抗体重排rearrangement
- 格式:pptx
- 大小:5.37 MB
- 文档页数:71


自然免疫:先天性免疫或固有性免疫,是个体出生是就具有的天然免疫,可通过遗传获得,是机体在长期进化过程中逐渐建立起来的主要针对入侵病原体的天然防御功能。
获得免疫:适应性免疫,是个体出生后,接触到生活环境中的多种异物抗原,并在不断刺激中逐渐建立起来的后天免疫,也称获得性免疫。
体液免疫:是指由B细胞介导的免疫应答。
B细胞在抗原刺激下分化增殖为浆细胞,合成分泌抗体,形成特异性体液免疫应答。
具有中和毒素、激活补体、调理吞噬等作用。
细胞免疫:是指由T细胞介导的免疫应答。
Td抗原刺激T细胞,T 细胞增殖分化为致敏T 细胞,通过Tc细胞的细胞毒作用以及Th1细胞释放淋巴因子的作用,从而表现出抗细胞内寄生的微生物、抗肿瘤、抗寄生虫和迟发型变态反应。
克隆选择:克隆选择学说,或称无性繁殖系选择学说,克隆选择学说的核心论点是:①带有各种受体的免疫活性细胞克隆早已存在,抗原的作用只是选择并激活相应的克隆;②细胞受体和该细胞后代所分泌的产物(抗体)具有相同的特异性。
抗原:抗原是一类能够诱导机体免疫应答并能与相应抗体或T细胞受体发生特异反应的物质。
半抗原与载体:能与相应的抗体结合而具有免疫反应性,但不能诱导免疫应答的物质。
载体:赋予半抗原以免疫原性的蛋白质。
半抗原+ 蛋白质→完全抗原三.抗原表位:抗原决定簇,又称表位,是抗原物质中能与其相应抗体或致敏淋巴细胞发生特异性结合的结构(免疫应答的特异性基础)。
超抗原:一类可直接结合抗原受体,激活大量(2%—20%)T细胞或B细胞克隆,并诱导强烈免疫应答的物质,主要包括细菌和病毒的成分及其产物等。
五.抗原组学:是建立在基因组学和蛋白组学基础上的新兴领域,它正在成长为继基因组学和蛋白组学后的学科重点。
抗原组学:抗原组学是建立在基因组学和蛋白质组学基础上的新兴领域。
利用抗原组学可以鉴定疫苗的候选抗原。
Ig:即免疫球蛋白,指具有抗体活性或化学结构与抗体相似的球蛋白。
Fab:免疫球蛋白(Ig)的抗原结合片断(Fab)。
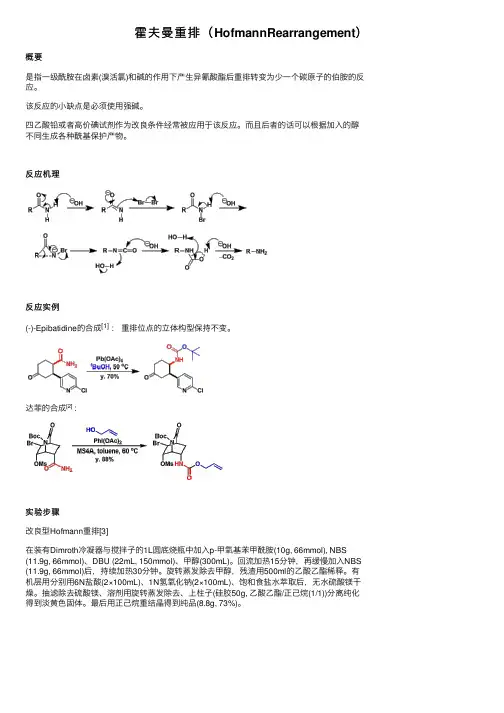
霍夫曼重排(HofmannRearrangement)概要
是指⼀级酰胺在卤素(溴活氯)和碱的作⽤下产⽣异氰酸酯后重排转变为少⼀个碳原⼦的伯胺的反应。
该反应的⼩缺点是必须使⽤强碱。
四⼄酸铅或者⾼价碘试剂作为改良条件经常被应⽤于该反应。
⽽且后者的话可以根据加⼊的醇不同⽣成各种酰基保护产物。
反应机理
反应实例
(-)-Epibatidine的合成[1] :重排位点的⽴体构型保持不变。
达菲的合成[2] :
实验步骤
改良型Hofmann重排[3]
在装有Dimroth冷凝器与搅拌⼦的1L圆底烧瓶中加⼊p-甲氧基苯甲酰胺(10g, 66mmol), NBS (11.9g, 66mmol)、DBU (22mL, 150mmol)、甲醇(300mL)。
回流加热15分钟,再缓慢加⼊NBS (11.9g, 66mmol)后,持续加热30分钟。
旋转蒸发除去甲醇,残渣⽤500ml的⼄酸⼄酯稀释。
有机层⽤分别⽤6N盐酸(2×100mL)、1N氢氧化钠(2×100mL)、饱和⾷盐⽔萃取后,⽆⽔硫酸镁⼲燥。
抽滤除去硫酸镁、溶剂⽤旋转蒸发除去、上柱⼦(硅胶50g, ⼄酸⼄酯/正⼰烷(1/1))分离纯化得到淡黄⾊固体。
最后⽤正⼰烷重结晶得到纯品(8.8g, 73%)。

第五章 免疫分子第一节 免疫球蛋白( Immunoglobulin, Ig )免疫球蛋白(immunoglobulin,Ig)包括成熟B 细胞的膜表面Ig(SmIg)和某个B 细胞克隆受抗原刺激、激活、分化、转化为浆细胞后分泌的Ig 。
由于Ig 具有与相应抗原发生特异结合的功能,故称为抗体(antibody,Ab)。
在人体内约有1×107个B 细胞克隆,各个B 细胞克隆的SmIg 分子的VL 、VH 结构都不一样,能识别差异极其微小的各种抗原决定簇,产生相应的Ig 分子。
分泌于血清中的Ig 分子结构极不均一,与抗原结合的特异性千差万别,Ig 自身表现的抗原性(血清型)也多种多样。
关于Ig 产生的遗传控制及多样性的起源,遗传学家们一直在进行研究,并存在分歧,提出的学说主要有胚系学说、体细胞突变学说和基因重排学说。
随着分子生物学技术的发展,基因重排学说不断被实验证实和完善,受到普遍赞同。
一、免疫球蛋白分子的基本结构Ig 分子的基本结构为,两条相同的轻链(light chain, L 链)在两条相同的重链(heavy chain, H 链)外侧,L 链和H 链之间,H 链与H 链之间,由二硫键连接成一个“Y ”字型。
每条L 链由210~230氨基酸组成,分子量约23ku ,每条H 链由420~446个氨基酸组成,分子量50~70ku 。
所有Ig 的L 链和H 链从氨基端起的100多个氨基酸的组成和顺序多变,称为可变区(variable region, V 区),L 链的V 区(VL)由108个氨基酸组成,H 链的V 区(VH)由107~130个氨基酸组成,VL 和VH 共同组成与相应抗原决定簇结合的部位。
在VL 和VH 中氨基酸组成的变化仅集中3个区域(占氨基酸总量的15%~20%),称为超变区(hypervariable region, HV)。
超变区是Ig 与抗原结定簇结合的互补决定区(complementarity-determining region,CDR)。




基因重排名词解释
基因重排(Gene Rearrangement)是指在某些生物体的基因组中,由于基因片段的移动、插入、删除或倒位等现象,导致基因在DNA分子上的排列顺序发生改变。
基因重排是基因组进化中的一个重要机制,可以增加基因组的多样性和复杂度,同时也可能对生物体的适应性和健康产生影响。
下面是一些与基因重排相关的名词解释:
1. 基因片段(Gene Segment):指编码蛋白质的DNA序列中的一段区域,通常具有特定的功能和序列特征。
2. 重组酶(Recombinase):一类催化基因重排的酶,能够切割DNA链并促使基因片段的互相重组。
3. V(D)J重排(V(D)J Rearrangement):在脊椎动物免疫系统中,负责生成抗体和T细胞受体的基因由不同的V、D和J基因片段
组成,通过V(D)J重排来产生不同的抗原识别结构,以应对不同的病原体。
4. 染色体易位(Chromosome Translocation):指两个染色体之间的基因重排,通常发生在染色体的两端或中间,可能影响基因的表达和功能。
5. 基因分化(Gene Divergence):由于基因重排等机制,在不
同物种或个体之间,基因片段序列和组合会发生变化,导致基因的结构和功能差异增加,称为基因分化。

抗体基因重排原理咱先得知道啥是抗体。
抗体就像是咱们身体里的小卫士,专门去识别那些外来的坏家伙,像细菌啊、病毒啊之类的。
那这些抗体是怎么来的呢?这就和基因重排有大大的关系喽。
在咱们的细胞里啊,抗体基因可不是一开始就整整齐齐地排好队准备工作的。
它们就像一堆乱七八糟的小积木块。
对于抗体来说,有好几个不同的基因片段,就比如说有V(可变区)片段、D(多样性区,这个不是每个抗体都有的哦)片段还有J(连接区)片段。
这些片段就散落在基因的各个角落。
当一个B淋巴细胞(这可是产生抗体的小工厂细胞呢)开始准备制造抗体的时候,就像是一场超级大抽奖。
这个细胞会从众多的V片段里挑出一个,然后呢,如果有D片段的话,再从D片段里选一个,最后从J片段里挑一个。
这一挑啊,那可就是随机的,就像你从好多不同颜色的糖果里乱抓一把一样。
选好这些片段之后呢,它们就要凑在一起啦。
这就像是把不同的小零件组装成一个超级武器。
细胞里有专门的酶来把这些片段连接起来,这个过程就像是小工匠在精心打造一件独一无二的宝贝。
一旦这些片段连接好了,就形成了抗体的可变区。
这个可变区可不得了,它就像是一把专门为识别特定敌人打造的钥匙。
为啥要这么复杂地重排基因来制造抗体呢?这就是大自然的智慧啦。
因为外界的细菌和病毒那可是千奇百怪的,如果抗体都是固定不变的,那肯定应付不过来呀。
通过这种基因重排,就可以产生数不清的不同抗体。
就好比你有了无数把不同的钥匙,总能找到一把去打开那些入侵坏家伙的锁。
而且哦,这个基因重排还不是一次性的。
在B淋巴细胞的发育过程中,还可能会发生一些小变化,让抗体变得更加多样化。
这就像是给小卫士们不断升级装备一样。
你看,咱们身体里的这个小世界多神奇呀。
这些微观的小基因,就像一群小精灵,通过这种奇妙的重排方式,为咱们的健康打造出强大的防御体系。
每次想到这个,我都觉得大自然真的是一个超级有创意的大艺术家呢。
它用这种看似有点“调皮”的方式,让我们的身体有了应对各种危险的能力。


多克隆抗体是由B淋巴细胞分泌的一种抗体分子,它们能够识别并结合多种抗原。
多克隆抗体的制备通常需要使用小鼠等动物进行免疫,然后从免疫动物的脾脏等组织中提取出B淋巴细胞,再通过一系列的细胞培养和筛选步骤来获得多克隆抗体。
基因重排是指在B淋巴细胞中发生的一种基因重组过程。
在B淋巴细胞的发育过程中,它们会经历一系列的基因重排,从而产生出具有不同抗原识别能力的抗体分子。
基因重排是通过重排基因的染色体位置来实现的,这一过程需要一系列的基因重组酶的参与。
在多克隆抗体的生产过程中,基因重排是非常重要的一个步骤,因为它能够产生出具有不同可变区的抗体分子,从而增加多克隆抗体的抗原识别能力和多样性。
RAG小鼠,全称为重组激活基因(Recombination-Activation Gene)小鼠,是一类通过基因工程方法构建的小鼠品系。
它们的特点是能够在体内或体外有效地重组抗体基因,从而产生特定类型的抗体。
RAG小鼠通常用于免疫学和分子生物学领域的研究,特别是在研究抗体基因的重组和表达、免疫系统发育和功能等方面具有重要价值。
RAG小鼠的品系特征主要体现在以下几个方面:
1. 抗体基因重排能力:RAG小鼠具有重组激活基因,能够促进抗体基因的重排,产生多样化的抗体。
这使得它们在研究抗体结构和功能、以及免疫应答机制方面具有优势。
2. 免疫缺陷:某些RAG小鼠品系可能因为基因改造而具有免疫缺陷,这使得它们在研究免疫系统的调控和病原体感染机制时更为便利。
3. 遗传背景:RAG小鼠的遗传背景通常是C57BL/6或BALB/c,这两种品系因其稳定的遗传背景和良好的繁殖性能,被广泛用于实验研究。
4. 基因敲除能力:通过Cre-loxP或其他基因敲除技术,RAG小鼠可以用于创建特定基因被敲除的模型,这对于研究基因功能和疾病机理非常有用。
5. 自发突变率:由于RAG小鼠的基因改造性质,它们的自发突变率可能较高,这为研究基因变异和突变效应提供了资源。
6. 实验应用:RAG小鼠在科研中的应用范围广泛,包括但不限于抗体工程、基因敲除、免疫学研究、肿瘤学研究等领域。
在使用RAG小鼠进行实验研究时,研究人员需要考虑到这些品系的特定特征,以及可能对实验结果产生的影响。
此外,由于RAG小鼠的基因改造性质,研究人员在实验设计和数据分析时需要特别注意基因敲除或插入可能带来的非特异性影响。