趋化因子研究进展ppt课件
- 格式:ppt
- 大小:3.80 MB
- 文档页数:48

全国趋化因子理论与实验学习班参考资料北京大学人类疾病基因研究中心2002年8月18-19日细胞因子总论1.导言在免疫系统中,各种免疫细胞均能合成和分泌小分子的多肽类因子,它们调节机体的免疫调节功能,参与免疫细胞的增殖,分化和行使功能,这些因子统称为细胞因子(cytokine)。
细胞因子除存在于免疫系统外,在机体的各个系统也广泛存在,发挥极为重要的生理调节作用,某些情况下可产生病理作用。
与免疫有关的细胞因子主要包括淋巴细胞产生的淋巴因子、单核巨噬细胞产生的单核因子、白细胞介素(interleukin,IL)、干扰素(interferon,IFN)、集落刺激因子(colony stimulating factor,CSF)、肿瘤坏死因子(tumor necrosis factor,TNF)、趋化因子(chemokine)、转化生长因子β(transforming growth factor β,TGFβ)等,它们在免疫系统中起着非常重要的调控作用,在异常情况下也会导致免疫病理反应。
1.1 研究历史1957年,Issacs等人发现病毒感染的细胞产生一种因子, 可抵抗病毒的感染, 干扰病毒的复制, 因而命名为干扰素,这是发现第一个细胞因子。
但以后很长时间内,对细胞因子的了解只限于发现一些细胞培养上清液具有各种生物学活性,例如,分裂素或抗原刺激的T淋巴细胞培养上清含有T细胞生长因子(T cell growth factor,TCGF)、B细胞生长因子(B cell growth factor,BCGF)、T细胞替代因子(T cell replacing factor)活性;脂多糖(LPS)刺激的单核巨噬细胞可产生淋巴细胞活化因子(lymphocyte activating factor,LAF)、肿瘤坏死因子活性等,这主要是由于这些细胞因子的含量非常低, 并且多种细胞因子同时存在, 纯化困难, 因而限制了对其进行结构和功能的研究。

趋化因子CXCL12又称基质细胞衍生因子1(SDF-1)—是小分子的细胞因子,属于趋化因子蛋白家族趋化因子CX CL12又称基质细胞衍生因子1(SDF-1)—是小分子的细胞因子,属于趋化因子蛋白家族。
它有两种形式,基质细胞衍生因子1α/CX CL12a和基质细胞衍生因子1β/CX CL12b。
趋化因子有4个保守的半胱氨酸残基形成两对双硫键以构成趋化因子的特殊结构。
第一第二半胱氨酸残基之间隔着一个介入氨基酸残基。
学术术语来源---内源性骨髓间充质干细胞与骨折微环境中的相关趋化因子文章亮点:1随着时间推移,骨折部位绿色荧光蛋白阳性细胞所占比例逐渐增高,增高的速率逐渐放缓,说明骨髓内的骨髓间充质干细胞参与骨折修复并在其中起到了重要作用。
2 基质细胞衍生因子1在骨折微环境中的表达亦呈先快速增高而后趋于平稳的趋势,初步证实了基质细胞衍生因子1在骨折微环境中的表达与骨髓间充质干细胞趋化到骨折部位之间存在关联。
3 骨折微环境中,肝细胞生长因子亦有一定量的表达,而且其表达亦呈逐渐增高的趋势。
这可能有利于上调骨髓间充质干细胞相应受体c-Met,进而诱导骨髓间充质干细胞的迁移。
4 骨折早期微环境中,单核细胞趋化蛋白1的表达量亦不小,它的表达可以招募单核巨噬细胞,参与炎症反应,似乎亦可以直接或间接地对骨髓间充质干细胞的趋化起到一定作用。
关键词:干细胞;骨髓干细胞;绿色荧光蛋白;骨髓间充质干细胞;嵌合体;骨折;趋化因子;基质细胞衍生因子1;集落刺激因子;肝细胞生长因子;单核细胞趋化蛋白1;间质金属蛋白酶9;省级基金;干细胞图片文章摘要背景:研究表明,骨髓间充质干细胞定向迁移依赖于损伤局部表达趋化因子与细胞表面相应受体的相互作用。

趋化因子CXCL10在恶性肿瘤发生、发展中的研究进展许锦辉【期刊名称】《重庆医学》【年(卷),期】2016(045)013【总页数】3页(P1849-1851)【关键词】趋化因子;CXCL10;IP10;恶性肿瘤;发生;发展【作者】许锦辉【作者单位】西南医科大学附属医院肿瘤科,四川泸州646000【正文语种】中文【中图分类】R73-3趋化因子是一种相对分子质量小的结构相关性蛋白,它通过与靶细胞膜上相应的受体结合引起靶细胞的定向迁移。
CXC趋化因子配体10(chemokine ligand-10,CXCL10)又称干扰素诱导蛋白(interferon-γ-inducible protein 10,IP-10),是CXC趋化因子超家族中的一种,具有调节免疫应答、血管生成、细胞凋亡、细胞周期以及细胞增殖等作用。
CXCL10与多种肿瘤的发生、发展、治疗疗效、预后均有相关性[1-2]。
研究发现CXCL10对肿瘤有抑制和促进的双重作用,其机制仍然不完全清楚。
本文就CXCL10在肿瘤发生发展中的作用进行综述。
趋化因子是一类小分子蛋白,相对分子质量为(8~10)×103,属于细胞因子超家族中的一大类。
基于在趋化因子分子N端的头两个半胱氨酸残基的不同位置,可以把趋化因子分成两类4个亚型:主要亚型(CX3C和CXC)和次要亚型(CC和C)[3]。
CXCL10是1985年Luster等[4]在研究干扰素-γ(IFN-γ)诱发的免疫应答时从U937细胞中克隆得到,因其N端的2个半胱氨基酸残基被一个非保守性氨基酸残基分割,把它归类于CXC趋化因子。
由于它具有炎性趋化作用,因此它也被命名为炎症趋化因子。
此外,因为它具有抑制血管的作用,因此也被称为做抗血管生成趋化因子。
它对肿瘤具有促进及抑制的双重作用,这可能与其本身的变体结构、受体的亚型、趋化的细胞种类及其靶细胞的种类有关系。
CXCL10主要通过与趋化因子受体3 (chemokine receptor 3,CXCR3)结合发挥生物学效应,CXCR3有两种变体结构:CXCR3-B以及CXCR3-alt,这两种变应体都是通过选择性剪接编码原始的CXCR3受体(从此被称为CXCR3-A)mRNA产生[5-6]。
趋化因子(chemokines):是指具有吸引白细胞移行到感染部位的一些低分子量趋化因子(多为8-10KD)的蛋白质(如IL-8、MCP-1等),在炎症反应中具有重要作用。
趋化因子Chemokine激发白细胞趋化性的小分子分泌性蛋白质。
是可受化学诱导物及细胞因子调节、并能刺激细胞趋化运动的一类细胞因子。
依照保守的半胱氨酸残基,可分为:①α趋化因子,含有保守的半胱-X-半胱(C-X-C)模体,主要是激发中性粒细胞趋化性;②β趋化因子,含有相邻的半胱氨酸残基(C-C),主要吸附单核细胞、嗜酸性粒细胞、嗜碱性粒细胞;③γ趋化因子,仅含有一对半胱氨酸残基,激发淋巴细胞趋化性(即淋巴细胞趋化因子);④δ趋化因子,含有一个半胱-X-X-X-半胱(C-X-X-X-C)模体,其作用仅局限于脑部,并被锚定在膜上。
所有受体都是G蛋白偶联受体。
简介人体在防御和清除入侵病原体等异物时,有一种使白细胞趋集的功能,有一些物质能引起这种功能称之为趋化剂或趋化因子,IL-8同时也是一种趋化因子。
趋化因子也称做趋化激素、趋化素或是化学激素。
是一小分子细胞因子家族蛋白。
趋化因子蛋白的共同结构特征包括,分子量小(约8-10 千道尔顿),有四个位置保守的半胱氨酸残基以保证其三级结构。
这些小蛋白因其有定向细胞趋化作用而得名。
当然,这些蛋白有些趋化因子历史上还有其他的名字, 包括已知的SIS细胞因子家族、 SIG细胞因子家族, SYC细胞因子家族和血小板因子-4家族。
有的趋化因子被认为促进炎症反应,而有些趋化因子被认为在正常的修复过程或发育中控制细胞的迁徙。
在所有脊椎动物和一些病毒和一些细菌中有趋化因子存在, 但不存在于其他无脊椎动物. 这些蛋白质结合到趋化因子受体而起作用,趋化因子受体是G蛋白偶连的跨膜受体,选择性地表达在靶细胞表面。
编辑本段功能趋化因子的主要作用是趋化细胞的迁移. 细胞沿着趋化因子浓度增加的信号向趋化因子源处的迁徙。
有些趋化因子在免疫监视过程中控制免疫细胞趋化,如诱导淋巴细胞到淋巴结。

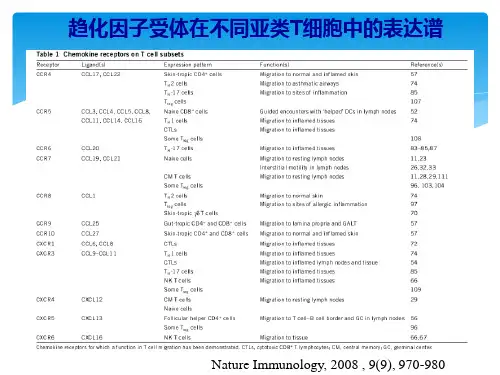
趋化因子受体起始期(80s中)未形成概念PF4成熟期(80s后)完整概念及分类大量趋化因子高潮期(90s中)发现CCR5 新趋化因子及受体趋化因子(chemokines):是一类由细胞分泌的小细胞因子或信号蛋白,由于它们具有诱导附近反应细胞定向趋化的能力,因而命名为趋化细胞因子,免疫细胞的定向迁移是机体免疫应答发生和完成的必须条件,趋化因子是一类控制细胞定向迁移的细胞因子。
趋化因子受体的简介:是一类介导趋化因子行使功能的GTP-蛋白偶连的跨膜受体(GPCR),通常表达于免疫细胞、内皮细胞等细胞膜上,属于细胞膜受体。
是白细胞表面发现的含有7个跨膜结构域的G蛋白偶联受体。
(依靠一个跨膜蛋白数据库,包含了每个序列的一些附加信息:如跨膜结构区域的数量、跨膜结构域的位置及其侧翼序列的情况,来预测是否是受体。
趋化因子的特征及功能其特征和功能为:a使细胞骨架重排,引起细胞形态改变;b 肌动蛋白的聚合与断裂,引起板层足(lamellipodia)的形成和退缩(retraction);c引起整合蛋白的上调和活化,使白细胞粘附血管壁的内皮细胞;d使活化的白细胞内游离钙离子浓度升高;e产生杀微生物的活性氧及有生物活性的脂类物质;f释放贮存于细胞内颗粒的内容物,如中性白细胞和单核细胞内的蛋白水解酶、嗜碱性白细胞内的组胺和白三烯、嗜酸性白细胞内的细胞毒性物质.而且在非正常生理状态.趋化因子受体的分类同CC 类趋化因子结合的受体称为CC类受体(CCR), 同CXC类趋化因子结合的受体称为CXC 类受体(CXCR),同样有C和CX3C受体(CR、CX3CR)。
趋化因子与受体的关系:在配体-受体结合实验中, 可以看到趋化因子与受体间结合的冗余现象, 即一个趋化因子可与数个趋化因子受体结合,一个趋化因子受体可与数个趋化因子结合。
因此, 在体外趋化实验中表现为。
一种趋化因子可以趋化表达不同趋化因子受体的免疫细胞做定向迁移,一种免疫细胞可以为多种趋化因子所趋化。

趋化因子家族及其受体基础研究进展趋化因子(Chemokine)是一类小分子碱性蛋白,主要的功能是能够趋化细胞定向移动。
目前已经发现的趋化因子有50多种,随着研究的深入,趋化因子及其受体的结构、功能及在体内的作用已经被众多的研究者发现。
趋化因子及其受体的相互作用,可以参与多种生理功能,比如细胞的生长、发育、分化、凋亡和分布等,在病理过程中也具有重要作用,如炎症反应、病原体感染、创伤修复及肿瘤形成和转移等。
趋化因子一般由70-125个氨基酸组成,分子量较小(6-14KD)。
按照一级肽链结构特点,其N端半胱氨酸残基的位置和数目可将趋化因子分为4个亚族:CC、CXC、C和CX3C(C为半胱氨酸,X为任意氨基酸)。
四类趋化因子结构相似性较高,氨基酸序列具有一定的同源性。
根据趋化因子的表达方式以及其在免疫系统中的作用,可以将他们分为两类:内环境稳定性趋化因子和炎症性趋化因子。
内环境稳定性趋化因子主要在归巢场所表达,有着维持内环境稳态的功能,并且对淋巴细胞归巢及成熟有着明确的作用。
炎症性趋化因子由受到刺激的细胞表达,如炎性细胞因子的诱导、细菌毒素或其它破坏内环境稳定的因素的刺激,主要功能是募集效应细胞,在协调天然和获得性免疫反应中起重要作用。
大多数的趋化因子属于CC和CXC两个亚族族。
其中CC亚族有28个成员(CCL1-CCL28),主要对中性粒细胞、单核细胞、肥大细胞、树突细胞、NK细胞、T和B淋巴细胞等具有强大趋化活性,比较重要的有:单核细胞趋化蛋白(MCP-1/CCL2)、巨噬细胞炎症蛋白(MIP/CCL3)、正常T细胞表达和分泌,活化时表达下降的因子(RANTES/CCL5)等;CXC亚族有17个成员(CXCL1-CXCL17),CXC亚家族主要作用于中性粒细胞,这个亚族比较重要的趋化因子有:白细胞介素-8(IL-8/CXCL8)、γ干扰素诱生的单核因子(Mig/CXCL9)、γ干扰素诱生蛋白10(IP-10/CXCL10)、基质细胞来源因子1(SDF-1/CXCL12)等。



趋化因子及其受体趋化因子是一类对细胞具有趋化作用的蛋白质分子,它们在生物体内控制着细胞的运动和定向迁移。
趋化因子通过结合与其相互作用的受体,激活下游信号传导途径,从而引导细胞朝特定方向移动。
趋化因子和受体的研究对于理解生物体内细胞运动的机制、生物发育、免疫系统的功能等具有重要意义。
趋化因子主要可以分为几类,包括细胞外基质趋化因子、细胞因子趋化因子以及补体活化产物等。
细胞外基质趋化因子主要包括纤维连接蛋白、胶原蛋白、血小板聚集素等,它们通过与细胞膜上的受体相互作用,引导细胞移动向特定的方向。
细胞因子趋化因子则包括趋化细胞催化蛋白、生长因子、细胞因子等,它们通过作用于特定细胞表面受体,调控细胞的迁移和定向移动。
补体活化产物是机体内免疫炎症反应过程中的趋化因子,在感染和组织损伤的情况下,补体系统会被激活产生许多趋化因子,引导免疫细胞聚集并参与免疫炎症反应。
酪氨酸激酶受体是另一类重要的趋化因子受体,它们是由单个或多个酪氨酸激酶蛋白亚基组成的受体。
酪氨酸激酶受体对于趋化因子的信号转导主要通过磷酸化和激酶级联反应来实现。
丝氨酸/苏氨酸激酶受体是受体酪氨酸激酶家族中的一个亚家族,它们的功能类似于酪氨酸激酶受体,但信号转导途径有所不同。
除了上述两个受体家族之外,还有一些其他类型的趋化因子受体。
例如,趋化因子蛋白激酶受体(PAR)可以通过激酶级联反应调控细胞的运动和定向迁移。
核受体趋化因子,如核内受体CXCR4,可以激活转录因子并介导细胞迁移。
趋化因子和受体的研究对于理解细胞的运动机制、发育和免疫系统的功能具有重要意义。
通过研究特定趋化因子和受体,可以揭示细胞运动和定向迁移的分子机制,并且为治疗炎症和癌症等疾病提供新的靶点。
因此,深入了解趋化因子和受体的结构和功能对于生物医学研究具有重要意义。

趋化因子SDF—1研究进展作者:张罡瑜杭秋琦来源:《中国民族民间医药·下半月》2016年第10期【摘要】趋化因子SDF-1属于CXC类趋化因子,其与机体的胚胎发育、细胞的迁徙与归巢、血管形成与修复、细胞分化、免疫、肿瘤发生等方面均有密不可分的联系。
文章对SDF-1来源、结构、生物特性、与疾病的联系、临床应用等方面的中、西医研究进展进行综述。
【关键词】 SDF-1; CXCL-12;研究进展【中图分类号】R2-031 【文献标志码】 A 【文章编号】1007-8517(2016)20-0051-06SDF-1(Stromal Cell-derived Factor-1)是一类具有趋化活性的细胞因子,系统命名为CXCL-12,是CXC类趋化因子亚家族的一名成员。
CXCL-12是由基质细胞持续分泌的,因此CXCL-12被命名为基质细胞衍生因子-1(Stromal Cell-Derived Factor-1, SDF-1)。
SDF-1有SDF-1α和SDF-1β两种亚型,SDF-1α为其主要亚型。
SDF-1在淋巴结、肺、肝、骨髓、小肠、肾、皮肤、脑和骨骼肌中均有表达,这些组织器官的血管内皮细胞、间质成纤维细胞和成骨细胞均可持续性分泌SDF-1[1]。
且在一些病理性疾病如炎症、缺血和缺氧,以及在促血管生成的环境下如肿瘤和自身免疫疾病中,SDF-1的表达量会显著升高[2]。
又因其对祖B和前B 细胞有趋化作用,故又称其为前B细胞刺激因子(Pre-B-Cell Growth-Stimulating Factor,PBSF)[3]。
1 SDF-1的结构特征趋化因子是一类结构相似的小分子(8-10 kD)蛋白质。
因靠近分子N-端的两个半胱氨酸(Cys)的位置状态不同,趋化因子被分为了4个亚类:CXC(插入一氨基酸残基)类、CC (不插入氨基酸残基)类、C(N端只有一个Cys)类和C3XC(插入3个氨基酸残基)类。
趋化因子SDF-1及受体CXCR4研究进展第23卷第1期2OO6年2月生物学杂志JOURNALOFBIOLOGYV o1.23No.1Feb,2006趋化因子SDF一1及受体CXCR4研究进展储子彦,陈晓萍,方晶晶(浙江工业大学生物与环境工程学院,杭州I310014)摘要:趋化因子(chenmklne)是一类一级结构相似,以对白细胞等多种细胞具有趋化定向运动作用为特征的小分子蛋白.功能研究表明,趋化因子在胚胎发育,血管生成,炎症,肿瘤,史滋病等机体多种生理和病理过程中发挥重要作用,部分趋化因子的衍生物或抑制物具有潜在的临床应用前景.不久的将来,趋化因子及其受体可能成为疾病治疗的分子靶点.关键词:趋化因子;SDF一1;CXCR4中图分类号:4文献标识码:A文章编号:1008—9632(2006)Oi一0011—03趋化因子是一类重要的免疫调节因子,直接引导自细胞,包括多种免疫活性细胞进行有方向性迁移,不仅能精确地调节免疫系统的反应,还对组织,器官形成,造血系统功能有调节作用.1趋化因子与受体的结构趋化因子分子结构中有4个保守的半胱氨酸,形成2对二硫键,可分成4个亚类:(1)CC亚类,2对二硫键间无其它氨基酸间隔;(2)CXC亚类,间隔1个氨基酸;(3)Cx3C亚类,间隔3个氨基酸;(4)C亚类,仅有1对二硫键.SDF一1(stromalcell—derivedfactor1)基质细胞来源因子,属于趋化因子CXC亚家族,编码区含267bp,编码89个氨基酸残基多肽.CXCR4为SDF一1受体,高度保守,a螺旋跨膜7次,由352个氨基酸组成,在人体内,编码基因位于人染色体2q21,有一个胞外N端,3个胞内环,3个胞外环和1个胞内C端,SDF—l与CXCR4的N端结合,并与CX—CR4和第二胞外环ECI_2(secondextracelluarloop)相互作用才能启动下游信号通路.2SDF一1/CXCR4生物学意义2.1与HW病毒感染的关系CXCR4为嗜T细胞性SI株辅助受体,能与CD4协同作用,参与CD4抗原与HW表面糖蛋白gpl20结合介导病毒吸附侵入的过程,CXCR4的N端结构参与病毒结合,有多个CXCR4结构域特别是第二细胞外环结构能与HⅣ相互作用.趋化因子与受体的结合能阻断受体与HⅣ的结合位点防治HⅣ进入细胞,同时趋化因子对受体的封闭和下调作用也成为一个有效的防治手段,故趋化因子及其衍生物是辅助受体拮抗剂主要成分之一.Fig1Representationof313modelforSDF—ld(NMR}围1SDF一1n的3D模型除了SDF一1的衍生物,还发现CXCR4的其他抑制剂,如:AMD一3100通过与CXCR4的第二膜外环的负电荷区域结合,成为迄今为止与CXCR4结合最有效的非肽类抑制剂.2.2与造血细胞的关系2.2.1SDF一1对造血于/祖细胞增殖和分化的影响Lataillade等【J采用无血清培养基在CD34细胞体外培养中发现,SDF—la对造血干细胞有刺激其增殖的作用,并且与其他造血细胞生长因子(HCF)如干细胞因子(SCF)及白介素一3(IL一3)有协调作用,它还能促进更多的CD34细胞进入细胞周期,使s期和C2期/M期细胞明显多于对照组.2.2.2SDF一1介导造血干/祖细胞的动员过程研究表明,G—SCF动员外周血时,多种蛋白水解酶作用于胞外基质,粘附因子,细胞因子及趋化因子解除细胞与细胞之间形成的紧密结合,促进造血干细胞收稿日期:2005—05—08;修回日期:2005—08—04作者简介:储子彦(1982一),女,汉族,本科生;通讯作者:陈晓萍(1961一),女.汉族,副教授,博士,研究方向为趋化因子作用的分子机制,E—mail:ch)66@.基金项目:国家人事部留学择优基金和浙江省教育厅基金资助项目,项目号~20040586第23卷第1期2006年2月生物学杂志JOURNALOFBIOLOGYV o1.23No.1Feb,2006跨越内皮细胞的转运[21,裂解SDF一1及HSC表面的CXCR4的N端,使HSC失去对SDF一1的趋化作用.骨髓动员的鼠中,骨髓SDF一1浓度会下降,直接导致体内丝氨酸蛋白酶的积累,后者直接引起骨髓造血干/祖细胞的动员[3l.Fig2Cartoonrepresentationof3DmodelforCXCR4receptor图2CXCR4的3D模型图2.2.3对造血干/祖细胞的归巢的影响2.2.3.1SDF一1/CXCR4介导造血干/祖细胞的趋化转移效应来源于骨髓,脐血,动员外周血中的CD34造血干细胞表面表达SDF一1受体CXCR4,而骨髓基质表达的SDF一1能特异对CXCR4产生趋化作用【.因此表达CXCR4的造血干细胞就能够沿着SDF一1的浓度梯度迁移实现归巢过程.CXCR4抗体孵育后的干细胞无法实现归巢,诱导CXCR4在CD34上的表达,能提高造血干l细胞移植成功率.Benb0ubke一5J在研究造血干细胞活动能力与SDF一1基因多态性之间相关性时发现SDF一13'A等基因存在时唯一能预测CD34细胞具有良好动员能力的因素.2.2.3.2SDF一1/CXCR4诱导CD34细胞穿越内层黏附于骨髓基质的作用对CD34细胞进行趋化活性试验发现,SDF一1能引起造血干/祖细胞迁移率增20~1[6].人的骨髓内皮细胞连续表达SDF一1,SDF—l吸附在内皮细胞表面的蛋白多糖上,结合CD34细胞表面的CXCR4捕获造血干/祖细胞,在黏附分子参与下. HSC与内皮细胞产生黏附,SDF—la可增强HSPC与骨12髓基质的黏附作用,这与SDF—la的聚糖和葡萄糖胺聚糖的特性有关.2.3SDF一1/CXCR4在免疫功能上的作用SDF一1/CXCR4是B细胞生长发育成熟的关键细胞因子,骨髓中前B细胞形成需要CXCR4,而胚胎期SDF一1缺乏会损害胎肝前B细胞的发育【.研究发现,SDF一1缺乏鼠的淋巴细胞和骨髓系细胞不能正常发育,由于SDF一1和CxcR4对T细胞,单核细胞的迁移及B淋巴细胞生成中的生物学效应,可能在免疫监视过程中也发挥重要生理功能,在炎症反应中,白细胞向炎症部位集中也与此有关.利用双室模型中研究发现SDF一1的趋化活性较其他趋化因子高1O倍,局部有放射状细胞积聚现象,并观察其趋化活性有浓度梯度依赖性,SDF一1是CD4T细胞活化的共刺激因子,在类风湿性关节炎病变的滑膜中有CD4T细胞的积聚现象,提示SDF一1在免疫及局部炎症过程中是一个重要的调节因子.2.4SDF一1/CXCR4维持胚胎发育敲除小鼠SDF一1基因的2个等位基因,小鼠出生后即死亡,有B细胞增殖,骨髓细胞发育,神经系统发育受阻以及室间隔缺损等缺陷,基因敲除CXCR4小鼠与基因敲除SDF一1小鼠有几乎相同的表现,说明SDF 一1/CXCR4在胚胎发育过程中具有非常重要的作用.2.5sDF一1/CXCR4与肿瘤转移的关系很多研究报告指出,CXCR4在肿瘤细胞高度表达,而SDF一1在某器官的高浓度表达代表肿瘤细胞首先转移的目的地,实验证明CXCR4的抗体有效地阻断了肿瘤细胞向肺部转移的过程,可见SDF一1/CXCR4在肿瘤细胞扩散转移中起到重要作用.CXCR4是肿瘤细胞上很常见的趋化因子受体,同时是其他治疗癌症中的调节因子,例如血管内皮生长因子(VEGF)能诱导CXCR4大量表达,因此ⅦCF抗体能通过抑制CXCR4表达,达到控制肿瘤细胞转移的作用.已发现AMD3100为CXCR4的特异性拮抗剂,通过抑制CXCR4/SDF一1的结合,抑制肿瘤细胞的定向转移,另外,在乳腺癌患者中雌激素受体(ER)表达明显增高,雌激素的真正标靶为SDF—l,能迅速诱导SDF—hx,SDF一1l3的产生,这种效应可被纯ER拮抗剂ICⅡ82780拮抗.3SDF—l/CxCR4的信号转导SDF—l与CXCR4结合,激活了CxCR4受体耦联的c蛋白,通过激活磷脂酰激醇一3激酶(PI3),丝裂原结合蛋白酶(mitogen—associatedproteinkinase,MAPK)和第23卷第1期2006年2月生物学杂志JOURNALOFBIOLOGYV0J.23No,lFeb,2006转录因子NF—KB,为SDF一1/CxCR4一PI3一~LAJPK—NF一皿信号通路,具体如下:(1)SDF一1刺激可使PI一3激酶P85亚单位及PLC一了酪氨酸磷酸化,PLC一可激活PKC,PKC进一步激化一系列局部黏附蛋白,从而介导细胞迁移.SDF一1诱导的细胞趋化可被PI一3激酶抑制剂wortmannin和PKC抑制剂GF109203X所阻断.(2)SDF一1刺激引起核转录因子NF—B活化,支持SDF一1/CXCR4在细胞,增殖及维持生存中发挥作用. (3)SDF一1刺激还经MEK(MAPKandERKkinase)激活P44/42ERK(ExtraceUnlarSignal—regulatedki—nase),但不能激活p38或JNKMA_PK(Mito—activated proteinkinase).SDF一1诱导的细胞也不能被MEK抑制剂PD98059阻断,考虑MEK可能在SDF一1/CXCR4诱导的细胞内信号转导中起调节作用.4结束语SDF一1和CXCR4的特异性结合及其所启动的下游信号通路在细胞的胚胎发育,造血,免疫及HIV治疗上具有重要的意义,对具体的特异性识别过程及调控环节的详尽研究,将有助于我们深入理解细胞问相互作用以及人体中正常的生理功能,并有助于人们开发各种抑制齐IJ运用于临床治疗.参考文献:[1]LatailladeJJ,ClayD,DupuyC,cta1.ChemoHneSDF一1enhanc~ clreulatlngCD34cellprolifemdoninsynergywithcy[0nespossi—bleroleinrogenitorsurvival[JJ.Blood,2000,95:756—768.[2]Va&yGG,Lidey0.Extracelhlarmatrixmot/ct/s,Cytokines,anden—zymes,dymmiceffectsonimmunecellbe|mviorandinflammation [J].JI_eukecBiol,2000,67:149—159.[3]LevesqueJP,HendyT,TakamatsuY,eta1.DisruptionoftheCX—CR4/CXCLRchemotacticinteractionduringhematopoictiestemcell mobilizationinducedbyG—SCForcyclophosphamidelJJ.JElla Invest,2003,111:187—196.[4]WrightDE,BowtnanEP,WagersAJ,eta1.Hematopoictic8[eIilceils areuniquelyselectiveintheirmigratoryresponsetochemokines[JJ. JExpMed,2002,195:1145~1154[5]BenboubkerL,WarierH,CationA,eta1.BrJHaematol,2001,ll3 (1):247.[6]NaiyerAL.JodY,AhnJ,eta1.Stromalderivedfactor一1induced clleImnesofcordbloodCD34cells(1ongtermcuhttreinitiatingcells)th~ue,hendothelialcellsismediatesbyE一~lecfin[J】.Blood,1999,94:柏11一柏19.[7]Fe~T,KawabataK,KawamotoH,eta1.TheearlieststagesofB celldevelopmentrequireachemokinestromalcell—derivedfactor/pm—B础growth—stimulatingfactor[J].Immunity,2001,15(2):323334.ProgressesofchemokineSDF一1andCXCR4CHUZi—yah,CHENXiao—ping,FANGJing—jing (ZhejiangUniversityofTechnology,BiologicalandEnvimnmentalEngineeringCollege,H angzhou310014,China)Abstract:Chemokineisagroupofsmallmolecularproteinswithsimilarilyprimarystructure andchemotaxisactivity.Re—cendyresearchshowedtheirimportantregulatingeffectsonembryonicdevelopment,vascul ogenesis,inflammation,tumor,AIDSandmanypathologicprocess.Thederivantofchemokineoritsinhibitorsshowdgoodclinical applicationprospect.Nearlyfuture, chemokinesandtheirreceptorswillbethetherapeuticmoleculartargettodiseases. Keywords:chemokine;stromalcell—derivedfactor一1;CXCreceptor4(上接7页)[17]张伟.滚筒式填料筛板萃取器反胶团法萃取蛋白质[J]化工,2000.12(增刊):227~231.[18]赵雪雁,佟晓冬,孙颜.StreamlineSP膨胀床纯化溶菌酶[J].过程工程2004,4(2):104—110[19]MamanoJGSandTsotsisTT.CatalyticMembranesandMembraaeReactors[M].Wiley—VCHVerlagGmbH,WeinheimPress,2002. ProgressesofresearchonthepurificationandseparationofthelysozymeFENGOuan.NGBin(AnhuiUniversityofTechnologyandScience,Wuhu241000,China)Abstract:Recentresearchprogressesonthepurificationandseparationofthelysozymewerereviewed.suchasion—ex—changechromatography,ultrafiltmtion,reversedmicellarextraction,affinity—basedbioseparationandsoon.Atthesadietime,de. velopmentprospectsofwaysofthepurificationandseparationofthelysozymewerediscusse d-Keywords:lysozyme;purification;separation13。
趋化因子CCL4和CCL5——新的肝癌标志物作者:宁秀景来源:临床肝胆病杂志原题Serum levels of chemokines CCL4 and CCL5 in cirrhotic patients indicate the presence of hepatocellularcarcinoma.大多数肝细胞癌被诊断时已经是晚期。
血清肿瘤标志物甲胎蛋白(AFP) 和脱-γ-羟基凝血酶原 (DCP) 的诊断价值是有限的。
我们研究的目的是评价血清生长因子、凋亡因子和炎症介质的水平对肝硬化患者是否伴有肝癌的诊断价值。
血清标本从患有肝癌(n = 61)和没有肝癌(n = 78)的肝硬化潜在肝脏移植患者(LTx) 以及健康对照组(HCs; n = 39)中收集。
测定血清CRP、新喋呤、炎症介质白细胞介素-6 、促血小板生成素 (TPO)、粒细胞集落刺激因子(GCSF)、纤维母细胞生长因子(FGF )、血管内皮生长因子(VEGF)、高迁移率族蛋白B1(HMGB1)、人细胞角蛋白18- M65 和人细胞角蛋白 18 凋亡片段 M30 以及一组促炎趋化因子 (CCL2 、CCL3 、CCL4、CCL5、 CXCL5、 IL-8)的浓度。
应用卡方检验,Fisher 确切检验,曼-惠特尼 U-检验(非参数检验)、 ROC 曲线分析和逐步逻辑回归分析方法。
结果: 肝细胞癌患者有较高的血清促血小板生成素和趋化因子(P < 0.001,TPO、CCL4、CCL5 和CXCL5) 水平,而趋化因子CCL2 (P = 0.008) 水平比不伴有肝癌的肝硬化患者低。
重要参数的多元逐步回归分析表明,研究参数中的 CCL4 和 CCL5 是 (P = 0.001) 是肝细胞癌的诊断标志物。
肝硬化患者血清中促血小板生成素和趋化因子水平较低,而人细胞角蛋白 18 凋亡片段 M30 水平却显著高于肝癌患者。
肝硬化患者血清高水平炎症趋化因子如CCL4 和 CCL5 提示肝细胞癌的存在。