第七章 乳糖操纵子知识讲解
- 格式:ppt
- 大小:5.37 MB
- 文档页数:69


乳糖操纵子的结构和调控机制1. 引言乳糖操纵子是一种具有重要生理功能的DNA序列。
它在哺乳动物中起着调控乳糖代谢的关键作用。
本文将详细介绍乳糖操纵子的结构和调控机制,以及其在生物学中的重要性。
2. 乳糖操纵子的结构乳糖操纵子通常位于哺乳动物基因组中与乳糖代谢相关基因附近。
它是一个DNA序列,由一系列核苷酸组成。
根据不同物种和基因型的差异,乳糖操纵子可以具有不同长度和组成。
乳糖操纵子通常包含两个重要的元件:增强子和启动子。
增强子位于启动子上游,可以增加启动子活性,促进基因转录。
启动子位于基因上游,包含转录起始位点(TSS),是转录过程中RNA聚合酶与DNA结合的地点。
除了增强子和启动子,乳糖操纵子还可能包含其他调控元件,如转录因子结合位点和DNA甲基化位点。
这些元件的存在与特定物种和基因型相关,对乳糖操纵子的调控起到重要作用。
3. 乳糖操纵子的调控机制乳糖操纵子的调控机制涉及多个因素,包括转录因子、共激活子和染色质结构等。
下面将详细介绍几个重要的调控机制。
3.1 转录因子转录因子是乳糖操纵子调控的关键因素之一。
在乳腺细胞中,乳糖操纵子上的转录因子LacI结合到增强子上,阻止RNA聚合酶与启动子结合,从而抑制基因转录。
而在肝脏细胞中,另一种转录因子HNF-1α结合到增强子上,促进RNA聚合酶与启动子结合,增强基因转录。
3.2 共激活子共激活子是在乳糖操纵子调控过程中发挥重要作用的辅助蛋白质。
它们与转录因子一起结合到乳糖操纵子上,增强转录活性。
共激活子可以通过多种方式影响乳糖操纵子的调控,如改变染色质结构、招募其他转录因子等。
3.3 染色质结构染色质结构在乳糖操纵子调控中起着重要作用。
在非活化状态下,乳糖操纵子通常处于紧密的染色质状态,难以被转录因子和共激活子访问。
而在活化状态下,染色质会发生重塑,使得乳糖操纵子暴露在核内,便于转录因子和共激活子的结合。
4. 乳糖操纵子的生物学重要性乳糖操纵子在生物学中具有重要的功能和意义。


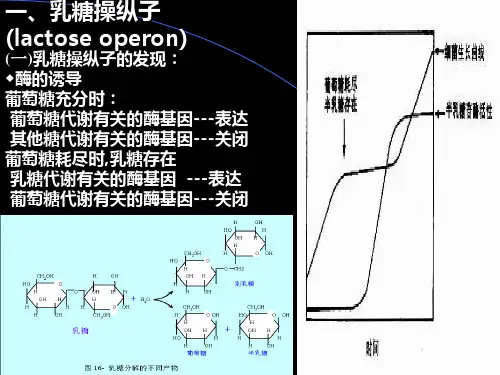
乳糖操纵子乳糖操纵子是参与乳糖分解的一个基因群,由乳糖系统的阻遏物和操纵序列组成,使得一组与乳糖代谢相关的基因受到同步的调控。
1961年雅各布(F.Jacob)和莫诺德(J.Monod)根据对该系统的研究而提出了著名的操纵子学说。
在大肠杆菌的乳糖系统操纵子中,β-半乳糖苷酶,半乳糖苷渗透酶,半乳糖苷转酰酶的结构基因以LacZ(z),Lac Y(y),Lac A(a)的顺序分别排列在染色体上,在z的上游有操纵序列Lac O(o),更前面有启动子Lac P(p),这就是操纵子(乳糖操纵子)的结构模式。
编码乳糖操纵系统中阻遏物的调节基因Lac I(i)位于和p上游的临近位置。
细菌相关功能的结构基因常连在一起,形成一个基因簇。
它们编码同一个代谢途径中的不同的酶。
一个基因簇受到同一的调控,一开俱开,一闭俱闭。
也就是说它们形成了一个被调控的单位,其它的相关功能的基因也包括在这个调控单位中,例如编码透过酶的基因,虽它的产物不直接参与催化代谢,但它可以使小分子底物转运到细胞中。
乳糖分解代谢相关的三个基因,lacZ、Y、A就是很典型的是上述基因簇。
它们的产物可催化乳糖的分解,产生葡萄糖和半乳糖。
它们具有顺式作用调节元件和与之对应的反式作用调节因子。
三个结构基因图的功能是:lacZ编码β-半乳糖苷酶(β-galactosidase),此酶由500kd的四聚体构成,它可以切断乳糖的半乳糖苷键,而产生半乳糖和葡萄糖lacY编码β一半乳糖苷透性酶(galactoside permease),这种酶是一种分子量为30kDd膜结合蛋白,它构成转运系统,将半乳糖苷运入到细胞中。
lacA编码β-硫代半乳糖苷转乙酰基酶(thiogalactosidetransacetylase),其功能只将乙酰-辅酶A上的乙酰基转移到β-半乳糖苷上。
无论是lacZ发生突变还是lacY发生突变却可以产生lac-型表型,这种lac-表型的细胞不能利用乳糖。

乳糖操纵子的结构和调控原理乳糖操纵子是一种广泛使用的基因表达调控工具。
它的结构和调控原理非常重要,对于科学家设计实验和开发新的应用具有重要意义。
乳糖操纵子是由两部分组成的:乳糖诱导子和乳糖操纵子蛋白。
乳糖诱导子是一种分子,能够与乳糖操纵子蛋白结合,从而改变其构象,使其能够与DNA结合并调控目标基因的表达。
乳糖操纵子蛋白则是一种转录因子,它能够识别并结合到特定的DNA序列上,并调节目标基因的转录水平。
乳糖操纵子蛋白通常被称为拉氏蛋白,因为它最初是从大肠杆菌中的λ噬菌体中分离出来的。
乳糖操纵子的调控原理是基于乳糖诱导子和乳糖操纵子蛋白之间的相互作用。
当乳糖存在时,乳糖诱导子能够与乳糖操纵子蛋白结合,使其构象发生改变,并使其与目标DNA序列结合。
这样,乳糖操纵子蛋白就能够调节目标基因的转录水平,以实现对基因表达的控制。
相反,当乳糖不存在时,乳糖诱导子无法与乳糖操纵子蛋白结合,从而使其无法与DNA结合。
这样,基因的转录就会被抑制。
乳糖操纵子的应用非常广泛,可以用于调控单个基因的表达,也可以用于调控整个基因组的表达。
其应用领域涵盖了基础科学研究、生物医学研究、生物工程、农业和环境保护等多个领域。
例如,在基础科学研究中,科学家可以利用乳糖操纵子来研究基因调控机制,进一步了解基因表达的调控途径。
在生物医学研究中,科学家可以利用乳糖操纵子来探究疾病的发生机制,同时也可以利用其来开发新的治疗方法。
在生物工程中,科学家可以利用乳糖操纵子来生产特定的蛋白质,例如,利用乳糖操纵子来生产重要的药物和酶类。
在农业和环境保护领域中,科学家可以利用乳糖操纵子来改良作物和微生物,提高其产量和抗病能力,同时也可以利用其来处理污染物和废弃物等。
乳糖操纵子的结构和调控原理是非常重要的,对于基因表达的控制和调节具有重要意义。
其应用前景广阔,对于推动生物科学的发展和应用具有重要作用。

乳糖操纵子原理乳糖操纵子(Lactose operon)是一种由基因组成的功能单元,它负责调控乳糖代谢的过程。
乳糖操纵子的研究为我们理解基因调控和代谢调控提供了重要的案例。
本文将介绍乳糖操纵子的组成和工作原理,并探讨乳糖操纵子在细菌的生长和适应性中的作用。
乳糖操纵子由三个关键元件组成:结构基因(structural genes)、调控基因(regulatory gene)和操作子(operator)。
结构基因包括三个相连的基因,分别编码乳糖酶(β-galactosidase)、内酰胺酶(permease)和转运蛋白(transacetylase)。
这些酶在细菌中起到催化乳糖分解的作用。
调控基因编码一个具有抑制功能的蛋白,称为乳糖抑制蛋白(lac repressor)。
操作子是一个DNA区域,位于结构基因和调控基因之间,它是乳糖操纵子的调控中心。
乳糖操纵子的工作原理是通过调控基因和操作子之间的相互作用实现的。
在没有乳糖的情况下,乳糖抑制蛋白结合在操作子上,阻止结构基因的转录和翻译,从而抑制乳糖酶、内酰胺酶和转运蛋白的合成。
当细菌环境中存在乳糖时,乳糖分子与乳糖抑制蛋白结合,导致乳糖抑制蛋白失活,从而使结构基因得以转录和翻译,合成乳糖酶、内酰胺酶和转运蛋白。
这些酶能够将乳糖分解为葡萄糖和半乳糖,进而参与细菌的能量代谢。
乳糖操纵子的调控机制是典型的负反馈调控。
当乳糖浓度较低时,乳糖抑制蛋白能够紧密结合在操作子上,有效地抑制结构基因的表达。
而当乳糖浓度增加时,乳糖分子与乳糖抑制蛋白结合,使其失活,从而解除对结构基因的抑制。
这种调控机制使得细菌能够根据环境中乳糖浓度的变化,灵活地调整乳糖代谢的速率,以适应不同的生长条件。
乳糖操纵子在细菌的生长和适应性中发挥着重要的作用。
乳糖是一种复杂的糖类,细菌需要通过乳糖酶等酶类将其分解为易于吸收利用的葡萄糖和半乳糖。
乳糖操纵子的存在使得细菌能够在环境中存在乳糖时快速合成乳糖酶等相关酶类,以满足细胞对乳糖的需求。



乳糖操纵子的结构和调控原理概述乳糖操纵子是存在于许多哺乳动物体内的一种能够调控乳糖代谢的关键分子。
它是由乳糖操纵子基因编码的蛋白质所组成,起着调控乳糖摄取和消化的重要作用。
本文将详细探讨乳糖操纵子的结构和调控原理。
乳糖操纵子的结构乳糖操纵子是一种单一的蛋白质,通常由若干不同的结构域组成。
根据其序列和结构的特点,乳糖操纵子可分为若干不同的亚型。
其中最为广泛研究的是LACZ、LACY和LACYD等。
LACZ亚型LACZ亚型是乳糖操纵子中最为常见的类型,它主要存在于大肠杆菌等细菌中。
LACZ亚型的乳糖操纵子通常是由若干结构域组成,包括信号肽、螺旋转位器、乳糖结合域和转运域等。
LACY亚型LACY亚型主要存在于大肠杆菌以外的一些细菌和真核生物中。
与LACZ亚型相比,LACY亚型的乳糖操纵子结构略有不同。
它包含了信号肽、乳糖结合域和转运域等主要结构域。
LACZD亚型LACZD亚型也是一种常见的乳糖操纵子亚型,主要存在于大肠杆菌中。
相较于LACZ 和LACY亚型,LACZD亚型的乳糖操纵子在结构上有一些差异,主要表现在转运域的结构上。
乳糖操纵子的调控原理乳糖操纵子的调控主要通过底物诱导和转录调控两种方式实现。
在底物诱导调控中,乳糖的存在会引起乳糖操纵子的构象改变,从而影响其功能。
而在转录调控中,一些转录因子会结合到乳糖操纵子的启动子区域,调节其转录活性。
底物诱导调控底物诱导调控是乳糖操纵子最常见的调控方式之一。
当乳糖存在于细胞外环境中时,它可以通过细胞膜上的乳糖操纵子结合域与乳糖操纵子进行结合。
这个结合过程会导致乳糖操纵子的构象改变,使得其转运功能得以激活。
乳糖操纵子的结构域之间存在着复杂的相互作用,转运态与非转运态之间的切换对乳糖摄取和代谢起着重要的调控作用。
转录调控除了底物诱导调控外,乳糖操纵子的转录也会受到一些转录因子的调控。
这些转录因子会结合到乳糖操纵子的启动子区域,调节其转录活性。
例如,在大肠杆菌中,CAP(catabolite gene activator protein)是一个重要的转录因子,它与RNA聚合酶结合,促进乳糖操纵子的转录。

原核生物基因表达转录水平调控之乳糖操纵子模型(2012-07-13 00:37:45)转载▼原核生物基因表达在转录水平上的调控最经典学说是操纵子学说。
一、操纵子细菌基因表达调控的许多原理是在研究E.coli乳糖代谢调节时被发现的。
法国巴斯德研究院的Francois Jacob与Jacques Monod于1960年在法国科学院院报(Proceeding of the French Academy of Sciences)上发表了一篇论文,提出乳糖代谢中的两个基因被一靠近它们的遗传因子所调节。
这二个基因为β半乳糖苷酶(β-galactosidase)和半乳糖苷透过酶(galactoside penmase)。
前者能水解乳糖成为半乳糖和葡萄糖,后者将乳糖运输到细胞之中。
在此文中他们首先提出了操纵子(operon)和操纵基因(operator)的概念,他们的操纵子学说(theory of operon)使我们得以从分子水平认识基因表达的调控,是一个划时代的突破,因此他们二人于1965年荣获诺贝尔生理学奖。
Jacob与Monod所提出的关于基因表达调控的操纵子学说可以简述如下:有一个专一的阻遏分子(蛋白质)结合在靠近β半乳糖苷酶基因上面,这段DNA他们称之为操纵基因。
由于阻遏分子结合在DNA的操纵基因上,从而阻止了RNA聚合酶合成β半乳糖苷酶的mRNA。
此外,他们还指出乳糖为诱导物,当乳糖结合到阻遏分子上时,即阻止阻遏分子与操纵基因的结合。
当有乳糖时,阻遏分子即失活,mRNA就可以转录出来。
如果去掉乳糖时,阻遏分子又恢复其活力,与操纵基因DNA结合,将乳糖基因关闭。
二、乳糖操纵子/fzswx/knowledge/knowledge01.asp?zsdBianhao=060302/s/blog_4b07ffbc01016v21.html乳糖操纵子(lac operon)是原核生物中研究得最清楚的一种操纵子。
乳糖操纵子简介操纵子(operon):很多功能相关的结构基因串联排列在染色体上,由一个共同的控制区来操纵这些基因的表达,包含这些结构基因和控制区的整个核苷酸序列就称为操纵子。
乳糖操纵子三个特异性序列:操纵序列O (operator): 阻遏蛋白结合位点。
启动子P (promoter): 位于结构基因的上游。
CAP结合位点:环cAMP受体蛋白(分解代谢物激活蛋白)结合位点。
一个调节基因●lac I:编码阻遏蛋白,能结合于操纵序列位点。
操纵子的组成:----结构基因(structural gene, SG) :操纵元中被调控的编码蛋白质的基因----启动子(promoter,P):是指能被RNA聚合酶识别、结合并启动基因转录的一段DNA序列。
----操纵基因(operator,O):是指能被调控蛋白特异性结合的一段DNA序列。
阻遏物基因(inhibitor,I),产生阻遏物(repressor)。
结构基因Z编码β-半乳糖苷酶:将乳糖水解成葡萄糖和半乳糖。
Y编码β-半乳糖苷透过酶:使外界的β-半乳糖苷(如乳糖)能透过大肠杆菌细胞壁和原生质膜进入细胞内。
A编码β-半乳糖苷乙酰基转移酶:乙酰辅酶A上的乙酰基转到β-半乳糖苷上,形成乙酰半乳糖。
当一个mRNA含有编码一个以上蛋白质的编码信息,而且这些蛋白质都是以独立的多肽被翻译时,这样的mRNA称之多顺反子mRNA。
多顺反子mRNA在细菌中是很普遍的。
多顺反子lac mRNA中的lacZ,lacY,lacA经翻译生成的产物分别生成代谢分解乳糖的三种酶始终存在着一定的比例关系( Z : Y : A = 5 : 2 : 1 )lacZ、Y、A基因的转录是由lacI基因指令合成的阻遏蛋白R所控制。
lacI一般和结构基因相毗连,但它本身具有自己的启动子和终止子,成为独立的转录单位。
由于lacI的产物是可溶性蛋白,按照理说是无需位于结构基因的附近。
它是能够分散到各处或结合到分散的DNA位点上。
乳糖操纵子的工作原理
嘿,朋友们!今天咱们来聊聊乳糖操纵子的工作原理,这可太有意思啦!
想象一下,乳糖操纵子就像是一个超厉害的团队在运作。
其中有个关键
角色,那就是调节基因。
它就好比是这个团队的总指挥,能产生一种叫阻遏蛋白的东西。
咱就说,如果没有乳糖的时候,阻遏蛋白就像个严格的守卫,紧紧地挡在乳糖操纵子的门口,不让相关基因表达,这厉害不厉害?好比没有任务的时候,不让大家随便行动。
但一旦有乳糖出现了呢!嘿,这情况可就不同啦。
乳糖会变成另一种东
西叫异乳糖,这异乳糖就像是一把钥匙,能和阻遏蛋白结合,让阻遏蛋白变形,就不再能好好拦住门口啦!于是,相关基因就可以开始表达啦。
这就好像有了紧急任务,那自然就得让大家行动起来呀,是不是很神奇?“哎呀,这也太有趣了吧!”
然后呢,这些基因表达出来的产物,像β-半乳糖苷酶等,就可以去分解乳糖,为细胞提供能量啥的。
这就像是团队成员们开始干活啦,忙得不亦乐乎。
比如说,β-半乳糖苷酶就像个专业的清洁工,把乳糖打扫得干干净净。
再想想,这和咱们的生活中很多情况是不是挺像的呀?就好比学校里有个活动,没有开始的时候大家都等着,一旦信号来了,大家就各就各位开始动起来啦。
总之,乳糖操纵子的工作原理就是这么奇妙!它能根据环境中的乳糖情况,精准地调控基因的表达,确保细胞能有效地利用资源,真的是太精妙啦!这就是我对乳糖操纵子工作原理的理解,你们觉得是不是这样呢?。
[复习]乳糖操纵子1、乳糖操纵子的组成:大肠杆菌乳糖操纵子含Z、Y、A三个结构基因,分别编码半乳糖苷酶、透酶和半乳糖苷乙酰转移酶,此外还有一个操纵基因、一个启动子和一个调节基因。
结构基因能产生一定的酶系统和结构蛋白。
操纵基因控制结构基因的转录速度,位于结构基因和启动子之间,本身不能转录成mRNA。
启动基因也不能转录成mRNA。
调节基因可调节操纵基因的活动,调节基因能转录出mRNA,并合成一种蛋白,称阻遏蛋白或调节蛋白。
2、阻遏蛋白的负性调节:没有乳糖存在时,I基因编码的阻遏蛋白结合于操纵序列O处,乳糖操纵子处于阻遏状态,不能合成分解乳糖的三种酶;有乳糖存在时,乳糖作为诱导物诱导阻遏蛋白变构,不能结合于操纵序列,乳糖操纵子被诱导开放合成分解乳糖的三种酶。
所以,乳糖操纵子的这种调控机制为可诱导的负调控。
3、CAP的正性调节:CRP是cAMP受体蛋白(cAMP receptor protein),cAMP(环腺苷酸)是细胞内广泛存在的第二信使。
细菌中的cAMP含量与葡萄糖的分解代谢有关,当细菌利用葡萄糖分解供给能量时,cAMP生成少而分解多,cAMP含量低;相反,当环境中无葡萄糖可供利用时,cAMP含量就升高。
cAMP浓度低,CRP未与cAMP结合,CRP不能被活化,当cAMP浓度升高时,CRP与cAMP结合并发生空间构象的变化而活化,称为CAP(CRP-cAMP activated protein),能以二聚体的方式与特定的DNA序列结合。
CAP的通用名称是分解代谢基因激活蛋白(catabolic gene activator protein)。
在启动子上游有CAP结合位点(CAP binding site),当大肠杆菌从以葡萄糖为碳源的环境转变为以乳糖为碳源的环境时,cAMP浓度升高,与CAP结合,使CAP发生变构,CAP结合于乳糖操纵子启动序列附近的CAP结合位点,增强RNA 聚合酶的转录活性,促进结构基因转录,调节蛋白结合于操纵子后促进结构基因的转录,对乳糖操纵子实行正调控,加速合成分解乳糖的三种酶。