2011年型207介绍
- 格式:pdf
- 大小:372.64 KB
- 文档页数:6
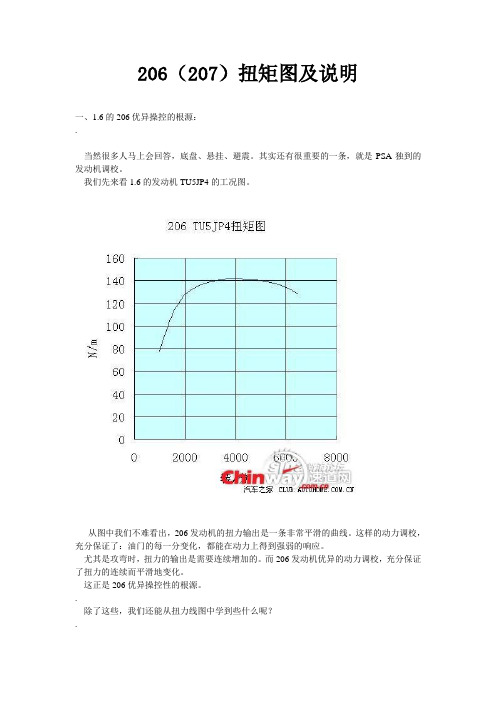
206(207)扭矩图及说明一、1.6的206优异操控的根源:.当然很多人马上会回答,底盘、悬挂、避震。
其实还有很重要的一条,就是PSA独到的发动机调校。
我们先来看1.6的发动机TU5JP4的工况图。
从图中我们不难看出,206发动机的扭力输出是一条非常平滑的曲线。
这样的动力调校,充分保证了:油门的每一分变化,都能在动力上得到强弱的响应。
尤其是攻弯时,扭力的输出是需要连续增加的。
而206发动机优异的动力调校,充分保证了扭力的连续而平滑地变化。
这正是206优异操控性的根源。
.除了这些,我们还能从扭力线图中学到些什么呢?.【高速攻弯的极限】注意:这个弯不是指高速公路上的弧形路。
而是指弧度更锐利的弯道。
在高速攻弯中,最危险的事情,就是:弯中失力。
弯中失力,车的操控性会急剧下降,乃至失控。
所以高速攻弯平顺,最重要的一条,就是动力输出要连续而平滑的增加。
.206光滑而连续的扭力输出曲线,正是弯道性能的有力保证。
但是,车的性能总是有极限的。
那么,1.6的206极限是多少呢?从扭力图中我们不难看出,当发动机转速超过3400转后,动力增长就非常平缓了(也可以说几乎不增加了)。
所以为了给攻弯时留有足够的动力升值余地,1.6的206高速入弯的发动机转速最好不要超过3000转。
3000转换算成五档的速度相当于时速100公里。
上次有个兄弟发贴说:他以时速120公里的入弯速度下四环匝道,小六发生了明显的侧滑,几乎要失控了。
问题的关键就在于:他那时高速入弯的转速已经超过了3400转,这时你弯中再想通过踩油门去增加动力,就超越了小六的极限。
.【收油的问题】从扭力曲线中我们可以看出,当我们踩下油门,发动机转速从一千多转升到两千多转时,发动机动力的增加非常迅猛。
但是,这个问题反过来看:当我们松掉油门(就是所谓的带档滑行)后,随着发动机转速的下降,发动机的动力输出下降得也非常迅速。
所以,206带档溜车是溜不远的。
也就是说:从操控上讲,不206想更省油,溜车溜更长的距离,只能用空档溜车。

小麦品种百农207高产栽培技术赵 彬(河南省沈丘县种子技术服务站,沈丘 466300)摘要:百农207是河南科技学院欧行奇教授主持培育的高产、稳产、大穗、抗病性强的小麦品种,2013年通过国家农作物审定委员会审定,审定编号为国审麦2013010。
结合高产户种植经验,总结了该品种的高产栽培技术。
关键词:小麦;百农207;高产栽培技术百农207是河南科技学院欧行奇教授主持培育以周16为母本、百农64为父本杂交选育而成的高产、稳产、大穗、抗病性强的小麦品种。
2013年通过国家农作物审定委员会审定,审定编号为国审麦2013010,适宜黄淮冬麦区南片的河南中北部、安徽北部、江苏北部、陕西关中地区高中水肥地块早中茬种植。
该品种在沈丘县大面积种植多年,表现抗病性强、产量高,深受种植户欢迎,结合试验、示范和群众高产典型户种植经验,总结出一套产量650kg以上的高产栽培技术要点。
1 特征特性该品种属中早熟。
幼苗半匍匐,长势旺,叶宽大,叶深绿色。
冬季抗寒性中等。
分蘖力较强,分蘖成穗率中等。
早春发育较快,起身拔节早,两极分化快,抽穗迟,耐倒春寒能力较强。
中后期耐高温能力较好,熟相好。
株高76cm左右,株型松紧适中,茎秆粗壮,抗倒性较好。
穗层较整齐,旗叶宽长、上冲。
穗纺锤形,短芒,白壳,白粒,子粒半角质,饱满度好。
每667m2穗数40.2万~47.3万穗,穗粒数35.6~47.4粒,千粒重41.7~47g[1-2],是一个很有推广前景的小麦新品种。
2 产量表现2009-2010年参加国家黄淮片小麦区试预备试验,比对照周麦18增产6.0%。
2010-2011年参加黄淮冬麦区南片冬水组品种区域试验,每667m2平均产584.1kg,比对照周麦18增产3.9%;2011-2012年续试,平均产510.3kg,比周麦18增产5.3%。
2012-2013年生产试验,平均产502.8kg,比周麦18增产7.0%[1-2]。
2012-2013年在沈丘种植20hm2,2013年春在其他小麦品种遭受不同程度倒春寒危害大幅减产的情况下,百农207无冻害,表现抗寒性较好,取得大丰收,特别是邢庄乡王新亮的责任田种植的百农207每667m2产量达670kg。

㊀第52卷第2期郑州大学学报(理学版)Vol.52No.2㊀2020年6月J.Zhengzhou Univ.(Nat.Sci.Ed.)Jun.2020收稿日期:2019-03-05基金项目:国家自然科学基金项目(31571658);南京农业大学作物遗传与种质改良国家重点实验室课题(ZW2011002);河南省科技攻关重大项目(151100110700)㊂作者简介:马超(1994 ),男,河南项城人,硕士研究生,主要从事小麦分子生物学研究,E-mail:machao0813@;通信作者:刘文轩(1964 ),男,河南巩义人,教授,主要从事小麦细胞与分子遗传研究,E-mail:liuwenxuan@㊂高产小麦新品种百农207及其姊妹系的基因组构成分析马㊀超1,㊀吴㊀皓2,㊀欧行奇3,㊀李新华4,㊀乔㊀红4,㊀秦广雍4,㊀刘文轩1,㊀亓增军2(1.河南农业大学生命科学学院㊀河南郑州450002;2.南京农业大学作物遗传与种质改良国家重点实验室㊀江苏南京210095;3.河南科技学院生命科技学院㊀河南新乡453003;4.郑州大学农学院㊀河南郑州450001)摘要:结合染色体荧光原位杂交和小麦15K 育种芯片杂交技术,对黄淮南片麦区的主导新品种百农207及其双亲百农64和周麦16,以及10个姊妹系的染色体和基因组构成进行了分析㊂结果显示:百农64对百农207基因组的贡献率为55.3%,周麦16的贡献率为40.7%,并且百农207具有64个新位点或重组位点㊂此外,在参试材料中检测到9种染色体变异类型,其中5种来自百农64,4种来自周麦16或由双亲染色体重组而产生㊂结合荧光原位杂交和双亲差异SNP 位点重组分析,将6B 染色体臂间倒位区段初步定位在短臂50.1Mb 到长臂475.5Mb 之间,5A 染色体长臂上的重复序列区段定位在66.1Mb㊂关键词:小麦新品种;百农207;基因组构成;荧光原位杂交;SNP 位点;染色体结构变异中图分类号:S512.1㊀㊀㊀㊀㊀文献标志码:A㊀㊀㊀㊀㊀文章编号:1671-6841(2020)02-0118-09DOI :10.13705/j.issn.1671-6841.20195580㊀引言面包小麦(Triticum aestivum L.,2n =2x =14,AABBDD)是小麦属异源六倍体物种,是目前世界上种植最广㊁交易量最大的粮食作物之一,也是全球植物蛋白的主要来源[1]㊂因此,小麦的可持续生产对世界范围内的社会进步和稳定具有重要意义㊂自20世纪60年代绿色革命以来,世界小麦产量增加了2倍,预计在21世纪中叶将继续保持增长[2]㊂在此期间,小麦产量的增长主要是通过提高单位面积的作物产量来实现的㊂培育遗传背景优良㊁种子耐贮藏的新品种,提高肥料利用率,施用农药和杀虫剂,改善灌溉等都是小麦单产增加的因素㊂其中,选育和推广高产㊁适应性广㊁品质好㊁抗逆性强㊁肥料利用率高的小麦新品种对小麦产量的提高发挥了重要作用[3]㊂小麦新品种选育的成功始于选择合适的亲本,优良亲本的杂交组合往往会选育出一批好的品种㊂例如,周8425B 是20世纪70年代从国际玉米小麦改良中心引进的合成六倍体小黑麦(2n =6x =42,AABBRR)产生的小麦-黑麦自发罗伯逊易位(RT)1BL.1RS 系㊂以其为亲本,在河南省及周边地区培育出了100多个品种,其中周麦16号㊁矮抗58号等79个品种通过国家或省级审定[4]㊂小麦品种小偃6号是小麦与十倍体长穗偃麦草(Thinopyrum ponticum ,2n =10x =70,J S J S J S J S JJJJJJ)杂交的后代,曾经是中国种植面积最大的小麦品种,以小偃6号为核心亲本,选育出了50多个大面积推广品种,累计推广面积达到2000万hm 2,增产1500万t [5]㊂因此,解析大面积推广品种及其核心亲本的遗传组成,有助于了解如何培育优良品种,提高重要农艺性状和育种效率㊂近年来,不少研究人员利用简单序列重复(SSR)标记㊁高密度单核苷酸多态性(SNP)标记或高分辨率荧光原位杂交(FISH)对小麦主栽品种和亲本的遗传基础进行了解析[6-7],但尚未见911㊀第2期马㊀超,等:高产小麦新品种百农207及其姊妹系的基因组构成分析到结合FISH和基因芯片杂交技术对小麦品种进行遗传解析的报道㊂百农207是2014年通过国家农作物品种审定委员会审定的小麦新品种,自2016年起成为河南省和我国小麦主产区黄淮南片年种植面积最大的高产品种(135万hm2)㊂其亲本周麦16和百农64也是在该地区曾经大面积推广的品种,是目前育种家最常使用的核心亲本[8-9]㊂本研究结合高分辨率FISH和小麦15K SNP芯片杂交,对百农207及其亲本和姊妹系进行了染色体和基因组构成2个层次的分析,旨在为我国黄淮河流域南部地区优良新品种的培育,以及为百农207及其亲本百农64和周麦16的更好利用提供依据㊂1㊀材料与方法1.1㊀植物材料本研究共选用13个小麦品种(系),其主要农艺性状如表1所示㊂其中百农207是以周麦16为母本,与百农64杂交选育的,同时从该杂交组合中还选育出10个不同农艺性状的姊妹系㊂所有材料均由河南科技学院提供㊂表1㊀所用材料及其主要农艺性状Table1㊀Materials used and their key agronomic characteristics品种(系)名称主要特征㊁特性备注百农64(BN64)综合抗性强,小粒饱满,优质;但分蘖成穗率低,穗轴易断,粒重一般父本㊀周麦16(ZM16)高产,矮秆,大粒;但饱满度差,不耐倒春寒,不抗穗发芽,穗粒数偏少母本㊀百农207(BN207)高产稳产,抗倒春寒和穗发芽,千粒重高;但偏晚熟,抗锈病能力一般华育166(HY166)与百农207株高相当,穗大,抗倒性好;但穗数偏少姊妹系冠麦1号(GM1)综合性状好;但籽粒黑胚率偏高姊妹系百旱207(BH207)较百农207高,抗倒性好,穗较百农207多;但穗粒数较百农207少姊妹系百农69-38(BN69-38)矮秆,抗倒;但穗数偏少,籽粒黑胚率偏高姊妹系PJ11-15综合性状好;但抗倒性较差姊妹系PJ11-52综合性状好;但抗倒性较差姊妹系百农10-8(BN10-8)综合性状好;但落黄较差姊妹系百农12-40(BN12-40)矮秆,抗倒;但年际间成穗数不稳定姊妹系百农14-818(BN14-818)矮秆,周麦16类型;但籽粒饱满度一般姊妹系华育198(HY198)穗多,籽粒商品性好,落黄好;但抗倒春寒能力一般姊妹系1.2㊀通过小麦15K基因芯片杂交筛选SNPs标记每个材料从20个幼苗中采集新鲜叶片,用CTAB(十六烷基三甲基溴化铵)法提取基因组DNA,并用1%琼脂糖凝胶电泳检测DNA提取液的完整性和浓度㊂然后将合格DNA样品与含有13947个SNP标记的15K基因芯片杂交,筛选SNPs标记㊂该芯片由中国农业科学院设计,杂交由中金玉生物技术有限公司实施完成㊂1.3㊀SNP数据分析与基因型图谱构建利用得到的SNP基因型数据对百农207及其亲本和姊妹系的基因组构成进行分析㊂如果特定SNP位点的百农207基因型与特定亲本的基因型相同,则认为百农207从该特定亲本遗传了该基因座㊂存在于百农207中但双亲均不存在的基因座被视为新基因座,而在百农207中不存在的双亲基因座被视为缺失基因座㊂随后,计算每个亲本对百农207基因组贡献的SNP位点数㊂一个亲本对百农207的基因组贡献被定义为从一个亲本遗传的SNP位点数量与亲本间多态型SNP位点总数之比㊂根据SNP位点侧翼序列与中国春参考序列v.1.1(IWGSC2018)的序列比对,将不同染色体上的SNP位点从短臂到长臂进行排序㊂用TASSEL v5.0主成分分析法(PCA)对百农207的亲本和姊妹系进行聚类分析[10]㊂1.4㊀根尖细胞染色体制备及荧光原位杂交按文献[7]所述进行根尖细胞染色体制备,使用BX51奥林巴斯相差显微镜(日本东京奥林巴斯公司)进行细胞学观察㊂以8种单链寡核苷酸为探针进行双色非变性原位杂交(ND-FISH)㊂用6-羧四甲基罗丹明(TAMRA)标郑州大学学报(理学版)第52卷记的pAs1-1㊁pAs1-3㊁pAs1-4㊁pAs1-6㊁AFA-3㊁AFA-4探针产生红色信号,而6-羧富勒烯(FAM)标记的pSc119.2-1和(GAA)10重复序列产生绿色信号㊂所有寡核苷酸均由上海生工生物科技有限公司合成㊂杂交后,在BX51奥林巴斯荧光显微镜下观察染色体,并通过SPOT CCD(点冷彩色数码相机)拍摄图像,使用Adobe Photoshop(v6.0)(美国Adobe)进行分析㊂1.5㊀抗穗发芽基因分子标记分析基于文献[11-14]设计抗穗发芽基因的PCR引物序列,由上海生工生物科技有限公司合成(表2)㊂进行PCR扩增反应混合液体积为15μL,分别含有7.5μL2ˑEasyTaq-PCR Supermix(北京全式金科技公司)㊁0.5pmol正向和反向引物以及100ng基因组DNA㊂PCR扩增使用F50SSR反应系统[15]㊂PCR产物经1.5%琼脂糖凝胶分离,溴化乙啶染色后用Tanon2500凝胶成像系统(上海Tanon科技有限公司)进行观察㊂表2㊀抗穗发芽基因分子标记引物Table2㊀Primers for molecular markers of pre-harvest sprouting resistance genes标记名称上游/下游引物序列退火温度/ħ特异片段/bp位置参考文献TaVp1B3TGCTCCTTTCCCAATTGG60569/8453BL[11]ACCCTCCTGCAGCTCATTGPM19-A1GAAACAGCTACCGTGTAAAGC611174AL[12]TGGTGAAGTGGAGTGTAGTGGDorm-1GTTCCTCCCACCAAATCTCA666067BL[13]GCCCGGTCTAAACGTACGATamyb10-LP9/RP3TAGGCCAACACCTTCTAAACG6013533DL[14]2㊀试验结果2.1㊀百农207的产量及主要农艺性状百农207是以周麦16为母本㊁百农64为父本杂交选育的半矮秆小麦品种㊂周麦16和百农64都是国家审定的高产小麦品种,在河南和邻近省份常作为核心亲本使用㊂其中周麦16是高产半矮秆品种,大穗大粒,但抗倒春寒和抗穗发芽性差㊁千粒重低;而百农64对条锈病和叶锈病的综合抗性强,品质好,籽粒饱满,但分蘖力低,成熟时小穗轴易断㊂百农207成功地整合了亲本的优势,弥补了亲本的不足,具有产量高㊁稳定性好㊁晚春耐倒春寒能力强㊁抗穗发芽㊁千粒重高等特点㊂根据黄淮南片麦区37个地点两年的区域试验和14个地点一年的生产试验(由国家农业技术推广服务中心和河南省农业科学院小麦研究中心联合进行),百农207在2010 2013年的年平均产量为7542~8761kg/hm2,比商品对照品种周麦18增产3.8%~7.0% (表3)㊂2014年,百农207通过国家农作物品种审定委员审定,自2016年起成为河南省推广速度最快的新品种㊂此外,从同一个杂交组合中还筛选出10个具有突出特点的百农207姊妹系(表1),但在产量和其他农艺性状上均未超过百农207㊂表3㊀百农207的产量及主要农艺性状Table3㊀Yields and key agronomic traits of BN207年份试验点数平均产量/(kg㊃hm-2)增产幅度/%穗粒数千粒重/g蛋白质含量/%试验2010 2011208761.50 3.85∗∗34.5041.5014.00黄淮麦区南片区域试验2011 2012177654.50 5.28∗∗36.7041.8015.04黄淮麦区南片区域试验2012 2013147542.007.00∗∗--生产试验∗∗:1%显著性水平2.2㊀百农207的基因组构成及其亲本的贡献共检测了13947个SNP位点,得到7565个(54.2%)有效位点㊂其中百农64和周麦16相同的SNP位点有4009个(52.9%),具有多态性的位点有3556个(47%)㊂在亚基因组中,B基因组SNPs的数量最多, 021㊀第2期马㊀超,等:高产小麦新品种百农207及其姊妹系的基因组构成分析占总SNPs 数量的41.3%,占多态性SNPs 数量的51%,其次是亚基因组A(35.1%,45.4%),而SNP 数量最少的是亚基因组D(20.2%,41.6%)㊂每个染色体上平均有360.2个SNP 位点,但SNP 位点数量在染色体之间变化很大,其中3B 染色体上SNP 位点最多,有694个,其次是2A 染色体,有528个,而4D 染色体上只有108个SNP 位点㊂基于多态性SNP 位点对百农207的基因组构成进行分析,结果表明,父本百农64对百农207基因组的贡献率为55.3%(1968/3556),母本周麦16对百农207基因组的贡献率为40.7%㊂其余142个(3.2%)SNP 位点包括111个(3.1%)杂合位点和31个(0.9%)仅百农207特有的新位点或重组位点㊂此外,在双亲相同的4009个SNP 位点中,检测到33个新SNP 位点,使百农207新位点或重组位点总数增加到64个,占有效SNP 总数(7565个)的0.85%㊂此外,亲本对百农207的亚基因组贡献明显不同,亚基因组B 中绝大多数SNP 位点来自百农64,而亚基因组A 和D 中相对较多的位点则来自周麦16㊂图1为百农207中SNP 位点的染色体分布及亲本贡献率㊂对百农207染色体SNP 位点分布的分析结果表明,其SNPs 来源(父本或母本)存在显著差异㊂用S 代表染色体短臂,L 代表长臂,其中3B㊁1BS㊁2BL㊁2DL㊁4AS㊁5AS㊁6A㊁6B㊁7AL 和7BS 染色体的SNP 位点主要来自父本百农64,而2A㊁3A㊁3D㊁4BS㊁4D㊁5BL㊁6D 和7D 染色体的多数等位基因则来自母本周麦16㊂大多数杂合SNP 位点位于5BL(34/111,30.6%)㊁4BL(20/111,18.0%)和2BS(8/111,7.2%),而大多数重组位点位于5B(25/64,39.1%),其次位于3D(7/64,10.9%)㊂图1㊀百农207中SNP 位点的染色体分布及亲本贡献率Figure 1㊀SNP distribution at chromosomes of BN207and contribution ratios of itsparents 图2㊀基于主成分分析法的百农207及其亲本和姊妹系间的亲缘关系Figure 2㊀Genetic relationship among BN207,its parents and sister lines based on PCA2.3㊀百农207与其姊妹系的遗传构成比较分析百农207与其双亲及10个姊妹系的SNP 位点比较结果表明,百农207与其姊妹系的染色体构成存在显著差异㊂但总体而言,百农207及其姊妹系的1A㊁4A㊁6A㊁3B染色体上的SNPs 绝大多数来自百农64,而2A㊁5A㊁1B㊁6D 和7D 染色体上的SNPs 则大部分是从周麦16遗传来的㊂基于主成分分析法的百农207及其亲本和姊妹系间的亲缘关系如图2所示㊂通过主成分分析法将百农207及其亲本和姊妹系分为3个集群㊂第1个集群包括周麦16和3个姊妹系(百农14-818㊁百农12-40和冠麦1号),第2个集群包括百农64㊁华育198㊁百农10-8㊁华育166㊁PJ11-52和PJ11-15,第3个集群有百农207以及2个姊妹系百旱207和百农69-38,进一步证实百农207和百旱207是所有姊妹系中结合双亲不同性状(SNP 位点)最多的品种㊂121郑州大学学报(理学版)第52卷在姊妹系中,百农14-818与母本周麦16几乎完全相同,只有3.55%的SNP 位点不同㊂在这些SNPs 中,只有1.66%的SNPs 是从百农64遗传来的,而且主要集中位于5B 染色体上(57.6%)㊂因此,本品系可作为周麦16的近等基因系,用于分析百农14-818和周麦16之间性状差异的遗传基础㊂与百农207相比,百旱207耐旱性更好,它与百农207的共有SNP 位点占89.8%㊂这2个品种之间剩余的10.2%的SNP 差异可能有助于研究与百农207的产量优势和百旱207的耐旱性相关的基因㊂此外,PJ11-15品系含有32.0%以上的杂合SNP 位点,说明该品系在遗传上仍不稳定㊂2.4㊀百农207及其双亲和姊妹系的染色体构成分析对33株小麦进行了FISH 分析,百农207及其亲本和姊妹系的核型如图3所示㊂图3中所用材料自上而下依次为:周麦16㊁百农64㊁百农207㊁华育198㊁冠麦1号㊁百旱207㊁PJ11-52㊁PJ11-15㊁百农14-818㊁百农10-8㊁百农12-40㊁百农69-38和华育166㊂探针pAs1-1㊁pAs1-3㊁pAs1-4㊁pAs1-6㊁AFA-3和AFA-4用TAMRA 标记并产生红色信号;pSc119.2-1和(GAA)10用FAM 标记并产生绿色信号㊂白色箭头表示多态性,黄色箭头表示结构重排,白色和黄色星号表示重组㊂图3㊀百农207及其亲本和姊妹系的核型Figure 3㊀Karyotypes of BN207,its parents and sister lines从图3可以看出,亲本百农64和周麦16及其衍生系存在9种染色体变异㊂其中在百农64中检测到6B 染色体的臂间倒位(perInv),与文献[7]的鉴定结果一致㊂另外5种结构变异包括:来自周麦16由小麦长臂1B 和黑麦1R 染色体短臂组成的罗伯逊易位RT1BL.1RS;位于5A 长臂中部的AFA 家族串联重复序列的红221㊀第2期马㊀超,等:高产小麦新品种百农207及其姊妹系的基因组构成分析色信号区段(Trsb);7BL终端区域的pSc119.2-1和(GAA)10重复序列的绿色信号区;7A染色体短臂末端的绿色信号重复序列;1DL末端的绿色信号重复序列㊂其余3种染色体变异包括百农64中3B染色体长臂的中段和4A染色体长臂上各有一个更强的绿色信号,而周麦16的6BS染色体上有一个更强的红色信号(AFA家族重复)㊂染色体核型分析结果表明,百农207中的7A㊁1B㊁3B㊁perInv6B和1D染色体来自百农64,4A和7B染色体来自周麦16,而5A和6B变异来自双亲染色体的重组㊂百农207中5A染色体与百农64相似,但在5A染色体长臂着丝粒附近有较弱的绿色信号;6B染色体与百农64同样有臂间倒位,但在6B染色体短臂的末端区域有较多的红色AFA家族重复,与周麦16一致㊂在10个姊妹系中,有5个(华育198㊁冠麦1号㊁百农14-818㊁百农12-40和百农69-38)含有来自周麦16的RT1BL.1RS染色体,4个(冠麦1号㊁百旱207㊁百农12-40和百农69-38)含有来自周麦16的Trsb5A,只有2个(百旱207和百农10-8)含有来自百农64的perInv6B㊂有3个品系(冠麦1号㊁百农12-40和百农69-38)同时有来自周麦16的Trsb5AL和RT1BL.1RS结构变异,百旱207则含有来自百农64的perInv6B和周麦16的Trsb5A㊂2.5㊀染色体重组区间的物理定位将FISH分析结果与SNP位点的物理位置结合,对染色体重组区域进行了物理定位㊂利用3个含有perInv6B的品系(百农207㊁百旱207和百农10-8),通过对6B染色体上不同亲本来源SNP位点的分析,发现6B染色体最靠着丝粒的位点重组,在短臂上发生在SNP标记AX-109464405(42.5Mb)和AX-110448396 (50.1Mb)之间,而在长臂上发生在AX-109595550(475.5Mb)和AX-111693296(476.2Mb)之间㊂因此,6B 染色体臂间倒位区段被定位在短臂50.1Mb到长臂475.5Mb之间,大小为425.4Mb,占整个6B染色体长度的一半以上㊂含有Trsb5A的4个品系(冠麦1号㊁百旱207㊁百农12-40和百农69-38)的SNP位点分析结果表明, Trsb5A有3个非重组区(重组抑制区)㊂第1个非重组区包括整个短臂,其余2个位于长臂,即包括从着丝粒(109.1Mb)到SNP位点AX-108726870(422Mb)的312.9Mb区段,以及从SNP位点AX-110578543 (480.4Mb)到AX-94589715(546.5Mb)的66.1Mb区段㊂综合双亲间非重组SNP位点的物理位置和FISH 显示重复序列片段位置分析,5A染色体长臂中部的AFA家族串联重复序列区段总长为66.1Mb㊂在母本周麦16中发现了来自其父本周麦8425B的RT1BL.1RS染色体㊂在5个携带RT1BL.1RS的高代品系(华育198㊁冠麦1号㊁百农14-818㊁百农12-40和百农69-38)中,经过百农64来源SNP位点与来自周麦16位点的比较分析,发现黑麦1R短臂与小麦1B短臂之间存在严重的重组抑制现象,并且在1B染色体长臂上也检测到2个不重组区段㊂第1个不重组区段长198.93Mb,从着丝粒(249.2Mb)到SNP位点AX-110687238(448.13Mb)结束,第2个不重组区段长126.35Mb,从位于451.58Mb的SNP位点AX-108725476到位于577.93Mb的AX-110091358之间㊂百农207及其一半姊妹系不含有RT1BL.1RS染色体,表明这种曾经存在于河南及邻省70%以上的小麦品种中的染色体正逐渐失去对现代小麦品种的主导地位㊂2.6㊀百农207抗穗发芽基因的分子标记鉴定百农207是耐穗发芽的硬白小麦品种,与感病对照周麦18相比,萌动种子和发芽种子的比率都较低,表现出较好的抗穗发芽能力,而其姊妹系华育166则和对照周麦18一样,不抗穗发芽(表4)㊂表4㊀百农207穗发芽抗性鉴定Table4㊀Identification of resistance to pre-harvest sprouting of BN207品种名称未发芽率/%萌动率/%发芽率/%平均差异显著性∗平均差异显著性∗平均差异显著性∗百农20769.58ʃ2.72a29.19ʃ2.38b 1.22ʃ0.94c周麦18(对照)46.95ʃ1.75b42.23ʃ1.65a10.79ʃ1.50b华育16636.67ʃ4.45c44.15ʃ5.60a19.18ʃ6.51a ∗:不同字母代表差异达到5%水平的显著性㊀㊀从小麦15K SNP芯片检测到百农207及其双亲百农64和周麦16具有2个穗发芽抗性基因的主效321郑州大学学报(理学版)第52卷QTL 位点,但其单倍型对穗发芽抗性有负效应[16]㊂为了鉴定百农207中与抗穗发芽相关的基因,选用了4个与抗性基因(TaVp1B3㊁PM19-A1㊁Dorm-1和Tamyb10)紧密相关的PCR 标记进行检测㊂百农207及其亲本等小麦品种抗穗发芽基因的PCR 扩增如图4所示(图中箭头表示穗发芽抗性基因的特异性标记)㊂结果表明,百农207及其父本百农64能扩增出与PM19-A1基因连锁的117bp 特异性片段,以及与TaVp1B3基因连锁的569bp 特异性标记,但母本周麦16未能扩增到这些特异性标记片段㊂因此,百农207至少含有2个穗发芽抗性基因,即位于4AL 的PM19-A1基因和3BL 的TaVp1B3基因,并且这2个基因均来自其父本百农64㊂进一步利用34株百农207单株进行抗穗发芽基因TaVp1B3鉴定,结果只有24株(70.6%)扩增到TaVp1B3的569bp 特异性片段㊂因此,百农207后代存在TaVp1B3基因分离,利用该基因特异分子标记继续选择,可以进一步提高百农207对穗发芽的整体抗性水平㊂M:100bp DNA marker;1:中国春;2:百农207;3:百农64;4:周麦16;5~24:周麦16㊁周麦22㊁平安602㊁平安0518㊁扬麦5㊁宁麦9㊁扬麦13㊁扬麦14㊁扬麦16㊁扬麦17㊁扬麦23㊁扬麦158㊁NAU01㊁南农0686㊁NAU617㊁南农9918㊁望水白㊁生选6号㊁偃展4110㊁西农979;a ~d:PM19-A1;TaVp1B3;Dorm-1;Tamyb10图4㊀百农207及其亲本等小麦品种抗穗发芽基因的PCR 扩增Figure 4㊀PCR patterns of pre-harvest sprouting resistance genes of BN207,its parents and other wheat varieties 3㊀小结本研究从染色体和基因组水平分析了百农207及其亲本百农64和周麦16,以及10个姊妹系的遗传组成㊂结果发现,百农207中父本百农64的SNP 贡献率(55.3%)高于母本周麦16(40.7%),染色体间有显著差异㊂其中1BS㊁3BL㊁6AS㊁2AS㊁4BS㊁4DL 染色体上的SNPs 几乎均来源于双亲之一㊂解析百农207及其亲本的染色体和基因组构成,有助于育种家更好地了解这些品种的遗传贡献,快速㊁准确地鉴定品种的真伪,改进育种程序,保护种植户和育种者的合法权益㊂6B 染色体臂间倒位是我国小麦品种中最常见的染色体结构变异类型(15.3%)[8],但目前尚未见对倒位区段进行物理定位的报道㊂本研究通过FISH 和SNP 位点重组分析,将该臂间倒位定位在425.4Mb 区间,从短臂的50.1Mb 到长臂的475.5Mb㊂此外,将来自周麦16染色体5A 长臂上的AFA 家族串联重复序列区段定位在SNP 位点AX-110578543(480.4Mb)到AX-94589715(546.5Mb)的66.1Mb 区间,为perInv 6B 和Trbs 5A 结构变异提供了直接证据㊂本研究利用TaVp1B3特异性标记对百农207单株进行穗发芽抗性基因筛选,在群体中仅鉴定出70.6%421521㊀第2期马㊀超,等:高产小麦新品种百农207及其姊妹系的基因组构成分析的阳性单株,表明百农207中存在TaVp1B3基因分离,筛选具有TaVp1B3基因的单株可以进一步提高百农207对穗发芽的抗性㊂总之,通过全基因组高密度SNP和FISH的联合分析,揭示了小麦品种百农207的染色体构成㊁基因组构成以及双亲的贡献,从染色体和分子2个层次证实百农207较大程度结合了双亲的遗传物质,并首次对染色体变异perInv6B和Trsb5A的重组区段进行了物理定位㊂结果表明,将高通量基因分型技术与FISH技术相结合,能够更准确地评价小麦品种的基因组来源,为新品种选育和优良育种亲本利用提供更全面的研究基础㊂参考文献:[1]㊀RASHEED A,MUJEEB-KAZI A,OGBONNAYA F C,et al.Wheat genetic resources in the post-genomics era:promise andchallenges[J].Annals of botany,2018,121(4):603-616.[2]㊀GODFRAY H C J,BEDDINGTON J R,CRUTE I R,et al.Food security:the challenge of feeding9billion people[J].Science,2010,327(5967):812-818.[3]㊀FEYERHERM A M,PAULSEN G M,SEBAUGH J L.Contribution of genetic improvement to recent wheat yield increases inthe USA[J].Agronomy journal,1984,76(6):985-990.[4]㊀庄巧生.中国小麦品种改良及系谱分析[M].北京:中国农业出版社,2003.ZHUANG Q S.Chinese wheat improvement and pedigree analysis[M].Beijing:China Agriculture Press,2003. [5]㊀LI Z S,LI B,ZHENG Q,et al.Review and new progress in wheat wide hybridization for improving the resistance to biotic andabiotic stresses[M]ʊOGIHARA Y,TAKUMI S,HANDA H.Advances in wheat genetics:from genome to field.Tokyo: Springer,2015:377-385.[6]㊀JIANG P,ZHANG P P,ZHANG X,et al.Genetic contribution of Ningmai9wheat to its derivatives evaluated by using SNPmarkers[J].International journal of genomics,2016:1-6.[7]㊀HUANG X Y,ZHU M Q,ZHUANG L F,et al.Structural chromosome rearrangements and polymorphisms identified in Chinesewheat cultivars by656high-resolution multiplex oligonucleotide FISH[J].Theoritical and applied genetics,2018,131(9): 1967-1986.[8]㊀王竹林,王德森,何中虎,等.小麦品种百农64慢白粉病抗性QTL的定位[J].中国农业科学,2006,39(10):1956-1961.WANG Z L,WANG D S,HE Z H,et al.QTL mapping for slow mildewing resistance in Chinese wheat cultivar Bainong64 [J].Scientia agricultura sinica,2006,39(10):1956-1961.[9]㊀JIN H,WEN W E,LIU J D,et al.Genome-wide QTL mapping for wheat processing quality parameters in a Gaocheng8901/Zhoumai16recombinant inbred line population[J].Frontiers in plant science,2016,7:1-16.[10]BRADBURY P J,ZHANG Z,KROON D E,et al.TASSEL:software for association mapping of complex traits in diverse sam-ples[J].Bioinformatics,2007,23(19):2633-2635.[11]YANG Y,MA Y Z,XU Z S,et al.Isolation and expression characterization of novel Vp-1genes in wheat varieties with distinctPHS tolerance and ABA responsiveness[J].Journal of experimental botany,2007,58(11):2863-2871.[12]曹雪连,张衡,姜昊,等.分子标记PM19-A1对1015份小麦抗穗发芽基因型的筛选及其有效性验证[J].麦类作物学报,2016,36(10):1283-1290.CAO X L,ZHANG H,JIANG H,et al.Detection and validation of molecular marker PM19-A1associated with pre-harvest sprouting resistance in1015wheat varieties[J].Journal of triticeae crops2016,36(10):1283-1290.[13]张春利.小麦抗穗发芽分子标记的发掘与验证[D].北京:中国农业科学院,2008.ZHANG C L.Development and validation of functional marker for resistance to pre-harvest sprouting[D].Beijing:Chinese Academy of Agricultural Sciences,2008.[14]HIMI E,MAEKAWA M,MIURA H,et al.Development of PCR markers for Tamyb10related to R-1,red grain color gene inwheat[J].Theoretical and applied genetics,2011,122(8):1561-1576.[15]LIU W X,KOO D,XIA Q,et al.Homoeologous recombination-based transfer and molecular cytogenetic mapping of powderymildew-resistant gene Pm57from Aegilops searsii into wheat[J].Theoretical and applied genetics,2017,130(4):841-848.[16]ZHOU Y,TANG H,CHENG M P,et al.Genome-wide association study for pre-harvest sprouting resistance in a large germ-plasm collection of Chinese wheat landraces[J].Frontiers in plant science,2017,8:1-13.621郑州大学学报(理学版)第52卷Genomic Composition Analysis of a Novel High-yielding WheatCultivar BN207and Its Sister LinesMA Chao1,WU Hao2,OU Xingqi3,LI Xinhua4,QIAO Hong4,QIN Guangyong4,LIU Wenxuan1,QI Zengjun2(1.College of Life Sciences,Henan Agricultural University,Zhengzhou450002,China;2.State Key Laboratory of Crop Genetics and Germplasm Enhancement,NanjingAgricultural University,Nanjing210095,China;3.School of Life Science and Technology,Henan Institute of Science and Technology,Xinxiang453003,China;4.School of Agricultural Sciences,Zhengzhou University,Zhengzhou450001,China) Abstract:Chromosomal and genomic compositions of BN207,a new dominant wheat variety in the south-ern region of Huang-Huai river valley wheat zone,and its parents BN64and ZM16,as well as ten sister lines were analyzed through an integrated analysis using fluorescent in situ hybridization(FISH)and wheat15K SNP array.The results showed that55.3%and40.7%of the genome of BN207were contrib-uted by BN64and ZM16respectively,and other64loci were novel or recombined in BN207.Besides,a total of nine chromosomal variation types were detected in all tested lines,including five from BN64,four from ZM16or from the recombination of parentsᶄchromosomes.By integration of FISH and SNP loci re-combination analyses,the region of the pericentric inversion of chromosome6B was physically mapped in the interval of50.1Mb at short arm to475.5Mb at the long arm,and the large tandem repeat sequence block at the long arm of5A being located to a region of66.1Mb.Key words:wheat variety;BN207;genomic composition;FISH;SNP locus;chromosomal variation(责任编辑:孔㊀薇)(上接第117页)Preparation and Identification of Monoclonal Antibodies Against M Protein of Avian Infectious Bronchitis VirusZHOU Jingming,MA Wenli,QI Yanhua,MA Qiang,ZHANG Gaiping,WANG Aiping(School of Life Sciences,Zhengzhou University,Zhengzhou450001,China) Abstract:The infectious bronchitis virus(IBV)M41strain was propagated in chicken embryos.Total RNA was extracted from the allantoic fluid.M gene was amplified by RT-PCR to construct the prokaryotic expression vector pGEX-6p-1-M,through which the recombinant M protein was expressed in E.coli Transetta(DE3).BALB/c mice were immunized with the purified recombinant M protein.Anti-M pro-tein monoclonal antibody hybridoma cells were prepared by hybridoma technique.Anti-M protein mono-clonal antibodies were prepared by in vivo induction ascites method.Ascites was purified by caprylic acid-ammonium sulfate method,and the purified monoclonal antibody was identified.Key words:infectious bronchitis virus;M protein;prokaryotic expression;monoclonal antibody(责任编辑:孔㊀薇)。


【最新资料,Word版,可自由编辑!】施工安全总体风险评估报告一、编制依据1、《公路桥梁和隧道工程施工安全风险评估指南》(试行)交质监发【2011】217号;2、《S323鸿门至旌德公路梅岭隧道及接线工程施工合同文件》、二阶段施工图设计;3、交通部颂发的《公路工程标准施工招标文件(2009年版)》、现行《公路工程技术标准》、现行《公路隧道施工技术规范》、现行《公路工程施工安全技术规程》等相关规范;4、《公路施工手册》、现行《工程建设标准强制性条文·公路工程部分》;5、现场踏勘调查、搜集的实地资料;6、我单位在类似工程中的施工经验和相关工程的技术总结、工法成果等。
7、依据以上文件、规范、标准及工程实地勘察情况,结合我公司现有技术装备、施工能力、管理水平,以及多年从事复杂地形地质条件隧道施工的丰富经验,并针对本工程施工特点,以“保质量、保工期、保安全、创精品”为目标,编制本梅岭隧道施工安全总体风险评估报告。
二、工程概况(一)、隧道工程概况S323鸿门至旌德公路梅岭隧道行政区域隶属于安徽省宣城市宁国市。
梅岭隧道进口位于宁国市胡乐镇梅树下村原曙光厂附近梅岭山脚,出口位于宁国市胡乐镇梅岭脚村附近梅岭山脚。
隧道进洞口离S323约300米,有机耕道路前往,出洞即为S323,交通条件较好。
隧道为单洞双车道,隧道长度为939m,起止里程为K7+722~K8+661,属全线重点控制性工程。
(二)、桥梁工程概况本桥位于鸿门至旌德公路,行政区域隶属于安徽省宣城市宁国市胡乐镇梅岭脚村附近。
本桥上部结构为5×16m现浇普通钢筋砼连续空心板,下部结构桥墩采用柱式桥墩下接扩大基础,桥台采用重力式U型桥台。
(三)、隧道设计技术标准1、公路等级:二级公路;2、隧道设计行车速度:40km/h;3、隧道建筑限界:净宽10.0m,净高5.0m;4、洞内路面设计荷载:BZZ-100;5、行车方式:双向行车;6、通风方式:机械通风;7、隧道防水等级:二级;二次衬砌砼抗渗等级不小于S6。

财政部住建部:关于进一步推进公共建筑节能工作的通知财建[2011]207号各省、自治区、直辖市、计划单列市财政厅(局)、住房城乡建设厅(委),新疆生产建设兵团财务局、建设局:近年来,按照国务院节能减排综合性工作方案的统一部署,财政部、住房城乡建设部在全国范围内开展国家机关办公建筑和大型公共建筑的能耗统计、能源审计、能效公示工作,在部分省市开展公共建筑能耗动态监测平台建设试点,取得了良好效果,为节能量审核、制定能耗定额、建立能效交易机制提供有力支撑,充分激发了节能改造市场需求。
但当前还存在大型公共建筑能耗水平高、增长势头猛、节能改造进展缓慢等突出问题。
为切实加大组织实施力度,充分挖掘公共建筑节能潜力,促进能效交易、合同能源管理等节能服务机制在建筑节能领域应用,财政部、住房城乡建设部将进一步开展公共建筑节能工作,现就有关事项通知如下。
一、明确“十二五”期间公共建筑节能工作目标建立健全针对公共建筑特别是大型公共建筑的节能监管体系建设,通过能耗统计、能源审计及能耗动态监测等手段,实现公共建筑能耗的可计量、可监测。
确定各类型公共建筑的能耗基线,识别重点用能建筑和高能耗建筑,并逐步推进高能耗公共建筑的节能改造,争取在“十二五”期间,实现公共建筑单位面积能耗下降10%,其中大型公共建筑能耗降低15%。
二、加强新建公共建筑节能管理(一)严格执行节能标准。
新建公共建筑应按照节能省地及绿色生态的要求指导工程建设全过程,要严格执行工程建设节能强制性标准,把能耗标准作为建筑项目核准和备案的强制性门槛,遏制高耗能建筑的建设。
新建公共建筑要大力推广绿色设计、绿色施工,广泛采用自然通风、遮阳等被动节能技术。
(二)实行建筑能耗指标控制。
要强化公共建筑特别是大型公共建筑建设过程的能耗指标控制,应根据建筑形式、规模及使用功能,在规划、设计阶段引入分项能耗指标,约束建筑体型系数、采暖空调、通风、照明、生活热水等用能系统的设计参数及系统配置,避免建筑外形片面追求“新、奇、特”,用能系统设计指标过大,造成浪费。
综采工作面“三机”配套几何关系分析王喜贵(中煤平朔煤业有限责任公司,山西平朔036006)[摘要]根据综采工作面中部“三机”几何配套,研究了采煤机与液压支架、采煤机与输送机、液压支架与输送机之间的中部配套关系,分析了采煤机在输送机弯曲段以及工作面机头、机尾处“三机”配套关系。
[关键词]采煤机;输送机;液压支架;几何配套关系[中图分类号]TD421.8;TD355.41[文献标识码]B [文章编号]1006-6225(2011)04-0078-02Geometry Relationship Analysis of Mining Equipments in Full-mechanized Mining Face[收稿日期]2011-01-19[作者简介]王喜贵(1964-),男,山西平朔人,高级工程师,中煤平朔煤业有限责任公司副总经理。
综采面设备配套主要是解决生产能力和几何关系配套两方面问题。
几何关系配套是要解决转载机、带式输送机、采煤机、输送机等设备间的连接关系,确定合理的配套尺寸,其中以输送机、采煤机、支架(“三机”)的相互配套关系最为重要,其几何关系配套主要是确定输送机、采煤机和支架的相互搭接尺寸,保证设备运行时不发生干涉。
1综采工作面中部“三机”几何关系分析“三机”几何关系配套尺寸主要有铲板间距e 、梁端距D 、机面高度A 、采高H 、过机空间C 、过煤高度E 等尺寸,如图1所示。
下面着重对这些配套尺寸的选取原理进行分析。
图1工作面中部“三机”几何尺寸关系配套1.1采煤机与液压支架的配套关系采煤机和液压支架之间没有直接连接关系,但有两个位置关系需要给予重视。
(1)梁端距D 梁端距主要依据煤的软硬程度、顶板冒落情况、底板沿走向起伏变化情况以及采高等因素来选取,过大将会加长支架控顶距,过小易造成采煤机滚筒割到顶梁。
(2)过机空间C采煤机沿工作面往返割煤时,其机身要在支架顶梁下通过,应保证两者在工作时不发生干涉。
现代伊兰特2011款参数配置:排量:1.6L变速器型式:5档(手动)、4档(自动)综合工况耗油:6.9L/100km(自动)、7.3L/100km(自动)最小转弯半径:5.06m驱动方式:前轮驱动整备质量:1250kg最高车速:187km/h(手动)、173km/h(自动)外部配置:车身颜色:铂金银,漫红,海黄蓝,贵族白,乌木黑,星辉银车窗:前后电动窗侧转向灯:外后视镜电动窗锁止功能、外后视镜电动调节、前雾灯、前照灯照射范围调整、侧转向灯、高位制动灯、LED尾灯内饰配备:2011款伊兰特沿用了老款的设计及功能,使用上深下浅的传统配色,一切均以实用为主;中控台部分略向驾驶员方向倾斜,操作上方便司机使用,也间接地将前排划分出主副驾驶区域。
影音空调:CD、扬声器(6)、空调、音频格式支持MP3便利功能:倒车雷达(车后)、中控门锁、遥控钥匙、遮阳板化妆镜安全配置:刹车防抱死(ABS)、制动力分配(EBD/CBC/EBV)、电子防盗系统、发动机防盗系统、驾驶安全气囊、副驾驶安全气囊、前安全带调节、安全带预收紧和限力功能、后排安全带、儿童锁、车门防撞杆(防撞侧梁)、可溃缩转向柱、安全带未系提示。
新款外观介绍1、整体:采用了“流体雕塑”作为新车的主要设计语言。
饱满的车头和六边形中网成为了新伊兰特的主要特征。
漂亮的大灯与尾灯之间通过充满力量感的Z字形腰线进行连接,塑造出新车性感的侧面造型,风格更加偏向于运动风格,运动型后视镜和铝合金运动轮毂的配备都让这款新车散发出运动气息2、正面:前水箱格栅将粗条中间的小细条也镀上了铬,使得整辆车的前格栅看起来更加的亮丽,也产生了一种厚重的感觉,让以前看起来脑袋比较小的伊兰特一下子就显得大气起来。
前大灯组也有了一定的改变,原来的转向灯使用的是白色的灯泡和黄色的灯罩。
而换了新装的伊兰特则正好相反,将灯泡换成了黄色而灯罩则换成了白色,这样整个前大灯的组合就如钻石般的闪亮,却不会影响转向灯的使用3、尾部:虽然新款伊兰特的尾灯形状并未改动,但大灯的内部结构已发生了改变,全新设计的保险杠配以修长的柳叶状雾灯,面积更大的后牌照上方的镀铬装饰条,有了亮条的衬托,整个车尾部的立体感马上就表现出来了,使整体造型更为运动,尾部还增加了一个VVT标志4、侧面:在大灯与尾灯之间采用Z字形腰线设计,使得车身侧面造型更加性感各部位材质:方向盘表面材料:PVC塑料坐椅:采用浅灰色织物材质,并带有透气孔前照灯类型:卤素盾形中控面板:由钢琴烤漆以及哑光金属效果的材质组合而成缸体材料:铁缸盖材料:铝轮胎品牌:锦湖轮鼓:标准配备的铝合金轮毂,但是备胎是钢轮毂的门饰件、玻璃黏着剂、门内把手、车内密封胶等都采用了环保材料制造采用的技术:配备1.6CVVT发动机,先进、成熟,搭配5MT、4AT,低油耗伊兰特采用的是前麦弗逊式、后多连杆式悬架系统2011款伊兰特全系标配15英寸铝合金轮毂及185/65 R15的轮胎。
目录一、总说明 (1)二、报表目录 (2)三、调查表式(一)综合年报、定期报表式1.能源平衡表(实物量)(P303—1表) (3)2.能源平衡表(标准量)(P303—4表) (4)(二)基层年报、定期报表式1.工业企业能源购进、消费及库存(P201表) (5)2.工业企业能源购进、消费及库存附表(P201-1表) (6)3.工业企业水消费(P206表) (6)4.主要耗能工业企业单位产品能源消耗情况 (P207表) (7)5.重点用能工业企业单位产品能源消耗情况(鲁P267表) (7)四、附录(一)P201表、P201-1表能源填报目录 (8)(二)主要耗能工业企业单位产品能源消耗情况(P207表)填报目录 (9)(三)重点用能工业企业单位产品能源消耗情况(鲁P267表)填报目录 (14)(四)主要指标解释 (15)(五)主要耗能工业企业单位产品能源消耗情况(P207表)指标的计算方法 (23)(六)重点用能工业企业单位产品能源消耗情况(鲁P267表)指标的计算方法 (42)(七)附表一:钢铁企业吨钢可比能耗计算方法 (57)(八)附表二:各种能源折标准煤参考系数 (58)一、总说明(一)为了解青岛市能源供应、需求的基本情况,为各级政府制定政策、计划,进行经济管理与调控提供依据,依照《中华人民共和国统计法》的规定,特制定本统计报表制度。
(二)本报表制度是国家统计调查的一部分,是青岛市统计局对各区、市统计局和市直有关企业集团(公司)的综合要求。
各区市统计局、各部门应按照全市统一规定的统计范围、计算方法、统计口径和填报目录,根据青岛市统计局拟订的能源统计报表制度的内容,认真组织实施,按时报送。
地方、部门特殊需要的统计资料应通过地方统计调查和部门统计调查搜集,并尽量避免与青岛市统计局调查内容相重复。
(三) 本报表制度主要由综合年报表、综合定期报表、基层年报表、基层定期报表构成,这些报表反映能源的生产、进出口、库存、购进、消费和能源单耗等情况。
小麦高产品种1、泛麦803:半冬性中熟品种,全生育期229.0-233.6天,在2012-2013年亩产500.2千克。
2、百农207:为半冬性中晚熟品种,全生育期231天,在2010-2011年亩产584.1千克。
3、中麦875:为半冬性中晚熟品种,在2010-2011年亩产574.8千克。
一、泛麦8031、特征特性(1)该品种属于半冬性中熟品种,全生育期229.0-233.6天。
(2)幼苗半匍匐,叶片宽短,叶色深绿。
冬季抗寒性较好,分蘖力一般,成穗率高,春季起身拔节早,但两极分化慢,抽穗偏晚。
(3)该品种株型较为紧凑,旗叶上举,干尖,茎叶蜡质较重,穗下节短,穗层整齐,株高73.0-77.9cm,茎秆弹性弱,抗倒性一般。
(4)该品种的穗为近长方型,小穗排列较密,长芒,白壳,白粒,籽粒半角质,饱满度一般。
根系活力好,耐后期高温,熟相好。
(5)该品种的亩成穗数为40.8-41.9万,穗粒数为33.6-35.6粒,千粒重为40.0-48.3克。
2、产量(1)该品种在2012-2013年度河南省冬水C组区域试验,平均增产点率84.6%,平均亩产500.2千克,比对照品种周麦18增产1.9%,居15个参试品种第4位。
(2)该品种在2013-2014年度河南省冬水C组区域试验,平均亩产583.1千克,比对照品种周麦18增产3.2%,居15个参试品种第7位。
(3)在2014-2015年度河南省冬水A组生产试验,平均亩产546.3千克,比对照品种周麦18增产7.0%,居8个参试品种第3位。
二、百农2071、特征特性(1)该小麦为半冬性中晚熟品种,全生育期231天,比对照周麦18晚熟1天。
(2)该小麦幼苗呈半匍匐状,长势旺,叶宽大,叶深绿色。
冬季抗寒性中等。
(3)该小麦分蘖力较强,分蘖成穗率中等。
早春发育较快,起身拔节早,两极分化快,抽穗迟,耐倒春寒能力中等。
中后期耐高温能力较好,熟相好。
(4)该小麦株高约76厘米,株型松紧适中,茎秆粗壮,抗倒性较好。