反转录病毒及反转录转座子
- 格式:pdf
- 大小:693.30 KB
- 文档页数:17



原核生物转座子类型
原核生物的转座子主要有以下几类:
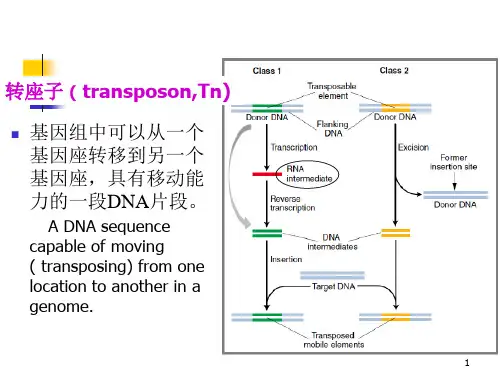
1.插入序列(Insertional Sequence,IS):两端有IR,只编码转座酶。
2.类转座因子(类转座子):结构同IS但不能独立存在,只作为复合子的两端组件。
3.复合转座子:两端由IS或类IS构成,带有某些抗药性基因或其他宿主基因,一旦形成复合式转座子,IS序列就不能再单独移动,只能作为复合体移动。
4.TnA转座子家族:两端为IR可编码转座酶、解离酶和抗性物质。
此外,真核生物的转座子主要包括转座子和反转录转座子两类。
具体分类如下:
1.转座子:包含两类。
a.玉米内的可控制因子,可分为自主性因子和非自主性因子,如Ac-Ds体系。
b.果蝇中的转座子,如P转座子,导致杂种不育。
2.反转录转座子:包含三类。
a.反转录病毒RNA:整合宿主靶DNA。
b.病毒超家族:有LTR,编码反转录或整合酶,可含内含子。
如Ty Copia LINSL1。
c.非病毒超家族:无重复序列,不编码转座产物、无内含子。
如SINSB1/ALU、假基因。
以上信息仅供参考,建议查阅关于原核生物转座子的文献资料或咨询相关生物学家,获取更准确的信息。


【⽂章知识点】深度解析长末端重复反转录转座⼦(LTR-RTs)提起 LTR,相信很多⼈和我之前⼀样都是熟悉⼜陌⽣的感觉,听过或者接触过却未深⼊了解过。
若您对 LTR 分析有兴趣,却苦于⽆从下⼿时,愿本⽂作为⼀个叩门砖,为您敲开 LTR 分析的⼤门。
本篇从 LTR 的定义、分类、⽣物学意义、结构特征、鉴定⽅法等⽅⾯层层递进,带您⾛进神奇的 LTR 世界。
1. LTR 与重复序列、转座⼦的关系LTR-RTs 是 Long terminal repeat-retrotransposons 的缩写,中⽂名是长末端重复反转座⼦。
LTR-RTs 名字中既有重复、⼜有转座⼦,那么它和重复序列、转座⼦是什么关系呢?图1 为您解答。
图1 重复序列主要分类重复序列:根据重复区域是否连续可分为串联重复序列和散在重复序列(⼜名转座⼦、转座元件)两⼤类,前者相连,后者不相连。
转座元件(transposable elements, TEs) ⼜称转座⼦:指在基因组中能够移动或复制,并可以整合到基因组新位点的⼀段 DNA 序列。
根据转座过程是否形成 RNA 中间体,转座⼦可分为 DNA 转座⼦和反转录转座⼦。
反转录转座⼦是以 RNA 为媒介,伴有反转录过程,以复制-粘贴的⽅式在基因组的新位置产⽣⼀个新的拷贝。
DNA 转座⼦的转座机制则是剪切-粘贴的形式。
LTR-RTs :是反转座⼦中的⼀种,因其两侧存在长的末端重复⽽得名。
不含长末端重复的反转座⼦统称 non-LTR-RTs,主要包含短散在重复(SINE)和长散在重复(LINE)。
2. LTR的分类动植物基因组中存在⼤量转座⼦,尤其是植物基因组中。
LTR 因其数量多且 LTR 长度巨⼤,在植物转座⼦中具有较⾼的基因组含量。
在⽟⽶基因组中 LTR 占基因组含量⾼达 75% ,⼭苍⼦基因组中 LTR 占⽐⾼达 47%,所以基因组 LTR 的鉴定尤为重要。
反转录转座⼦根据转座元件结构的完整性和转座特点可分为⾃主元件(编码转座酶)和⾮⾃主元件(⾃⾝不编码转座酶)。


东北师范大学2020—2021学年第1学期
《现代分子生物学》考试试卷(A卷)
院/系年级专业姓名学号
考生答题须知
1.所有题目答题答案必须做在考点发给的答题纸上,做在本试题册上无效。
请考生务必在答题纸上写清题号。
2.评卷时不评阅本试题册,答题如有做在本试题册上而影响成绩的,后果由考生自己负责。
3.答题时一律使用蓝、黑色墨水笔或圆珠笔作答(画图可用铅笔),用其它笔答题不给分。
4.答题时不准使用涂改液等具有明显标记的涂改用品。
东北师范大学2020—2021学年第1学期《现代分子生物学》考试试卷(A卷)
标准答案。

逆转录原理
逆转录,又称逆转录转座子,是一种DNA复制的遗传工具,其分子机制和DNA复制类似,但在实际操作过程中,所需的酶比DNA复制所需的酶要多。
RNA反转录是将RNA反转录成cDNA的过程,在这个过程中,RNA由一个核糖核酸分子聚合成双链DNA分子。
cDNA是逆转录酶把cDNA合成出来的“原材料”。
逆转录酶能催化RNA的合成,使cDNA分子的4种核苷酸聚合成双链。
当病毒感染细胞时,细胞膜上的受体蛋白(通常是病毒和宿主细胞的特异性受体)通过与病毒的结合而进入细胞内。
病毒DNA与受体蛋白结合后,形成互补链,再与相应的RNA结合形成双链。
双链通过内吞作用进入细胞内,然后进入细胞核内进行转录和复制。
由于双链上各碱基之间有一定比例(如腺嘌呤与胸腺嘧啶之比为1∶1),RNA在进入细胞核后形成两条单链RNA。
当逆转录酶把一条单链RNA降解成一段双链RNA后,就可以进行翻译了。
在这个过程中需要许多酶参与。
—— 1 —1 —。


植物反转录转座子及其分子标记王子成1,2李忠爱2邓秀新1(1华中农业大学作物遗传改良国家重点实验室,湖北武汉,4300702 河南大学生命科学学院,河南开封,475001)摘要:反转录转座子(retrotransposon)是真核生物中一类可移动因子,可分为LTR反转录转座子和非LTR反转录转座子。
反转录转座子以高拷贝在植物界广泛分布,可以通过纵向和横向分别在世代之间和不同种之间进行传递,同一家族的反转录转座子具有高度的异质性. 在一些生物的和非生物的逆境条件下,反转录转座子的转录可以被激活。
由于反转录转座子的特点,使其作为一种分子标记得以应用。
S-SAP,IRAP,REMAP和RBIP等分子标记相继发展起来,在基因作图、生物遗传多样性与系统进化、品种鉴定等方面具有广泛的应用前景。
关键词反转录转座子,分子标记Plant retrotransposons and their molecular markersWang Zicheng1,2Li Zhongai2Deng Xuixin11 National Key Laboratory of Crop Genetic Improvement, Huazhong Agriculture university HubeiWuhan, 4300702 College of life science ,Henan University, Henan Kaifeng, 475001Abstract: Retrotransposons are a class of eukaryotic transposable elements, consisting of the long terminal repeat (LTR) and non-LTRretrotransposons. Retrotransposons are ubiquitous in the plant kingdom by high copy number and can be transmitted between generations by vertical transmission and between species by horizontal transmission. The same family retrotransposons presented highly heterogeneous populations in all higher plant genomes. Many of the plant retrotransposons are transcriptionally activated by various biotic and abiotic stress factors. Retrotransposons are used as molecular markers for their traits. S-SAP, IRAP, REMAP and RBIP are developed and will be applied widely in gene mapping, genetic biodiversity and phylogeny studies, and cultivar certification.Key words: retrotransposons molecular markers反转录转座子是广泛分布于真核生物中的一类可移动因子,因其转座需经过由RNA介导的反转录过程而得名。
病毒的繁殖方式病毒的繁殖方式是指病毒如何在寄主细胞中进行复制和传播的过程。
病毒的繁殖过程可以分为吸附、侵入、复制、组装和释放等阶段。
不同类型的病毒有不同的繁殖方式,下面将分别介绍几种常见的病毒繁殖方式。
1. 吸附:病毒通过表面的受体与寄主细胞表面的配体结合,实现吸附。
这个过程类似于磁力吸附的原理,病毒依靠与细胞表面的特定受体结合,使病毒固定在宿主细胞上。
2. 侵入:病毒侵入是指病毒将其遗传物质植入寄主细胞的过程。
大多数病毒进入细胞是通过寄主细胞表面的特定受体介导的。
病毒在侵入过程中,会通过改变细胞膜的结构或引发细胞内信号传导等方式,使得病毒能够穿过细胞膜,进入细胞质内。
3. 复制:病毒侵入了寄主细胞后,会开始进行复制。
病毒的复制方式主要有两种:非生产破裂和生产破裂。
非生产破裂是指病毒将其遗传物质直接复制并在细胞内积累,而生产破裂则是指病毒将其遗传物质复制成新的病毒颗粒,然后破坏宿主细胞释放出来。
复制过程中,病毒会利用宿主细胞的资源合成蛋白质和核酸,从而实现自身的复制。
4. 组装:在复制完成后,病毒会将新合成的病毒颗粒组装起来。
病毒组装过程中,病毒的蛋白质和核酸会自发地结合起来,形成完整的病毒颗粒。
5. 释放:组装完成后,病毒会寻找适当的机会离开宿主细胞。
病毒的释放方式主要有两种:溶解和膜融合。
溶解是指病毒在细胞内积累到一定数量后,使细胞溶解从而释放出来;膜融合是指病毒将其外膜与细胞膜融合,使病毒释放出来。
除了上述常见的病毒繁殖方式外,还有一些特殊的病毒繁殖方式。
1. 转座子:转座子是一种具有轉位酶活性的短片段DNA 或RNA分子,它可以在基因组中不同位置的DNA或RNA之间移动。
这种病毒可以通过改变其基因组的位置,导致寄主细胞的突变。
2. 寄生反转录病毒:寄生反转录病毒是一种具有反转录酶活性的病毒,它可以将自己的RNA转录成DNA,然后将其整合到寄主细胞的基因组中。
这种病毒可以通过改变寄主基因组的结构和功能,导致细胞突变。
逆转录转座子名词解释逆转录转座子是一类存在于真核生物基因组中的DNA序列,具有能够在基因组内移动、复制和插入至新位置的能力。
这种移动过程是通过逆转录酶进行的,即先将逆转录酶使用RNA短片(也称为逆向转录)转录DNA,然后将该DNA插入到某个新的基因组位置。
逆转录转座子在基因组中的存在广泛而普遍。
它们构成了基因组的一大部分,并且在物种间和个体内都显示出高度多样性。
逆转录转座子通过在基因组内移动和插入等机制,对基因组的结构和功能产生重要影响。
逆转录转座子的命名通常基于它们的结构、序列和机制特点。
以下是一些常用的逆转录转座子名词解释:1. 逆转录酶(Reverse transcriptase):逆转录酶是逆转录转座子中的一个关键酶。
它能够将RNA序列转录成DNA序列,并通过与Host DNA发生杂交,使其插入到基因组的新位置。
逆转录酶被广泛研究,也是研究逆转录转座子机制的重要工具。
2. 逆转录转座(Retrotransposition):逆转录转座是逆转录转座子的核心过程,即将逆向转录得到的DNA插入到基因组中的新位置。
逆转录转座可以在同一个个体内的不同位置之间移动,也可以在不同个体、不同物种之间传递。
3. 长转座子(Long interspersed element,LINE):长转座子是一类具有逆转录转座能力、较长的DNA序列。
它们通常具有反转录酶和核酸酶活性,能够自主复制和插入到基因组的新位置。
LINEs 在基因组中占据很大比例,它们通过多种机制对基因组进行重塑和转录调控。
4. 短转座子(Short interspersed element,SINE):短转座子是一类较短的逆转录转座子,通常长度为100-500个碱基对。
它们依赖于其他转座子或逆转录酶的插入和转录活动。
短转座子在物种间高度变异,可能对基因组中的基因表达和调控起重要作用。
5. 宿主基因组(Host genome):宿主基因组指逆转录转座子所插入的基因组,也就是它们的承载环境。