虾夷扇贝生活史
- 格式:doc
- 大小:4.07 MB
- 文档页数:4

辽宁省海洋水产研究所引进虾夷扇贝
秦传信
【期刊名称】《现代渔业信息》
【年(卷),期】1987(000)006
【摘要】获悉,辽宁省海洋水产研究所于1980—1982年先后4次从日本引进虾夷扇贝亲贝,进行人工育苗获得成功。
最近几年经过科学试验,突破了虾夷扇贝人工育苗和增养殖的一系列技术关键。
育苗和养殖试验结果表明,每立方水体培育稚贝为70万个,壳高600微米,每台筏挂养9,00个饲养一年,
【总页数】1页(P27)
【作者】秦传信
【作者单位】无
【正文语种】中文
【中图分类】S968
【相关文献】
1.辽宁省海洋水产研究所利用本所育成的第一代二龄虾夷扇贝采苗成功 [J], 王庆成
2.辽宁省海洋水产研究所《刺参人工育苗研究》通过技术鉴定 [J], 辽林
3.辽宁省海洋水产研究所完成海蜇生活史的研究 [J], 吕水
4.浙江省海洋水产研究所:担当海洋渔业高质量发展和渔场修复振兴前哨 [J], 许文军
5.辽宁省海洋水产科学研究院加挂辽宁省海洋环境监测总站 [J], 王年斌
因版权原因,仅展示原文概要,查看原文内容请购买。

虾夷扇贝的引进及其在我国北方增养殖的前景
王庆成
【期刊名称】《水产科学》
【年(卷),期】1984(000)004
【摘要】虾夷扇贝(Pecten yessoesis)属软体动物门瓣鳃纲扇贝科,为扇贝科动物中个体最大(最大个体壳长16厘米、重450克)、产量最多的种类,其肉味鲜美、营养价值高。
贝柱部含蛋白质20.8%,脂肪0.8%,碳水化合物2.4%,灰分1.8%,贝柱部含蛋白质占软体部湿重量的35%。
生长快,自采苗算起,养殖17-23个月的个本平均壳长可达10厘米,重140-150克。
【总页数】4页(P24-27)
【作者】王庆成
【作者单位】辽宁省海洋水产研究所
【正文语种】中文
【中图分类】S968
【相关文献】
1.我国北方地区奶牛规模化养殖现状及发展前景探析 [J], 王兰;杨波
2.虾夷扇贝增养殖中存在的问题和对策 [J], 高学文
3.漠斑牙鲆在我国北方养殖前景的探讨 [J], 梁毅锋
4.持续发展虾夷扇贝的健康增养殖 [J], 李文姬;薛真福
5.虾夷扇贝引种育苗和增养殖试验成功并通过技术鉴定 [J], 寇宝增
因版权原因,仅展示原文概要,查看原文内容请购买。


不同壳色虾夷扇贝家系F1幼虫生长及遗传结构的比较分析程鹏;杨爱国;周丽青;吴彪;刘志鸿;李霞;李德军【期刊名称】《中国水产科学》【年(卷),期】2010(017)005【摘要】虾夷扇贝(Patinopecten yessoensis)群体存在2种壳色个体,简称白贝和褐贝.本研究通过近交和杂交方法建立了褐贝(♀)×褐贝(♂)、白贝(♀)×褐贝(♂)、褐贝(♀)×白贝(♂)和白贝(♀)×白贝(♂)4个家系,分别用1F、2F、3F和4F代表,每家系设3个平行,共12个家系.对各家系的卵径、幼虫的孵化率、成活率和生长性状等生物学参数进行了比较,建立了各个家系幼虫壳长、壳高的生长方程,并利用微卫星标记比较4个家系的遗传差异.结果表明,不同壳色家系间卵径大小、幼虫的孵化率、成活率差异不显著(P>0.05),家系4F幼虫期壳长和壳高日生长速率均高于1F、2F和3F,且差异显著(P<0.05),家系2F、3F和4F间幼虫期壳长和壳高日增长速率差异不显著(P>0.05).家系1F的遗传多样性最低;遗传相似性系数(0.635 1~0.977 2)和遗传距离(0.023 1~0.454 0)分析表明,1F和4F之间遗传相似性(0.635 1)最低.遗传距离(0.454 0)最远;不同家系间存在着遗传差异,1F和4F差异最大.本研究结果初步说明虾夷扇贝群体内白贝个体和褐贝个体之间产生了遗传分化,这种遗传结构的差异造成了不同壳色家系在幼虫阶段生长性状上的不同.【总页数】9页(P960-968)【作者】程鹏;杨爱国;周丽青;吴彪;刘志鸿;李霞;李德军【作者单位】农业部海洋渔业资源可持续利用重点开放实验室,中国水产科学研究院,黄海水产研究所,山东,青岛,266071;大连水产学院,生命科学与技术学院,辽宁,大连,116023;农业部海洋渔业资源可持续利用重点开放实验室,中国水产科学研究院,黄海水产研究所,山东,青岛,266071;农业部海洋渔业资源可持续利用重点开放实验室,中国水产科学研究院,黄海水产研究所,山东,青岛,266071;农业部海洋渔业资源可持续利用重点开放实验室,中国水产科学研究院,黄海水产研究所,山东,青岛,266071;农业部海洋渔业资源可持续利用重点开放实验室,中国水产科学研究院,黄海水产研究所,山东,青岛,266071;大连水产学院,生命科学与技术学院,辽宁,大连,116023;胶南市海洋与渔业局,山东,青岛,266400【正文语种】中文【中图分类】S917【相关文献】1.不同壳色虾夷扇贝外套膜形态学及显微结构的研究 [J], 赵乐;丁君;王俊杰;郝振林;常亚青2.微卫星标记在不同壳色虾夷扇贝家系亲权鉴定的适用性 [J], 程鹏;杨爱国;吴彪;周丽青;李霞3.不同壳色杂色鲍F1家系的早期生长及遗传结构比较 [J], 范飞龙;游伟伟;战欣;骆轩;陈楠;柯才焕4.不同壳色皱纹盘鲍杂交家系J1RhF1全长cDNA文库的构建 [J], 刘晓;高其康;张国范5.虾夷扇贝不同壳色间的遗传结构及微卫星标记与生长性状相关分析 [J], 张婧;常亚青;丁君;王晓燕;徐铭键因版权原因,仅展示原文概要,查看原文内容请购买。

虾夷扇贝增养殖中存在的问题和对策建议作者:高学文来源:《河北渔业》 2013年第6期高学文(辽宁省海洋水产科学研究院,辽宁大连 116023)摘要:虾夷扇贝养殖呈现出规模化、产业化的良好发展势头,但也存在着一些不可忽视的问题。
本文分析了虾夷扇贝养殖过程中出现的问题以及产生问题的原因,并提出了解决的对策。
关键词:虾夷扇贝;养殖;问题;原因;对策虾夷扇贝属于冷水性双壳贝类,是扇贝家族中的优良增养殖品种之一,该种原产于日本北部及俄罗斯远东地区沿海水域,我国于上世纪80年代初,由辽宁省海洋水产科学研究院科技人员从日本引种虾夷扇贝,经过生活史研究、人工繁殖培育和驯化推广,其养殖规模不断扩大。
尤其是近年来,随着人工育苗、中间育成、增养殖技术的逐渐成熟,现已成为北方海域海水贝类的主要养殖对象之一,呈现出规模化、产业化的良好发展势头。
但在养殖过程中仍存在着一些不可忽视的问题。
1虾夷扇贝养殖存在的主要问题1.1大量死亡近年来,北方的虾夷扇贝死亡率一直居高不下。
15月龄~20月龄的贝,夏季死亡明显増加。
张明明、赵文研究发现死亡的现象表现为养殖区由外缘向内缘蔓延,底层向中层、表层扩散,底层死亡尤其严重的现象[1]。
1.2人工育苗孵化率低虾夷扇贝育苗技术经过30多年的发展,各项工艺已经逐渐完善。
通过亲贝人工促熟方法,保证了受精卵的质量,有利于幼虫培育;有结聚乙烯网片的使用改善了稚贝生长环境,提高了育成率。
但是,近几年在人工育苗过程中出现了亲贝采卵不集中,受精卵孵化率低,幼虫培养过程中突然大批量下沉死亡等现象[2]。
1.3底播增殖回捕率低底播增殖由于低成本、低风险的特点深受养殖企业喜爱。
同时,由于是最接近于虾夷扇贝自然生活状态的一种增养殖方式,在市场上多被作为野生品种对待,产品销量高、价格贵。
但是,随着底播规模的不断扩大,也暴露出一些问题,例如由于底播后回捕率低导致单位面积产量不高,回捕个体趋向小型化等等。
2虾夷扇贝增养殖出现问题的原因2.1环境污染环境污染、赤潮以及环境的突然变化,导致扇贝死亡。

大连獐子岛海域虾夷扇贝养殖容量张继红;方建光;王诗欢【期刊名称】《水产学报》【年(卷),期】2008(32)2【摘要】现场测定了獐子岛海域的叶绿素a浓度、初级生产力的季节性变化和养殖虾夷扇贝的种群结构;采用生物沉积法,测定了不同规格的虾夷扇贝的滤水率和虾夷扇贝的基本生物学特性.调查结果显示,叶绿素浓度在1.23~2.85 mg·m-3范围内,均值为(1.78±0.57)mg·m-3;初级生产力的平均值为(76.6±41.9)mg C·m-2·d-1,虾夷扇贝单位个体的滤水率为(0.55±0.25)L·h-1.结合虾夷扇贝的年产量、海域面积和有关的水文状况等数据,计算了食物限制性指标的数值,摄食压力和调节比率在0.05和1.0之间,而滤水效率小于0.05.结果显示,由于该海域的叶绿素浓度和初级生产力水平较低,海水流速大,因此,水交换带来的悬浮颗粒物为主要食物来源.目前的虾夷扇贝的养殖量(32亿粒)接近生态容量,如果开展筏式养殖,利用整个海域的水体,养殖量增大20倍的空间并达到养殖容量,预计年产量将达到256亿粒.【总页数】6页(P236-241)【作者】张继红;方建光;王诗欢【作者单位】中国水产科学研究院黄海水产研究所,山东,青岛,266071;中国水产科学研究院黄海水产研究所,山东,青岛,266071;大连獐子岛渔业集团股份有限公司,辽宁,大连,116001【正文语种】中文【中图分类】S968.31+6.9【相关文献】1.大连獐子岛人工鱼礁海域夏季水质变化与评价 [J], 刘永虎;程前;田涛;陈勇;尹增强;刘汉超;王月;2.大连獐子岛人工鱼礁海域夏季水质变化与评价 [J], 刘永虎;程前;田涛;陈勇;尹增强;刘汉超;王月3.虾夷扇贝的摄食生理及其在獐子岛海域的群体滤水能力 [J], 张继红;方建光;王巍;蒋增杰;臧有才;王诗欢;张媛4.獐子岛养殖海域颗粒有机碳、颗粒氮的时空分布特征 [J], 刘毅;张继红;杜美荣;蔺凡;丁刚;吴文广;房景辉5.北黄海獐子岛养殖海域营养水平与虾夷扇贝增殖渔获量评估 [J], 段丽琴;宋金明;袁华茂;李学刚;李宁;马继坤因版权原因,仅展示原文概要,查看原文内容请购买。


虾夷扇贝幼体资源采集技术研究
周志刚
【期刊名称】《科学养鱼》
【年(卷),期】2018(0)10
【摘要】虾夷扇贝为品质优良的冷水性双壳贝类,原产于日本北部沿海。
在日本,虾夷扇贝海区于20世纪30年代进行了试验性采苗,逐渐形成了成熟的采苗技术,目前全日本虾夷扇贝的增养殖苗种全部依赖天然采苗。
虾夷扇贝自20世纪80年代初引入我国以来,其苗种生产方式主要依赖于人工育苗。
随着增养殖规模的不断扩大,虾夷扇贝繁殖季节在我国大连地区南部沿海形成较大的幼体资源,经常能看到附着在海带叶片、贝类养殖笼中的虾夷扇贝稚贝,有些海区数量可观,为虾夷扇贝海区采苗提供了条件。
【总页数】3页(P48-50)
【作者】周志刚
【作者单位】辽宁旅顺口区水产技术推广站 116041
【正文语种】中文
【相关文献】
1.1株虾夷扇贝苗种培育用益生菌的筛选及其对幼体存活和生长的影响 [J], 赵学伟;李明;梁峻;刘继晨;孙雪莹;马悦欣;
2.1株虾夷扇贝苗种培育用益生菌的筛选及其对幼体存活和生长的影响 [J], 赵学伟;李明;梁峻;刘继晨;孙雪莹;马悦欣
3.虾夷扇贝育苗阶段幼体细菌性病原的确定 [J], 刘继晨;金晓杰;李明;陈娟;郭宗明;
马悦欣
4.黄海北部大连沿岸虾夷扇贝天然苗采集技术研究 [J], 常亚青;王庆志;宋坚;曹学彬;李石磊
5.4种多环芳烃对虾夷扇贝(Patinopecten yessoensis)幼体的急性毒性研究 [J], 刘帅;刘欢;魏海峰;赵肖依;宋雪
因版权原因,仅展示原文概要,查看原文内容请购买。

虾夷扇贝对温度饥饿胁迫的应答机制浅析虾夷扇贝是一种重要的海洋经济贝类,它们广泛分布于北太平洋海域。
虾夷扇贝的生长和繁殖很大程度上受到环境因素的影响,其中温度和饥饿是最主要的生物学因素之一。
近年来,随着全球气候变化的加剧,海洋水温的波动日益剧烈,虾夷扇贝的生存环境也受到了不小的挑战。
因此,研究虾夷扇贝对温度和饥饿胁迫的应答机制,对于保护和利用虾夷扇贝资源具有极其重要的意义。
虾夷扇贝的生物学特性虾夷扇贝主要分布于北太平洋海域,包括日本、中国、俄罗斯等地。
它们生活在寒冷的海域底层,常年受到温度和饥饿的双重影响。
虾夷扇贝的生活史包括卵、幼体、稚贝、壳长1~2 cm的小扇贝以及壳长3 cm以上的成年扇贝。
成年扇贝通常在3~4年后性成熟,每年发生一次繁殖,产卵期一般在7月至8月间,卵径在0.1~0.2mm之间,孵化出来的幼体称为浮游幼体。
虾夷扇贝对温度的适应范围比较窄,一般在5~20摄氏度之间生长最为适宜。
在此范围之外,虾夷扇贝的生长和繁殖能力都会受到明显影响。
研究表明,与高温相比,低温对虾夷扇贝的影响更为严重。
低温会对虾夷扇贝的肌肉和胃肠系统产生不良影响,进而导致食欲下降,身体健康状况下降。
同时,低温还会影响虾夷扇贝的代谢和呼吸功能,导致能量代谢下降和生长停滞。
虾夷扇贝对温度的应答机制主要是通过基因表达调控来实现的。
研究表明,在高温下,虾夷扇贝的核糖体生物合成、核苷酸代谢以及蛋白质代谢等基因表达会明显上调,而在低温下,这些基因表达则会明显下调。
此外,低温还会导致虾夷扇贝的转录机制发生改变,进而影响到虾夷扇贝的生长和发育。
饥饿是虾夷扇贝生存的一个重要问题。
在野外环境中,虾夷扇贝常常会面临食物不足的情况,从而影响它们的生长和繁殖,甚至导致死亡。
虾夷扇贝对饥饿的应答机制主要是通过生理和代谢调节来实现的。
在饥饿状态下,虾夷扇贝的营养代谢会发生明显变化,包括体内能量代谢、蛋白质分解和合成、和糖代谢等方面。
此外,饥饿还会影响虾夷扇贝的消化系统和免疫系统。

扇贝生长特性介绍(1)栉孔扇贝的生长。
栉孔扇贝的生长有明显的季节性,一年之中有两个快速生长期,它们是5-7月(水温15℃--25℃)和9-11月(水温25℃- 15℃)。
此时,月平均生长速度分别为0.8-lcm和0.5-0.8cm。
夏季水温超过25℃时生长缓慢,月平均增长0.3-0.5cm;冬季水温低于5℃时停止生长。
栉孔扇贝在壳高4.5cm以前主要生长贝壳,软体增重缓慢,此时每增高lcm,软体增重约4.4g;壳高4.5cm以后,贝壳生长变慢,软体生长较快,此时壳高每增长lcm软体可增重12.6g。
在人工筏式养殖的情况下,当年人工育的苗,入冬前壳高可达2-2.5cm;1龄贝有80%以上的个体壳高达到4cm左右;1.5龄贝(即翌年11- 12月)有70%以上的个体壳高超过7cm。
人工养殖的栉孔扇贝比自然生长的要快一年。
(2)海湾扇贝的生长。
海湾扇贝是一年生贝类,生长较快。
4月份人工培育的苗种,到11月中下旬,平均壳高达5.3cm,平均重量34.5g。
在水浅温度高的海区,海湾扇贝生长较快,在6-9月高温期壳高月增长约lcm,进入11月份生长变慢,水温5℃以下停止生长。
生长一周年后,80%以上的个体死亡。
(3)虾夷扇贝的生长。
虾夷扇贝每年有两个生长季节,一是4-7月,平均月增高lcm左右;二是10 - 12月,平均月增高0.5-0.8cm,8-9月高温季节停止生长,夏季水温连续15 - 20天超过23℃就会死亡。
虾夷扇贝1龄贝壳高5-6cm,2龄8-l0cm,3龄10-13cm,4龄13 -15cm。
经人工筏式养殖17-23个月,平均壳高可达l0cm以上,体重140 - 150g,出肉率39% -42%,闭壳肌直径2.8- 3.6cm,闭壳肌平均鲜重14 - 18g,出品率约为12%。
一种虾夷扇贝海上分苗的方法我折腾了好久虾夷扇贝海上分苗这事儿,总算找到点门道。
最开始的时候,我真的是瞎摸索。
我就知道虾夷扇贝得在合适的时候分苗,但啥是合适的时候呢,我一开始可不懂。
我就看着苗的大小,感觉差不多大了就想分苗。
我第一次尝试的时候,没选好天气。
海上风浪可大了,那船晃悠得人都站不稳。
我带人去分苗,结果苗刚拿出来就被晃得散得到处都是,很多苗都受损了,那一次简直失败得一塌糊涂。
后来我就长记性了,天气非常重要。
一定要挑那种风平浪静的天气,就像你要在平平稳稳的桌子上摆弄小物件一样,海上也得平静。
选好日子以后,就开始准备工具。
这就跟你做饭得先备好厨具一样,分苗得有专门的分苗器械,可不能瞎凑合。
然后就是挑选合适的海域来分苗。
这个我试过好多地方,有的海域水太浑,我感觉扇贝在那里可能会不舒服,分苗后的生长效果就不太好。
有的海域水流特别急,苗刚放下去就被冲走了。
所以最后我就确定了一个水流比较平缓,水质干净清澈的海域,就像你给孩子选宜居的地方一样,得找那种特别适宜的环境。
在实际分苗操作的时候,我也可小心了。
苗不能暴力对待,得轻轻拿轻轻放,就像照顾娇嫩的花朵一样。
而且每一批苗的数量要控制好,别放太多,不然它们就太挤了,就像一堆人住在很小的屋子里,难受得很。
我一开始不确定放多少合适呢,就做实验。
我放不同数量的苗在小范围的海域,过段时间去看看哪个区域里的扇贝长得最好,这些实际的操作都非常有用。
还有就是分苗的时间,我发现白天暖和的时候,苗比较活跃,分苗就好像比较顺利。
但是也不能太热的时候分,太晒的话,那些苗暴露在空气中很快就不好了。
这个温度的把握我还在不断摸索,大体感觉像是春天或者秋天那种不冷不热的时候就比较好。
我还犯过一个错误呢,就是分苗后没有及时去查看。
我以为只要放下去就万事大吉了,但是后来发现有些苗被海水里的杂质覆盖了,生长受影响。
所以现在我都会定期去查看分苗后的情况,查看到有啥不对劲的地方,就赶紧想办法解决,不能拖,就像人病了得早治一样。
大连旅顺渤海侧沿海虾夷扇贝幼虫来源调查李文姬;薛真福;李华琳;张明;罗鸣;鲁学辉;臧有才;刘项峰【期刊名称】《水产科学》【年(卷),期】2008(27)11【摘要】旅顺渤海侧沿海没有虾夷扇贝资源,但是每年的5月中旬-6月中旬出现虾夷扇贝幼虫,使该海域形成采苗场.2006-2007年笔者通过在虾夷扇贝主要增养殖海区投放漂流瓶跟踪移动方向,监测浮游幼虫数量及个体大小分布等方法,调查了旅顺渤海侧沿海虾夷扇贝幼虫来源.结果表明,2006年4月30日、2007年4月8日和4月28日,在獐子岛海域投放的漂流瓶,回收率分别为7.5%、14.0%和17.2%,其中在獐子岛至旅顺渤海侧沿海回收的比例分别为6.0%、10.4%和4.0%.2006年4月6日,大钦岛海域投放的漂流瓶,回收率为13.0%,在旅顺渤海侧回收率为0%.獐子岛至旅顺渤海侧沿海设置10~12个站位,调查浮游幼虫数量及个体大小分布,结果表明,进入5月中旬以后随着时间的推移,幼虫向旅顺渤海侧沿海集中,而且大个体幼虫的比例增加.调查结果证明,旅顺渤海侧沿海虾夷扇贝幼虫主要来自黄海北部增养殖海区.【总页数】4页(P588-591)【作者】李文姬;薛真福;李华琳;张明;罗鸣;鲁学辉;臧有才;刘项峰【作者单位】辽宁省海洋水产科学研究院,辽宁省应用海洋生物技术开放实验室,辽宁省水产分子生物学重点实验室,辽宁,大连,116023;辽宁省海洋水产科学研究院,辽宁省应用海洋生物技术开放实验室,辽宁省水产分子生物学重点实验室,辽宁,大连,116023;大连獐子岛渔业集团股份有限公司,辽宁,大连,116503;辽宁省海洋水产科学研究院,辽宁省应用海洋生物技术开放实验室,辽宁省水产分子生物学重点实验室,辽宁,大连,116023;辽宁省海洋水产科学研究院,辽宁省应用海洋生物技术开放实验室,辽宁省水产分子生物学重点实验室,辽宁,大连,116023;辽宁省海洋水产科学研究院,辽宁省应用海洋生物技术开放实验室,辽宁省水产分子生物学重点实验室,辽宁,大连,116023;大连獐子岛渔业集团股份有限公司,辽宁,大连,116503;大连獐子岛渔业集团股份有限公司,辽宁,大连,116503;辽宁省海洋水产科学研究院,辽宁省应用海洋生物技术开放实验室,辽宁省水产分子生物学重点实验室,辽宁,大连,116023【正文语种】中文【中图分类】S932.6【相关文献】1.大连沿海虾夷扇贝养殖群体遗传结构的研究 [J], 高悦勉;李国喜;赵银丽2.基于生态足迹模型沿海旅游资源开发规模评估--以大连旅顺口区为例 [J], 袁仲杰;高范;孙岳;钟慧颖3.旅顺口区虾夷扇贝海区采苗技术研究与现状调查 [J], 周志刚;蔡之昕;朱伟国;耿帅帅;潘欣茹4.大连沿海虾夷扇贝体内重金属含量研究与质量评价 [J], 庞艳华;姜伟;薛大方;黄大亮5.打造辽宁沿海经济带的重要增长极——来自大连旅顺经济技术开发区的调查 [J], 盛隆言因版权原因,仅展示原文概要,查看原文内容请购买。
虾夷扇贝养殖技术
柳中传;宋宗贤
【期刊名称】《海洋科学》
【年(卷),期】1991(000)004
【摘要】虾夷扇贝是海珍品之一,是日本、朝鲜的主要海水养殖品种,为目前世界扇贝科中经济价值最高、产量最大的种类。
虾夷扇贝与栉孔扇贝在生物学上有些不同,它不是终生附着生活, 一般当壳高达7~8mm后便失去足丝。
【总页数】4页(P11-14)
【作者】柳中传;宋宗贤
【作者单位】不详;不详
【正文语种】中文
【中图分类】S968.323
【相关文献】
1.海阳海域虾夷扇贝养殖技术研究 [J], 包家国;张晓霞;王卫民;孙鹏飞
2.海阳海域虾夷扇贝养殖技术研究 [J], 王卫民;包华君;纪玉娥;孙鹏飞
3.黄海北部虾夷扇贝吊耳养殖技术研究 [J], 张明;刘项峰;李华琳;付成东;李石磊;于佐安;腾炜鸣;乔英;李文姬
4.海阳海域虾夷扇贝养殖技术 [J], 王卫民;纪玉娥;刘红蕾;孙鹏飞
5.浅议日本虾夷扇贝的增养殖技术(上) [J], 侯雪菲;张光发;张晓芳;刘鹰
因版权原因,仅展示原文概要,查看原文内容请购买。
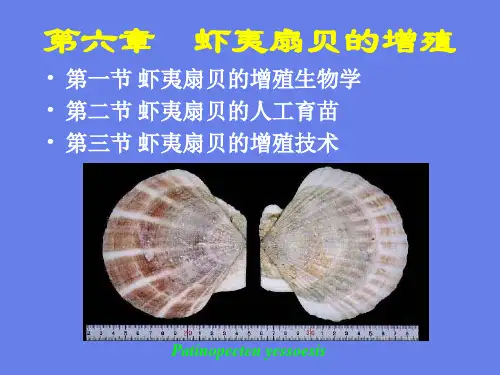
虾夷扇贝生活史
俗名:虾夷扇贝
名称由来:虾夷为中国古代对日本北海道地区的旧称,所以在那发现的扇贝就叫虾夷扇贝
生长过程:
育苗阶段
一、生殖细胞:由滤泡壁和生殖管的上皮细胞(生殖原细胞)分化
而成。
二、受精:虾夷扇贝为雌雄异体,体外受精。
卵不透明直径约为
55um,细胞质淡红色。
精子头部三角形,颈长5um,尾部自颈部后缘中央向后延伸,长约50-60um。
受精是指雌雄配子相遇,两者质膜相互融合,随之两个原核相互形成一个合子的过程,受精的结果产生了受精卵。
受精卵在受精后1小时产生第一极体,2小时产生第二极体,在第二极体产生半小时,形成极叶。
此时开始第一次卵裂。
三、卵裂:受精卵经过多次分裂,形成很多分裂球的过程,称为卵
裂。
受精后3小时2细胞、4小时40分4细胞、6小时10分8细胞、13小时40分32细胞。
四、囊胚与原肠胚:当桑葚胚的中央出现空腔,胚体呈现囊状,表
面遍生短纤毛时,称为囊胚。
受精后16小时进入囊胚期。
囊胚继续发育,26小时后形成原肠胚。
五、担轮幼虫:受精后34小时,原肠胚渐渐拉长呈倒梨形,原口
前移到胚体腹面,在原口的前端形成一轮口前纤毛环,纤毛环的中央长出一束顶纤毛束,中间为1-2根较长的主鞭毛。
在胚体的后端有一束较小的端纤毛。
原口相对应的背侧,外胚层细胞转化为壳腺能分泌几丁质的胚壳。
此时的胚胎称为膜内担轮幼虫。
担轮幼虫在受精膜内快速转动,最后破膜而出,成为膜后担轮幼虫(但产生上不把此阶段称做孵化,而是待发育到D形幼虫时才计算孵化率),此时的担轮幼虫为浮游生活,一般在直线方向上做旋转前进,并具有趋光性。
担轮幼虫不摄食,以卵黄为营养。
六、面盘幼虫:担轮幼虫继续发育,在12-15℃的水温条件下,受
精后63小时形成面盘幼虫,壳长102um*壳高78um。
其壳腺形成马靴形的胚壳,并从身体两侧逐渐下包,将口前纤毛环推向身体的两侧,形成面盘。
以面盘为运动和摄食器官的幼虫称面盘幼虫(又称D形幼虫)
七、壳顶幼虫:
1、壳顶初期:受精后第8天形成早期壳顶幼虫,壳长136um*壳
高115um。
原呈直线铰合的铰合部中央逐渐隆起,形成壳顶,面盘发达,消化盲囊包裹了大部分的胃,外套膜、鳃原基、足丝腺、和平衡囊等都已形成,但未很好分化。
足呈棒状,无伸缩能力。
2、壳顶中期:受精后14天,壳长大约可达155um、壳高131um,
进入壳顶中期,左右壳不对称。
3、壳顶后期:受精后21天,壳长215um*壳高191um,壳顶充分
隆起,消化盲囊覆盖了整个胃周围。
足呈靴状,能自由伸出壳外,
此时幼虫既能用面盘浮游,又能用足匍匐爬行。
鳃原基明显可见,上有鳃纤毛。
外套膜中央,投影在消化盲囊的腹缘有黑色的眼点,身体两侧各一个(又称眼点幼虫)。
4、匍匐幼虫:受精后25天,当壳长221um*壳高197um时足形成,
即将进入附着期。
八、稚贝期:受精后28天,壳长244um*壳高223um。
变态后形成
稚贝。
稚贝进入附着生活后,除壳顶部外,在其壳外部有极薄而透明的硬膜形成,此即所谓的周缘壳。
当壳长达到1mm时,壳如耳状。
壳长达3mm时才能见到一根根放射肋,壳长达6-10mm时,失去足丝附着生活结束。
九、幼贝期和成贝期:稚贝进一步发育,壳长达到0.5cm以上,形
态结构、生态习性与成贝基本一致时称为幼贝期;幼贝发育到第一次具有繁殖能力之后,直到老死之前,称为成贝期。
养殖阶段
底播增殖(放流增殖):遵循自然生长规律,为其选择砂泥底质渔场,清除敌害生物;播苗密度为8枚/平方米,亩播苗5500枚;每年的十一月份是虾夷扇贝的播种期,我们将幼贝直接撒播在20-40米深的海底,任其“野生野长”,大约两年左右壳高达到10厘米以上,开始拖网采捕或人工潜水采捕。
食物需求:扇贝为滤食性动物,对食物的大小有选择能力,但对种类无选择能力。
大小合适的食物随纤毛的摆动送入口中,不合适的颗粒由足的腹沟排出体外。
其摄食量与滤水速度有关,滤水速度在夜间1-3点为最低值。
因此摄食量在夜间最大。
主要食物为有机碎屑、悬浮在海水中的微型颗粒和浮游生物,如硅藻类、双鞭毛藻类、桡足类等;其次还有藻类的孢子、细菌等。
其食物种类组成与环境中的种类相一致。