糖操纵子的研究和应用提供新的思路和方法。
1.2 文章结构文章结构部分的内容可以包括以下内容:文章按照以下结构组织:引言、正文和结论。
- 引言部分介绍了乳糖操纵子的研究背景和重要性,概述了本文要阐述的乳糖操纵子的基本调控过程,并简要介绍了文章的结构。
- 正文部分包括了乳糖操纵子的基本调控过程要点1和要点2的详细介绍。
乳糖操纵子的名词解释
乳糖操纵子是一种食物添加剂,是乳糖的一种合成代用品,通常用于替代糖分,改善口感和结构,降低糖分含量,尤其是低盐产品。
它的合成原理就是以甲醇(乙醇)为主要原料,加入不同类型的乙酸或醋酸钠,进行加氢反应,最后再经过糖化反应制成的糖分。
乳糖操纵子的类型有乳糖磷酸酯、乳糖酯类、乳糖酮类,以及乳糖衍生物多种多样,例如乳糖醛酸酯。
其中乳糖磷酸酯类是由乙酸磷酸和乙醇反应制成的,乳糖酯类则是由乳酸和乙醇反应生成的,乳糖酮类是由醋酸酯和乙醇反应生成的,乳糖衍生物则是由醋酸酯和乙醇反应生成的,最终形成不同糖分。
乳糖操纵子的特点是拥有新颖的口感和较低的热量,而且乳糖替代的热量比糖分更低,具有较强的抗氧化能力,从而降低糖分含量,从而帮助改善血糖控制,从而让使用者不受糖尿病影响。
乳糖操纵子目前应用范围很广,如果冰激凌、饮料和西点中都有乳糖操纵子的存在,点心面包和奶酪也非常常见,这是因为它们的口感更微妙,而乳糖操纵子可以改善食物的口感,并降低糖分含量,从而防止血糖过高。
此外,乳糖操纵子有助于改善血糖控制,可以帮助糖尿病患者预防并发症的发生,从而提高质量生活。
因此,乳糖操纵子可以被广泛应用于食品领域,它可以改善食物的口感,降低糖分,而且还有利于改善血糖控制。
尽管乳糖操纵子有许多优点,但我们仍然需要谨慎使用,让我们不要过度摄入。
我们可以选择更多的低糖和低盐的食物,以及经常进行体育锻炼,以增强我们的体质,减少糖尿病风险,更好的保护我们的健康。
原核生物基因表达转录水平调控之乳糖操纵子模型(2012-07-13 00:37:45)转载▼原核生物基因表达在转录水平上的调控最经典学说是操纵子学说。
一、操纵子细菌基因表达调控的许多原理是在研究E.coli乳糖代谢调节时被发现的。
法国巴斯德研究院的Francois Jacob与Jacques Monod于1960年在法国科学院院报(Proceeding of the French Academy of Sciences)上发表了一篇论文,提出乳糖代谢中的两个基因被一靠近它们的遗传因子所调节。
这二个基因为β半乳糖苷酶(β-galactosidase)和半乳糖苷透过酶(galactoside penmase)。
前者能水解乳糖成为半乳糖和葡萄糖,后者将乳糖运输到细胞之中。
在此文中他们首先提出了操纵子(operon)和操纵基因(operator)的概念,他们的操纵子学说(theory of operon)使我们得以从分子水平认识基因表达的调控,是一个划时代的突破,因此他们二人于1965年荣获诺贝尔生理学奖。
Jacob与Monod所提出的关于基因表达调控的操纵子学说可以简述如下:有一个专一的阻遏分子(蛋白质)结合在靠近β半乳糖苷酶基因上面,这段DNA他们称之为操纵基因。
由于阻遏分子结合在DNA的操纵基因上,从而阻止了RNA聚合酶合成β半乳糖苷酶的mRNA。
此外,他们还指出乳糖为诱导物,当乳糖结合到阻遏分子上时,即阻止阻遏分子与操纵基因的结合。
当有乳糖时,阻遏分子即失活,mRNA就可以转录出来。
如果去掉乳糖时,阻遏分子又恢复其活力,与操纵基因DNA结合,将乳糖基因关闭。
二、乳糖操纵子/fzswx/knowledge/knowledge01.asp?zsdBianhao=060302/s/blog_4b07ffbc01016v21.html乳糖操纵子(lac operon)是原核生物中研究得最清楚的一种操纵子。
14原核生物基因的表达调控 生物体在其生命活动中,基因的表达严格有序,任何影响到基因开启与关闭、转录和翻译等基因表达程序的调节作用,都属于对基因表达的调控。
原核生物是单细胞生物,没有核膜和明显的核结构。
它们与周围环境关系密切。
在长期进化过程中产生了高度的适应性和应变能力,这是它们赖以生存的保证。
由此可见,原核生物的基因表达既与自身的遗传结构相适应,又体现了它们对环境的应变能力。
原核生物基因表达调控主要发生在转录水平上,这可以最经济地在基因表达的第一步实行最有效的控制。
原核生物以操纵子为单位的调控系统即体现了这一特点。
然而,转录调控的方式多种多样,如噬菌体基因表达的时序调控;大肠杆菌色氨酸合成代谢的衰减调控,即是转录调控的明显例证。
此外,也有许多翻译水平上的调控机制,如核糖体蛋白质合成的自身调节;反义RNA或小RNA对mRNA翻译的调控作用等等。
有时,原核生物甚至还能从DNA水平上对基因表达进行调节,如沙门氏杆菌的相变过程,就是以基因重排的方式调控基因转录。
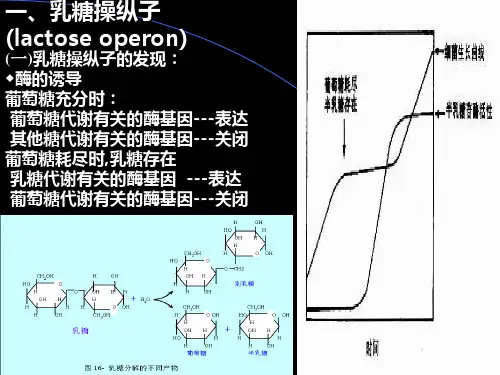
327 14畅1 大肠杆菌乳糖操纵子的调控机制14畅1畅1 大肠杆菌对乳糖的利用和酶诱导 早在20世纪初期就发现,酵母细胞只有在某种底物存在时才产生相应的酶。
这种由底物诱导而产生酶的效应,称为诱导作用(i nducti on )。
酶诱导普遍存在于细菌中,如大肠杆菌(E 畅co li )的乳糖利用系统便是诱导过程的典型例证。
大肠杆菌的乳糖代谢需要有β半乳糖苷酶(βgalactosidase)的催化,该酶能把乳糖水解为半乳糖(gal acto se )和葡萄糖(g l u co se )(图141)。
如果在大肠杆菌的培养基中所用的碳源不是乳糖,而是其他种类的糖(如葡萄糖),那么细胞内的β半乳糖苷酶的分子极少,平均只有0畅5~5个分子。
可是,一旦培养基的碳源完全用乳糖取代葡萄糖,则在2~3m i n 内,细胞中就合成了大量β半乳糖苷酶分子,数量骤增,分子数可达1000~10000个。
当从培养基中除去半乳糖,细菌很快就停止合成β半乳糖苷酶。
显然,新合成的β半乳糖苷酶是在底物乳糖诱导下产生的。
可见,乳糖是合成β半乳糖苷酶的诱导物,而β半乳糖苷酶是可诱导酶(i n duci b l e enzym e )。
这个系统称为可诱导系统(i nduci b l e system )。
大肠杆菌对乳糖的分解利用,除了需要β半乳糖苷酶外,还需要半乳糖苷透性酶(gal acto si de permease )。
半乳糖苷透性酶是一种膜蛋白,可协助乳糖分子穿膜进入细胞。
除上述两种酶外,还产生了硫代半乳糖苷转乙酰基酶(thi ogal acto si de transacetyl ase )。
14畅1畅2 大肠杆菌乳糖操纵子的负控制 为解释上述现象,1961年法国分子生物学家F 畅Jacob 和J 畅M onod 通过对大肠杆菌乳糖代谢系统的一系列研究,根据其基因的活动和表达的调节提出了操纵子学说(operon hypo thesis )。
实验证明,3种蛋白质:β半乳糖苷酶(Z )、半乳糖透性酶(Y )和硫代半乳糖苷转乙酰基酶(A )的编码基因l a cZ、l acY 图141 乳糖操纵子的结构(引自G riffiths 等,2005)和l acA 依次连接在一起,形成了一个转录单位。
操纵子学说主张,该转录单位的转录是从启动子14畅1 大肠杆菌乳糖操纵子的调控机制14 原核生物基因的表达调控328 (p rom o ter,P)开始,并受操纵基因(opera to r,O)和调节基因(regul ato r,I)的控制。
启动子和操纵基因位于乳糖结构基因的上游,依次互相连接。
这样依次排列的P,O,Z,Y,A序列片段便构成了一个共表达的遗传单位———乳糖操纵子(l ac operon)(图141)。
调节基因是一个独立的转录单位,它有自己的启动子。
其表达产物为阻遏物(rep resso r),阻遏物既能阻止转录,又能识别小分子的诱导物。
乳糖结构基因能否转录为mRN A受到操纵基因的控制。
阻遏物同操纵基因结合则使结构基因关闭,未结合,基因则开启。
因此,具有活性的阻遏物只要结合到操纵基因上,即可阻断RN A聚合酶的转录活动。
实际上,P和O在序列上有一定序列重叠,O被阻遏物占据时,RN A聚合酶就不可能与P结合,因而不能催化转录。
阻遏物是否具有活性要受诱导物的影响,它一旦与诱导物结合,便会发生构象变化,而丧失活性,不再能结合操纵基因。
在这种情况下,RN A聚合酶可顺利地通过操纵基因,启动结构基因的转录,3个基因所转录出的mRN A进而翻译成物质(图141)。
在乳糖操纵子系统中,当没有乳糖等一类诱导物时,阻遏物与操纵序列结合,使结构基因不能表达。
只有加入小分子诱导物,于是阻遏物失活,结构基因才能表达。
可见,调节基因表达的阻遏物的作用在于阻止转录,这种作用原理称为负控制(nega ti ve contro l)。
乳糖操纵子学说已为实验所证实,为原核生物基因表达调控机制提供了重要的模式,在遗传学的发展中具有划时代的意义。
为此,F畅Jacob和J畅M onod获得了1965年诺贝尔奖。
14畅1畅3 建立乳糖操纵子模型的相关实验分析 关于细菌中酶诱导的现象早在20世纪初即已发现。
为什么细菌只有生长在合适的基质中才会有某种酶的产生?Jacob和M onod利用乳糖代谢系统进行了一系列研究,他们分离到一些Z、Y、A酶系统发生改变的大肠杆菌突变体。
其中有一类是结构基因本身的改变,如β半乳糖苷酶基因l acZ发生突变,由Z+突变为Z-,失去了合成β半乳糖苷酶的能力。
另外一类称为组成型突变体(constitu唱tive mutant),是指原来只有诱导物存在时才能进行酶合成的诱导性菌株,变成为没有诱导物时也能进行酶合成的突变型。
对这种恒定型突变株的遗传分析表明,这种突变多在I基因和O基因上,使野生型I+恒定地突变为I-,而O+则突变成O C。
还有一类称为超阻遏物突变体(superrep ressi on m utant),这种突变株丧失了所有合成结构基因产物的能力。
在获得一系列乳糖代谢突变体的基础上,运用大肠杆菌的有性杂交,可得到由染色体和F′因子组合的部分合子(m ero zygo te)。
这样,在受体细胞中除本身环状染色体外,还能通过质粒F因子带来供体细胞的相关基因,可用于观察其部分二倍体的行为。
Jacob和M onod根据大量实验对乳糖代谢中的相关基因进行了系统分析。
(1)调节基因I的功能及其产物的分析 从分离出的大肠杆菌乳糖代谢系统突变体进行遗传分析,Jacob和M onod用I-突变体菌株与野生型I+菌株进行比较,发现I+对结构基因l ac Z的调控来说都是显性,而在I-细胞中就不同,I-Z-/F I-Z+成了恒定型,I-总是隐性(表141中的第1~3号菌株)。
至于第4号菌株,显示其I+基因产物是反式作用,意味着其基因产物不论是顺式还是反式方式,都能够调节乳糖操纵子的结构基因。
表141 在单倍体和部分二倍体中β半乳糖苷酶、透性酶的合成β半乳糖苷酶透性酶菌株号基因型不经诱导诱导不经诱导诱导1I+Z+Y+-+-+2I-Z+Y+++++3I+Z-Y+/FI-Z+Y+-+-+4I-Z-Y+/FI+Z+Y--+-+329 续表菌株号基因型β半乳糖苷酶透性酶不经诱导诱导不经诱导诱导5I S Z +Y +----6I S Z +Y +/F I +Z +Y +---- 细菌生长在以甘油为碳源的培养基中,以IPTG 为诱导物。
“+”表示酶是高水平的,“-”表示酶是低水平或缺乏。
用另外一种突变型I S对I 基因功能的进一步研究表明,突变株丧失了合成所有结构基因产物的能力,在部分合子I S /F I +中,I S 是显性,如I S Z +Y +/F I +Z +Y +菌株在诱导物存在时既不合成β半乳糖苷酶,也不合成透性酶。
这些遗传学分析证明,I 基因是一个控制因子,但它的控制需要一个在细胞内扩散的组分来协助。
为什么在l ac I -突变菌株中阻遏物一失活,就不能与操纵基因相结合?为什么一个l ac I +菌株突变成l ac I S菌株后,合成酶的能力就全部丧失? 根据操纵子学说,上述现象显然都与调节基因I 的产物———阻遏物有关。
用14C 标记的诱导物异丙基βD 硫代半乳糖苷(isop ropyl βD thiogalactoside ,IPTG )作为一种检测剂,从大肠杆菌细胞抽提物中纯化阻遏物,然后再在体外将纯化的蛋白质与IPTG 进行结合实验。
IPTG 不仅与阻遏物有很强的结合能力,而且这种结合在抽提物中很稳定。
按照纯化蛋白质的方法收集与IPTG 结合的特异性抽提物,通过透析去除IPTG ,即可获得纯化的阻遏物。
但是,从带有I S 基因的菌株中分离得到的阻遏物,在体外实验中与IPTG 未显示亲和性。
IPTG 是β半乳糖苷酶的一种无关的诱导物,它不能被β半乳糖苷酶分解。
这种既能诱导酶的产生,而本身又不分解的诱导物称为义务诱导物(gra tuitous i n ducer )。
图142 诱导物结合引起阻遏物变构示意图(引自L ew i n ,2006)(a )头部片段结合到D N A 大沟的连续螺旋中 (b )诱导物结合,改变结合部位的D N A 构象,头部片段不能插入大沟中 对阻遏物其他的物理化学性质检测和X 射线晶体学分析表明,阻遏物是由4个相同亚基组成的同源四聚体。
每个亚基的相对分子质量约为38500,可分为几个功能域:N 端由两个被转角分开的α螺旋(H -T -H )组成,这两个α螺旋可插入到DN A 大沟中,因而是阻遏物的DN A 结合域。
该域由铰链与阻遏物的主体核心相连。
核心区可分成两个结构相同的区域(核心功能域1和2),各自具有夹板式结构,即在两边的α螺旋间夹有6列平行的β折叠片。
诱导物可结合在两核心区间的缝隙中。
C 端含有一个由两个亮氨酸七聚体重复的α螺旋,又称为寡聚体功能区,用于4个单体的聚合,维持阻遏物的四聚体结构(图142)。
对乳糖操纵子阻遏物结构分析表明,其四聚体实际上是由两个二聚体组成,二聚体各亚基含有明显的功能域,N 端的头部片段(head p i ece )是与DN A 结合的操纵子位点,而核心区则具有与诱导物结合的诱导物位点[图142(a )]。
因此,当二聚体各亚基的头部片段同时插入DN A 双螺旋的两个连续的大沟中时,与操纵基因区段紧密接触,大大增强了阻遏物和DN A 之间的亲和力,可阻断乳糖结构基因的表达。
然而,诱导物的结合却可引起阻遏物分子的变构,使头部片段的方向转而朝向核心部位弯曲,从而失去与操纵基因特异结合的能力[图142(b )]。