乳糖操纵子
- 格式:doc
- 大小:48.00 KB
- 文档页数:1
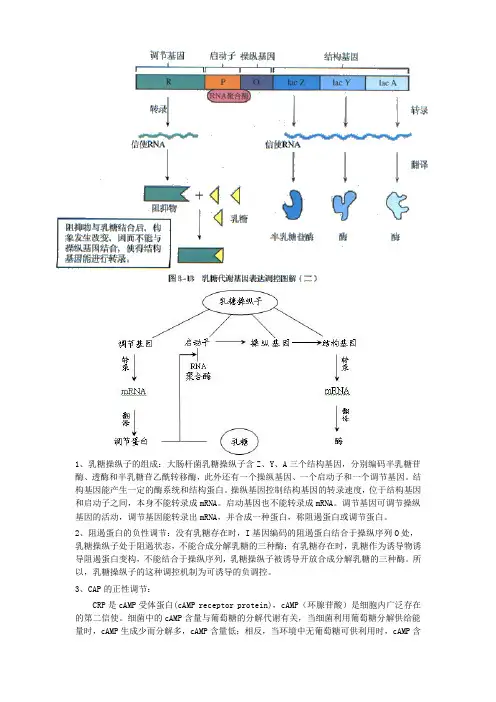
1、乳糖操纵子的组成:大肠杆菌乳糖操纵子含Z、Y、A三个结构基因,分别编码半乳糖苷酶、透酶和半乳糖苷乙酰转移酶,此外还有一个操纵基因、一个启动子和一个调节基因。
结构基因能产生一定的酶系统和结构蛋白。
操纵基因控制结构基因的转录速度,位于结构基因和启动子之间,本身不能转录成mRNA。
启动基因也不能转录成mRNA。
调节基因可调节操纵基因的活动,调节基因能转录出mRNA,并合成一种蛋白,称阻遏蛋白或调节蛋白。
2、阻遏蛋白的负性调节:没有乳糖存在时,I基因编码的阻遏蛋白结合于操纵序列O处,乳糖操纵子处于阻遏状态,不能合成分解乳糖的三种酶;有乳糖存在时,乳糖作为诱导物诱导阻遏蛋白变构,不能结合于操纵序列,乳糖操纵子被诱导开放合成分解乳糖的三种酶。
所以,乳糖操纵子的这种调控机制为可诱导的负调控。
3、CAP的正性调节:CRP是cAMP受体蛋白(cAMP receptor protein),cAMP(环腺苷酸)是细胞内广泛存在的第二信使。
细菌中的cAMP含量与葡萄糖的分解代谢有关,当细菌利用葡萄糖分解供给能量时,cAMP生成少而分解多,cAMP含量低;相反,当环境中无葡萄糖可供利用时,cAMP含量就升高。
cAMP浓度低,CRP未与cAMP结合,CRP不能被活化,当cAMP浓度升高时,CRP 与cAMP结合并发生空间构象的变化而活化,称为CAP(CRP-cAMP activated protein),能以二聚体的方式与特定的DNA序列结合。
CAP的通用名称是分解代谢基因激活蛋白(catabolic gene activator protein)。
在启动子上游有CAP结合位点(CAP binding site),当大肠杆菌从以葡萄糖为碳源的环境转变为以乳糖为碳源的环境时,cAMP浓度升高,与CAP结合,使CAP发生变构,CAP结合于乳糖操纵子启动序列附近的CAP结合位点,增强RNA聚合酶的转录活性,促进结构基因转录,调节蛋白结合于操纵子后促进结构基因的转录,对乳糖操纵子实行正调控,加速合成分解乳糖的三种酶。

乳糖操纵子复习题
乳糖操纵子是分子生物学中一个重要的概念,它涉及到基因表达的调
控机制。
以下是关于乳糖操纵子的复习题内容:
1. 定义:请简述乳糖操纵子是什么,并解释其在细胞中的作用。
2. 组成:描述乳糖操纵子的基本组成部分,包括启动子、操纵基因、
结构基因等。
3. 调控机制:解释乳糖操纵子的正调控和负调控机制是如何工作的。
4. 诱导:阐述乳糖如何作为诱导剂激活乳糖操纵子的表达。
5. 抑制:描述在没有乳糖的情况下,乳糖操纵子是如何被抑制的。
6. cAMP-CAP复合物:解释cAMP-CAP复合物在乳糖操纵子调控中的作用。
7. 乳糖操纵子的发现:简述乳糖操纵子是如何被发现的,以及这一发
现对分子生物学的意义。
8. 应用:讨论乳糖操纵子在现代生物技术中的应用,特别是在基因工
程和基因治疗中的作用。
9. 比较:将乳糖操纵子与其他类型的操纵子(如色氨酸操纵子)进行
比较,指出它们的异同点。
10. 实验研究:列举一些实验方法,用于研究乳糖操纵子的调控机制。
11. 问题解决:提出一些可能在研究乳糖操纵子时遇到的问题,并给出可能的解决方案。
12. 未来方向:探讨乳糖操纵子研究的未来方向,以及这些研究可能对医学和生物技术带来的影响。
通过这些问题的复习,可以加深对乳糖操纵子及其调控机制的理解,为进一步的学习和研究打下坚实的基础。

乳糖操纵子名词解释乳糖操纵子(lactose operation)能合成和分泌乳糖的一类重要细胞,它们分布在不同类型的细胞内。
乳糖操纵子中的酶系有的是糖苷酶,有的是羧酸酯酶,还有一些是复合酶。
目前已发现的操纵子有七种类型,但只有两个编码,不论是催化水解乳糖还是释放乳糖的酶均是如此。
乳糖操纵子分布在所有高等动物组织中,哺乳类有两种:insulin- like autoantibody-抗胰岛素样蛋白4;一种乳糖操纵子,由四个区域组成,分别编码降血糖蛋白4(hypoglycemic-like autoantibody-抗胰岛素样蛋白4),降血糖蛋白5(hypoglycemic-like albumin-抗胰淀粉样蛋白)和乳糖操纵子自身。
此外尚有由insulin- like autoantibody-抗胰岛素样蛋白4(抗-4)与血浆蛋白G、铁蛋白、转铁蛋白结合的复合体,即乳糖操纵子复合体(oligosaccharide-like autoantibody complex-球蛋白操纵子复合体),也可能存在于不同细胞。
其中抗-4分子量为50万,具有与抗-4同源的抗胰岛素样蛋白4抗原决定簇,不受胰岛素影响,当它和其它球蛋白合成后,会结合于巨噬细胞膜上,并被膜内的锌粒子中和,再与巨噬细胞内的受体结合,从而阻断胰岛素与受体结合,进入细胞内的胰岛素失去降血糖作用。
至今只发现一种能降低血糖的操纵子,此种操纵子也称为受体型操纵子。
此种操纵子是位于酪氨酸磷酸酶基因上游,酪氨酸激酶基因下游,有关的其他基因几乎均已克隆。
当激活后,位于上游的酪氨酸磷酸酶基因激活使细胞内游离的酪氨酸浓度增加,酪氨酸水解成磷酸肌醇和磷酸胆碱释放入血液循环中,这将使血糖降低;而酪氨酸磷酸酶则与血清白蛋白结合,阻止白蛋白转运氨基酸,抑制氨基酸通过白蛋白进入血液,也可以抑制外周组织对氨基酸的利用。
该操纵子中的受体称为“受体酪氨酸磷酸酶”。
该操纵子也可能参与葡萄糖和脂肪酸的代谢。


乳糖操纵子乳糖操纵子是参与乳糖分解的一个基因群,由乳糖系统的阻遏物和操纵序列组成,使得一组与乳糖代谢相关的基因受到同步的调控。
1961年雅各布(F.Jacob)和莫诺德(J.Monod)根据对该系统的研究而提出了著名的操纵子学说。
在大肠杆菌的乳糖系统操纵子中,β-半乳糖苷酶,半乳糖苷渗透酶,半乳糖苷转酰酶的结构基因以LacZ(z),Lac Y(y),Lac A(a)的顺序分别排列在染色体上,在z的上游有操纵序列Lac O(o),更前面有启动子Lac P(p),这就是操纵子(乳糖操纵子)的结构模式。
编码乳糖操纵系统中阻遏物的调节基因Lac I(i)位于和p上游的临近位置。
细菌相关功能的结构基因常连在一起,形成一个基因簇。
它们编码同一个代谢途径中的不同的酶。
一个基因簇受到同一的调控,一开俱开,一闭俱闭。
也就是说它们形成了一个被调控的单位,其它的相关功能的基因也包括在这个调控单位中,例如编码透过酶的基因,虽它的产物不直接参与催化代谢,但它可以使小分子底物转运到细胞中。
乳糖分解代谢相关的三个基因,lacZ、Y、A就是很典型的是上述基因簇。
它们的产物可催化乳糖的分解,产生葡萄糖和半乳糖。
它们具有顺式作用调节元件和与之对应的反式作用调节因子。
三个结构基因图的功能是:lacZ编码β-半乳糖苷酶(β-galactosidase),此酶由500kd的四聚体构成,它可以切断乳糖的半乳糖苷键,而产生半乳糖和葡萄糖lacY编码β一半乳糖苷透性酶(galactoside permease),这种酶是一种分子量为30kDd膜结合蛋白,它构成转运系统,将半乳糖苷运入到细胞中。
lacA编码β-硫代半乳糖苷转乙酰基酶(thiogalactosidetransacetylase),其功能只将乙酰-辅酶A上的乙酰基转移到β-半乳糖苷上。
无论是lacZ发生突变还是lacY发生突变却可以产生lac-型表型,这种lac-表型的细胞不能利用乳糖。

乳糖操纵子的原理
乳糖操纵子是真核生物基因中一段编码乳糖酶的 DNA序列。
该基因的发现,为研究在人体内存在的、与乳糖代谢有关的酶提供了线索。
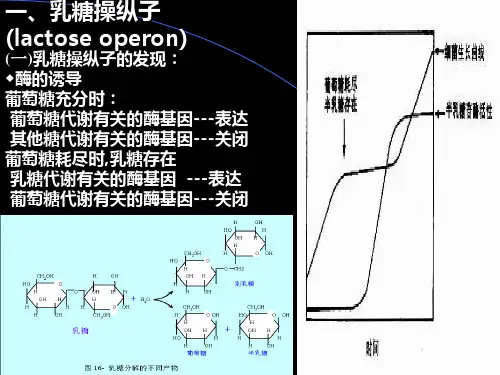
我们知道,牛奶是由各种不同类型的乳糖组成,即半乳糖和葡萄糖。
牛奶中除了这两种成分外,还含有其他一些成分,如钙、磷、铁等。
我们人体不能合成这些成分,必须由食物来补充。
乳糖是一种简单的碳水化合物,在人类和哺乳动物体内都存在。
人和动物食用牛奶后,小肠中的乳糖酶就会将其分解成葡萄糖和半乳糖。
在这一过程中,半乳糖苷被水解成单糖,进入大肠中与细菌产生的细菌素结合,细菌素进一步分解成酸和二氧化碳,经肠道排出体外。
这种由碳水化合物分解成单糖和二氧化碳的过程称为“分解代谢”。
乳糖是哺乳动物乳汁中最重要的碳水化合物成分之一。
在婴儿出生后3~6个月内,主要是靠母乳来提供能量。
在哺乳期内,由于母亲体内乳糖酶活力下降或缺乏,以及婴儿消化道尚未发育成熟等原因,母乳中的乳糖酶活性很低或缺乏。
—— 1 —1 —。


乳糖操纵子名词解释乳糖操纵子:乳糖操纵子也称为乳糖运输蛋白,是一种催化乳糖分解的酶。
表1乳糖操纵子名词解释 1。
催化酶。
由于其构成的蛋白质分子中有一个特殊序列(N),即所谓“不对称区”,在这个区域内有两种催化结构的酶同时起作用,因此对绝大多数天然底物有高度选择性。
乳糖是一种低聚糖,它与人体所需要的葡萄糖、半乳糖、阿拉伯糖等都属于单糖,在人体内具有特殊的功能,是糖类的主要成分之一,被吸收后参与新陈代谢。
因此,乳糖可作为食品添加剂应用到食品中去,作为提高人们生活水平的功能食品。
在食品工业上主要用来制造无糖糕点和饼干、奶粉、冰淇淋等,并将乳糖经发酵变成乳酸,以提高甜度,改善风味。
2。
乳糖操纵子。
存在于胞质溶胶内,又称胞质溶胶。
分布于质膜下,含有一个多肽复合物,由100多个氨基酸残基组成,不完全相同的数目超过10万个,该区域为不对称的α螺旋形结构。
3。
表位( mRNA)。
存在于胞质溶胶的不对称区,由RNA聚合酶在转录前的间隔期间转录。
表位是一种特殊的RNA,除了与操纵子有关外,还在RNA结构和进化中扮演着重要角色。
4。
基因家族。
位于基因簇,位于多个细胞中,组织中或系统中。
5。
编码区( coding region)。
基因组DNA上特定位置,在大多数哺乳动物和鸟类的基因中,在前驱物导入时,必须改变。
6。
操纵子( modulator)。
在一个基因簇上控制转录的一段DNA序列,有时可以是RNA。
这些序列有几个特征,例如与位于转录起始位置的操纵元结合,与酶有关联的氨基酸序列。
7。
基因簇( locus)。
在不同种或同种不同个体间,共享一段相同的核苷酸序列的核苷酸序列集合,包括基因。
8。
微卫星( Microsatellite)。
在染色体的DNA序列中,短的重复序列,常常出现的两个重复序列之间的间隙,是连续的DNA序列中的一段。
9。
小卫星( mini satellite)。
在染色体的DNA序列中,长的重复序列,常常出现的三个重复序列之间的间隙,是连续的DNA序列中的一段。

乳糖操纵子引言乳糖操纵子是一种能够操纵乳糖代谢的物质或机制的称谓。
乳糖是一种存在于奶制品中的碳水化合物,而对乳糖的消化和代谢需要人体内的乳糖酶来完成。
然而,有些人的体内缺乏乳糖酶,导致乳糖无法被消化和代谢,从而引发乳糖不耐受症状。
乳糖操纵子的出现为乳糖不耐受症患者提供了新的治疗和饮食选择。
本文将介绍乳糖操纵子的定义、分类以及应用等相关内容。
乳糖操纵子的定义乳糖操纵子是一种能够通过调节乳糖代谢的物质或机制。
乳糖操纵子可以分为两类:一类是能够增强乳糖酶活性的物质或机制,另一类是能够减弱或抑制乳糖酶活性的物质或机制。
通过作用于乳糖酶或其他相关代谢途径,乳糖操纵子可以改变人体对乳糖的代谢过程,从而影响乳糖不耐受症状的发生和程度。
乳糖操纵子的分类根据其对乳糖酶活性的影响,乳糖操纵子可以分为以下几类:1.增强型乳糖操纵子:这类乳糖操纵子能够增加乳糖酶的活性,提高乳糖的代谢速度。
常见的增强型乳糖操纵子包括乳糖酶替代物以及对乳糖酶具有促进作用的物质。
2.抑制型乳糖操纵子:这类乳糖操纵子能够减弱或抑制乳糖酶的活性,降低乳糖的代谢速度。
常见的抑制型乳糖操纵子包括乳糖酶抑制剂以及对乳糖酶具有抑制作用的物质。
乳糖操纵子的应用随着对乳糖不耐受症的研究深入,乳糖操纵子逐渐被应用在乳糖不耐受症的治疗和饮食调节中。
下面是乳糖操纵子在不同应用场景中的具体应用:1.药物治疗:某些乳糖操纵子可以作为药物用于治疗乳糖不耐受症。
例如,一些乳糖酶替代物可以在人体内代替缺乏的乳糖酶,帮助消化和代谢乳糖。
2.膳食调节:乳糖操纵子还可以通过膳食调节的方式应用于乳糖不耐受症的患者。
例如,饮食中可以添加含有增强型乳糖操纵子的食品,增加乳糖酶的活性,从而促进乳糖的代谢。
3.乳制品加工:乳糖操纵子的应用还可以扩展到乳制品的加工过程中。
例如,在酸奶的生产中可以添加抑制型乳糖操纵子,降低乳糖的含量,从而适应乳糖不耐受症患者的需求。
总结乳糖操纵子作为一种能够调节乳糖代谢的物质或机制,为乳糖不耐受症的患者提供了新的治疗和饮食选择。
乳糖操纵子名词解释乳糖操纵子( lac- G cluster),指能够对半乳糖等六种乳糖分子进行跨膜转运的转运体。
相关酶存在于高尔基体上的乳糖操纵基因( lac- G)调控亚单位(操纵子)内。
通常一个亚基由一个DNA 分子(操纵子)及若干个操纵蛋白(亚基)所组成,蛋白质为单顺反子。
当前的研究表明,乳糖操纵子可分为三个基本组件:操纵基因、蛋白转运亚基及转运蛋白。
在每一亚基中,均含有一对核酸( DNA或RNA)和蛋白质。
蛋白质的氨基酸序列和其相应的顺反子序列都是保守的,而与这些保守的氨基酸序列相互作用的核苷酸序列则因亚基而异。
操纵基因可以有若干个,通常是整合到基因组上;有些细菌的转运体可包括四个基因。
在不同的细菌间,同一乳糖操纵子也存在很大的差异,甚至同一菌属不同菌株间也存在着相当大的差异。
各种转运体之间有极其复杂的交叉现象,导致乳糖操纵子具有非常高的多样性。
乳糖操纵子的结构十分保守,对于鉴定遗传标记具有重要意义。
同一乳糖操纵子存在不同的亚基,其生理功能也有所差异。
一般来说,乳糖操纵子各亚基的基本功能是不同的,其主要区别有以下几点:( 1)转运亚基的形态和位置,取决于该亚基所携带的乳糖分子的大小和电荷。
此外,一个亚基往往含有两种或两种以上的乳糖分子,从而使该亚基的活性受到不同程度的影响。
( 2)转运蛋白的空间结构与转运性质。
转运蛋白以一种特定的方式结合在内质网上的特殊位点上,在转运过程中对于转运方向起重要作用。
人类及其它哺乳动物细胞中有多种转运体,这些转运体并不编码一种乳糖分子,但都有类似的结构,并且能将外源性乳糖分子从胞外转运至胞内。
不同细胞中乳糖操纵子的亚基数目可有很大差别,例如,分泌性胃肠道的Lac- G cluster包括一个编码前导蛋白的操纵基因,一个与受体蛋白结合的转运蛋白,还有两个负责ATP生成的转运蛋白亚基。
在乳糖操纵子中,有一部分的Lac- G亚基是不编码蛋白质的,这些亚基称为操纵基因。
乳糖操纵子的基本调控过程1.引言1.1 概述乳糖操纵子是一种广泛存在于真核生物中的基因调控系统,它在乳糖代谢过程中发挥重要作用。
乳糖是一种双糖,由葡萄糖和半乳糖组成,可被一些生物利用。
在某些条件下,乳糖操纵子可以调控与乳糖相关的基因的表达水平。
乳糖操纵子通常由两个组分组成:乳糖操纵子启动子和乳糖操纵子结合蛋白。
乳糖操纵子启动子位于被调控基因的上游区域,其中包含着与乳糖操纵子结合蛋白相互作用的DNA序列。
乳糖操纵子结合蛋白则通过与乳糖操纵子启动子结合,调控被调控基因的转录水平。
乳糖操纵子的基本调控过程涉及到乳糖操纵子结合蛋白的结合和解离,以及与乳糖的结合。
当环境中存在乳糖时,乳糖操纵子结合蛋白会与乳糖结合,形成复合物,进而与乳糖操纵子启动子结合。
这个复合物会招募其他转录因子和RNA聚合酶,促使被调控基因的转录。
而在乳糖缺乏的环境下,乳糖操纵子结合蛋白会解离,并不再与乳糖操纵子启动子结合,阻碍被调控基因的转录。
乳糖操纵子的基本调控过程对于真核生物的生存和生长具有重要意义。
它可以帮助真核生物适应环境中乳糖的变化,从而合理利用有限的营养资源。
同时,乳糖操纵子的基本调控过程也为我们研究基因调控的机制提供了一个良好的模型系统。
通过深入研究乳糖操纵子调控过程的细节,我们可以更好地理解基因的表达调控网络,为疾病的诊断和治疗提供新的途径。
在本文中,我们将详细讨论乳糖操纵子的基本调控过程的要点,以及其对真核生物的重要意义。
通过对其相关机制的探究,我们希望能够增进对基因调控的理解,并为进一步的研究提供启示。
同时,我们也将展望未来在这一领域的研究方向,为乳糖操纵子的研究和应用提供新的思路和方法。
1.2 文章结构文章结构部分的内容可以包括以下内容:文章按照以下结构组织:引言、正文和结论。
- 引言部分介绍了乳糖操纵子的研究背景和重要性,概述了本文要阐述的乳糖操纵子的基本调控过程,并简要介绍了文章的结构。
- 正文部分包括了乳糖操纵子的基本调控过程要点1和要点2的详细介绍。
乳糖操纵子的名词解释
乳糖操纵子是一种食物添加剂,是乳糖的一种合成代用品,通常用于替代糖分,改善口感和结构,降低糖分含量,尤其是低盐产品。
它的合成原理就是以甲醇(乙醇)为主要原料,加入不同类型的乙酸或醋酸钠,进行加氢反应,最后再经过糖化反应制成的糖分。
乳糖操纵子的类型有乳糖磷酸酯、乳糖酯类、乳糖酮类,以及乳糖衍生物多种多样,例如乳糖醛酸酯。
其中乳糖磷酸酯类是由乙酸磷酸和乙醇反应制成的,乳糖酯类则是由乳酸和乙醇反应生成的,乳糖酮类是由醋酸酯和乙醇反应生成的,乳糖衍生物则是由醋酸酯和乙醇反应生成的,最终形成不同糖分。
乳糖操纵子的特点是拥有新颖的口感和较低的热量,而且乳糖替代的热量比糖分更低,具有较强的抗氧化能力,从而降低糖分含量,从而帮助改善血糖控制,从而让使用者不受糖尿病影响。
乳糖操纵子目前应用范围很广,如果冰激凌、饮料和西点中都有乳糖操纵子的存在,点心面包和奶酪也非常常见,这是因为它们的口感更微妙,而乳糖操纵子可以改善食物的口感,并降低糖分含量,从而防止血糖过高。
此外,乳糖操纵子有助于改善血糖控制,可以帮助糖尿病患者预防并发症的发生,从而提高质量生活。
因此,乳糖操纵子可以被广泛应用于食品领域,它可以改善食物的口感,降低糖分,而且还有利于改善血糖控制。
尽管乳糖操纵子有许多优点,但我们仍然需要谨慎使用,让我们不要过度摄入。
我们可以选择更多的低糖和低盐的食物,以及经常进行体育锻炼,以增强我们的体质,减少糖尿病风险,更好的保护我们的健康。
乳糖操纵子结构一、引言二、乳糖操纵子结构的概念1. 乳糖的结构和功能2. 操纵子结构的定义和作用三、乳糖操纵子结构的种类及特点1. Lactose repressor (LacI)a. 结构特点b. 功能机制c. 应用领域2. Lactose permease (LacY)a. 结构特点b. 功能机制c. 应用领域3. β-galactosidase (LacZ)a. 结构特点b. 功能机制c. 应用领域四、乳糖操纵子结构在基因表达中的作用及应用前景1. 基因调控机制2. 生物技术应用五、总结与展望一、引言乳糖是一种重要的营养物质,广泛存在于动物奶类和某些植物中。
在细胞内,乳糖被代谢成葡萄糖和半乳糖,提供能量和碳源。
然而,在某些情况下,细胞需要对乳糖进行调控,以适应不同的环境条件。
这种调控过程主要依靠乳糖操纵子结构来实现。
本文将介绍乳糖操纵子结构的概念、种类及特点,以及在基因表达中的作用和应用前景。
二、乳糖操纵子结构的概念1. 乳糖的结构和功能乳糖是由葡萄糖和半乳糖组成的二糖,化学式为C12H22O11。
在细胞内,乳糖需要通过膜转运蛋白LacY进入细胞质,在细胞质中被β-galactosidase (LacZ)水解成葡萄糖和半乳糖。
2. 操纵子结构的定义和作用操纵子结构是指一类能够调节基因表达的DNA序列或蛋白质分子,它们可以与RNA聚合酶或其他转录因子相互作用,从而影响基因转录水平。
在乳糖代谢中,存在三种重要的操纵子结构:Lactose repressor(LacI)、Lactose permease (LacY)和β-galactosidase (LacZ)。
三、乳糖操纵子结构的种类及特点1. Lactose repressor (LacI)a. 结构特点LacI是一种DNA结合蛋白,由360个氨基酸残基组成。
它含有两个结构域:N-末端结构域和C-末端结构域。
N-末端结构域是一个立体互锁的四联体,包括两个相同的二聚体,每个二聚体都包括两个α螺旋和一个β折叠片。
一、简述乳糖操纵子的结构和诱导机制(英文)-(大题)
Functional and regulatory components of the lac operon(作用)
Lac R = Regulatory gene,that encodes for the lac Repressor protein that is concerned with regulating the synthesis of the structural genes in the operon. Lac R is adjacent to the Promoter site of the operon. The lac repressor is inactivated by lactose, and is active in the absence of lactose.
O = Operator,specific nucleotide sequence on DNA to which an active Repressor binds.
P = Promoter,specific nucleotide sequence on DNA to which RNA polymerase binds to initiate transcription. If the Repressor protein binds to the operator, RNAp is prevented from binding with the promoter and initiating transcription. Under these conditions the enzymes concerned with lactose utilization are not synthesized. Structual gene Lac Z, Y and A = Structural Genes in the lac operon. Lac Z encodes for Beta-galactosidase; Lac Y encodes the lactose permease; Lac A encodes a transacetylase.(lac = lactose),the inducer molecule. When lactose binds to the Repressor protein, the Repressor is inactivated; the operon is derepressed; the transcription of the genes for lactose utilization occurs.
Response to lactose(作用机制)
①Lack of inducer: the lac repressor binds to operator and blocks all. This prevents binding RNAp to promoter subsequent transcription of lac genes but a very low level of trans-cription of lacZYA .
②Lactose is present, the low basal level of permease allows its uptake, andβ-galactosidase catalyzes the conversion of some lactose to allolactose.
③Allolactose acts as an inducer, binding to the lac repressor and inactivate it. RNAp initiates transcription of lac structual genes.
二、Microbes are preferred to plants and animals as sources of enzymes because:(英文)1)they are generally cheaper to produce.
2)their enzyme contents are more predictable and controllable,
3)plant and animal tissues contain more potentially harmful materials than microbes, including phenolic compounds (from plants), endogenous enzyme inhibitors and proteases.
三、固定化酶的优点(Advantages of Immobilized Enzymes)(英文)
Immobilised enzymes are very important for commercial uses as they possess many benefits which include:
①Convenience: Minuscule amounts of protein dissolve in the reaction, so workup can be much easier. Upon
completion, reaction mixtures typically contain only solvent and reaction products.
②Economical: The immobilized enzyme is easily removed from the reaction making it easy to recycle the biocatalyst.
③Stability: Immobilized enzymes typically have greater thermal(热的)and operational stability than the soluble form of the enzyme.。