第三章 纤维素纤维的结构和性能
- 格式:doc
- 大小:146.50 KB
- 文档页数:17

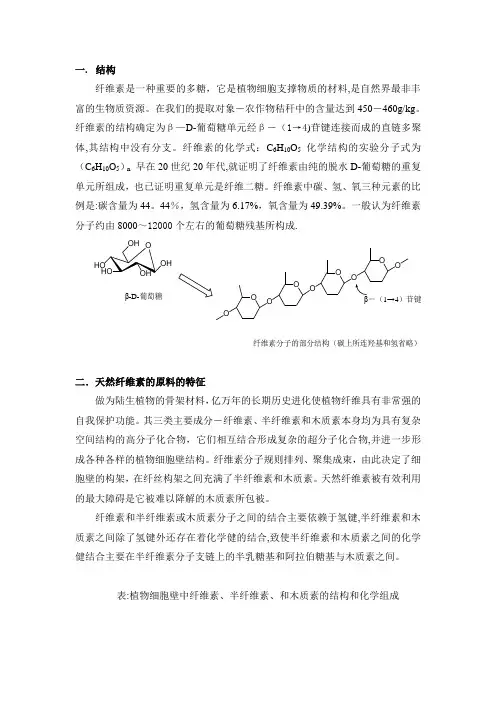
一.结构纤维素是一种重要的多糖,它是植物细胞支撑物质的材料,是自然界最非丰富的生物质资源。
在我们的提取对象-农作物秸秆中的含量达到450-460g/kg。
纤维素的结构确定为β—D-葡萄糖单元经β-(1→4)苷键连接而成的直链多聚体,其结构中没有分支。
纤维素的化学式:C6H10O5化学结构的实验分子式为(C6H10O5)n早在20世纪20年代,就证明了纤维素由纯的脱水D-葡萄糖的重复单元所组成,也已证明重复单元是纤维二糖。
纤维素中碳、氢、氧三种元素的比例是:碳含量为44。
44%,氢含量为6.17%,氧含量为49.39%。
一般认为纤维素分子约由8000~12000个左右的葡萄糖残基所构成.O OOOOOOOO1→4)苷键β-D-葡萄糖纤维素分子的部分结构(碳上所连羟基和氢省略)二.天然纤维素的原料的特征做为陆生植物的骨架材料,亿万年的长期历史进化使植物纤维具有非常强的自我保护功能。
其三类主要成分-纤维素、半纤维素和木质素本身均为具有复杂空间结构的高分子化合物,它们相互结合形成复杂的超分子化合物,并进一步形成各种各样的植物细胞壁结构。
纤维素分子规则排列、聚集成束,由此决定了细胞壁的构架,在纤丝构架之间充满了半纤维素和木质素。
天然纤维素被有效利用的最大障碍是它被难以降解的木质素所包被。
纤维素和半纤维素或木质素分子之间的结合主要依赖于氢键,半纤维素和木质素之间除了氢键外还存在着化学健的结合,致使半纤维素和木质素之间的化学健结合主要在半纤维素分子支链上的半乳糖基和阿拉伯糖基与木质素之间。
表:植物细胞壁中纤维素、半纤维素、和木质素的结构和化学组成项目纤维素木质素半纤维素结构单元吡喃型D-葡萄糖基G、S、H D-木糖、苷露糖、L-阿拉伯糖、半乳糖、葡萄糖醛酸结构单元间连接键β—1,4—糖苷键多种醚键和C—C键,主要是β-O-4型醚键主链大多为β-1,4-糖苷键、支链为β-1,2-糖苷键、β-1,3—糖苷键、β-1,6-糖苷键聚合度几百到几万4000 200以下聚合物β—1,4-葡聚糖G木质素、GS木质素、GSH木质素木聚糖类、半乳糖葡萄糖苷露聚糖、葡萄糖甘露聚糖结构由结晶区和无定型区两相组成立体线性分子α不定型的、非均一的、非线性的三维立体聚合物有少量结晶区的空间结构不均一的分子,大多为无定型三类成分之间的连接氢键与半纤维素之间有化学健作用与木质素之间有化学健作用天然纤维素原料除上述三大类组分外,尚含有少量的果胶、含氮化合物和无机物成分。

第三章纤维素纤维的结构和性能
纤维素是一种在自然界广泛存在的生物高分子物质,具有多种性能和应用。
它是由一系列葡萄糖单位通过β-1,4-键连接而成的线性聚合物。
纤维素在植物细胞壁中扮演着重要的结构支撑和生物功能的角色。
本章将介绍纤维素纤维的结构和性能。
纤维素纤维的结构主要有两种,一种是微纤结构,即纤维素分子在单个纤维中的排列方式;另一种是纤维结构,即由多个纤维素分子相互聚集而成的整体结构。
纤维素纤维的微纤结构可以分为两种:晶体区和无定型区。
晶体区主要由纤维素链的有序排列构成,具有较高的结晶度和强度。
无定型区则为纤维素链的非有序排列部分,结晶度和强度较低。
纤维素纤维的微纤结构对其性能有重要影响,晶体区的存在可以增加纤维的强度和刚度。
纤维素纤维的纤维结构主要有两种:平行排列和螺旋排列。
平行排列的纤维结构是指纤维素纤维中的纤维素链平行排列,形成平行纤维束。
螺旋排列的纤维结构则是指纤维素纤维中的纤维素链呈螺旋状排列。
纤维结构对纤维素纤维的柔软性和延展性具有重要影响。
纤维素纤维的性能与纤维结构和微纤结构密切相关。
首先,纤维结构决定了纤维的柔软性和延展性。
平行排列的纤维结构较硬而缺乏延展性,螺旋排列的纤维结构则具有良好的柔软性和延展性。
其次,微纤结构决定了纤维的强度和刚度。
晶体区的存在可以增加纤维的强度和刚度,而无定型区的存在则会降低纤维的性能。


第三章纤维素一、一次结构1.近程结构:链结构单元的组成和链接方式2.远程结构:分子的大小、形态、链的柔顺性和构象3.构象:构型一定的分子,在其键允许的限度内各基团绕单键内旋转形成聚合物的不同形态(分子热运动)4.构型:分子中的原子和基团由化学键所固定的空间几何排列。
(化学键断裂)5.纤维素大分子化学结构特点:⏹基本结构单元是D-吡喃式葡萄糖⏹纤维素大分子葡萄糖基都是β-苷键链接⏹纤维素大分子每个基环具有三个醇羟基-(2、3仲醇羟基,6伯醇羟基)⏹纤维素大分子的两个末端基性质不同(C1位还原性醛基,C4位隐形醛基非还原性),不同分子链具有极性和方向性6.纤维素链构象:⏹葡萄糖环的构象:4C1椅式构象(会画)⏹纤维素大分子链的构象:葡萄糖单元成椅式构象,每个单元上C2位羟基,C3位羟基和位取代基均处于水平位置C6⏹C5位羟甲基构象:tg构象⏹配糖角:β-1-4苷键的键角扭转角:葡萄糖苷键绕C1-O键形成夹角键形成夹角:葡萄糖苷键绕O-C4纤维素分子模型:伸直链模型弯曲链模型二、二次结构1.聚集态结构(超分子结构):处于平衡态时纤维素大分子链相互间几何排列特征2.聚集态结构研究:结晶结构(晶区和非晶区、晶胞大小及形式、分子链在晶胞内的堆砌形式)、取向结构(分子链和微晶取向)、原纤结构3.纤维素晶体:c键直立,a键前后(氢键),b轴位于左右方向4.辨认不同晶胞结构:X射线衍射、红外光谱、正交极化/幻角旋转13C核磁共振谱5.纤维素ⅠMeyer-Misch模型的特点:⏹纤维素分子链占据晶胞的4个角和中轴⏹四角上的链为4个相邻晶胞所共有,每个晶胞只含有两个分子链⏹晶胞中间链的走向和角上链的走向相反——反平行链排列;在轴向高度彼此半个葡萄糖基⏹b轴长度正好为纤维素二塘的长度。
分子链葡萄糖基团绕纵轴扭转180°(纤维二糖为基本结构单元)6.纤维素ⅡBlackwell 模型特点⏹纤维素分子链占据晶胞的4个角和中轴⏹晶胞中间链的走向和角上链的走向相同——同向平行链;在轴向高度彼此半个葡萄糖基⏹分子链平行于ac 面,-CH 2OH 均为-tg 构象;(1,4苷键键角为114.8°)⏹a 轴方向分子间氢键(020面,O(3)-H...O(6’);分子内氢键O(3)-H...O(5’)、O(2’)-H...O(6),晶胞对角线无氢键)7.纤维素Ⅱ结晶结构特点⏹存在两条空间群为P21的分子链,具有二次螺旋对称,角上链和中心链为反向平行链⏹中心链—-CH 2OH 具有-tg 构象,角上链-CH 2OH 具有-gt 构象⏹中心链和角上链在高度上相差半个葡萄糖基⏹分子链投影与ac 面有偏角(30°),与110面方向一致在020面【O(3)-H...O(5’)、O(2’)-H...O(6)】和110面O(2)-H...O(2’)内形成氢键。




教学课题:纤维素纤维的结构与性质教学目的:1、了解纤维及纺织纤维的概念及纤维的分类;2、理解麻纤维与棉纤维的结晶构造对纤维材料机械性能影响;3、掌握纤维素的化学性质教学重点:纤维及纺织纤维的概念/纤维素的化学性质教学难点:纤维素纤维的结构教学内容一、纤维概念纤维:直径一般为几微米,而长度比直径大百倍、千倍以上的细长物质称做纤维。
纺织纤维:可以用业制造纺织制品(纱、线、绳等)的纤维。
聚合度:小分子通过聚合反应生成的聚合物中小分子的个数。
结晶度:表示大分子结晶区含量大小的指标。
二、纺织纤维必须具备的性质1、牢度:纺织纤维必须具有相当之抗张强度,强度之最低限度可定为20千克/平方毫米或1.5克/旦尼尔(Denier)2、长度:纤维长度,愈长愈佳。
如长度在五毫米以下,则不易纺纱。
3、粗细:纤维愈细,纺成之纱线及织成之织物愈精致。
但若过细时,则强度随之变弱。
一般人造纤维之直径约在0.01至0.04毫米之间。
4、延展性及弹性:供制衣用者可弯曲不致折断,且能恢复原状等特性。
这些性质,皆与弹性及延展性直接有关,故纺织纤维必须具有适当之弹性及延展性。
5、胶著性:纺织纤维须具备能互相缠绕或胶著以成坚固纱线之特性,此性质大半因纤维之形状而定。
如棉纤维纤维扭曲,毛织维表面具有鳞片组织,因而易于互相缠绕而胶著。
这种性质于制造人造短纤维时,尤其重要。
6、保温性:衣服主要功能之一,为具有防寒御暑之性能,故纺织纤维必为热之不良导体。
7、耐久性:纺织纤维于织造、加工及使用时,均须承受相当程度力之作用,如无耐久之力,实无法使用。
8、不溶性:纺织纤维不但须有不溶于水的特性,即使对于肥皂水、弱碱、弱酸水等亦须不溶,因无论在织物之制成、加工过程中或制后使用时,皂水及弱碱、弱酸液,均为无法避免接触之物。
9、手感:手感柔软、乾、不粗硬,第一节纤维素纤维的结构与性质一、纤维素1、纤维素(cellulose):由葡萄糖组成的大分子多糖。
不溶于水及一般有机溶剂。


第三章纤维素纤维的结构和性能天然纤维素纤维(棉、麻)纤维素纤维再生纤维素纤维(粘胶纤维、铜氨纤维、醋酯纤维)§3.1纤维素纤维的形态结构一棉纤维的形态结构棉纤维是种子纤维,其主要成分为纤维素、果胶、蜡质、灰分、含氮物质。
外形:上端尖而封闭,下端粗而敞口,细长的扁平带子状,有螺旋状扭曲,截面呈腰子形,中间干瘪空腔。
最外层:初生胞壁从外到里分三层:中间:次生胞壁内部:胞腔1 初生胞壁决定棉纤维的表面性质,它又分为三层,最外层为果胶物质和蜡质所组成的皮层。
因而具有拒水性,在棉生长过程中起保护作用。
但在染整加工中不利。
2 次生胞壁纤维素沉积最后的一层,是构成纤维的主体部分,纤维素含量很高,其组成和结构决定棉纤维的主要性能。
3 胞腔输送养料和水分的通道,蛋白质、色素等物质的残渣沉积胞壁上,胞腔是棉纤维内最大的空隙,是染色和化学处理时重要的通道。
二麻纤维的形态结构麻纤维主要有:苎麻、亚麻是属于韧皮纤维,以纤维束形式存在单根纤维是一个厚壁、两端封闭、内有狭窄胞壁的长细胞苎麻两端呈锤头形或分支亚麻两端稍细呈纺锤形纵向有竖纹和横节主要化学组成和棉纤维一样是纤维素,但含量低。
§3.2纤维素大分子的分子结构纤维素是一种多糖物质,其大分子是由很多葡萄糖剩基连接而成,分子式为(C6H10O5)n复杂的同系物混合物,n为聚合度,棉聚合度为2500~ 10000,麻聚合度为10000~ 15000,粘胶纤维聚合度为250~ 500纤维素大分子的化学结构是由β-d-葡萄糖剩基彼此以1,4-甙键连接而成,结构如下每隔两环有周期性重复,两环为一个基本链节,链节数为(n-2)/2,n为葡萄糖剩基数,即纤维的聚合度,葡糖糖剩基上有三个自由存在的羟基,其中2,3位上是仲羟基,6位上伯羟基§3.3棉纤维的超分子结构超分子结构也称为微结构,主要指棉纤维中次生胞壁纤维素大分子的聚集态结构,纤维素大分子的排列状态,排列方向,聚集紧密程度等。
一X射线研究1棉纤维的X射线研究结果超分子结构中有晶体存在,有一定的取向度2棉纤维中纤维素的单元晶格单元晶格属于单斜晶系3 纤维的结晶度与取向度棉纤维的结晶度约为70%,麻纤维为90%,无张力丝光棉为50%,粘胶纤维为40%二电子显微镜的研究1 棉纤维的电镜图棉纤维中存在粗大的原纤,但原纤又是由更小的微原纤组成2 边缘(缨状)原纤模型及理论(见P43的图3-8)纤维素大分子通过整齐排列组成微原纤,又由微原纤进行整齐排列形成原纤,原纤中少数大分子的分支与其他大分子分支合并组成另外的连续网状组织。
§3.4 纤维素纤维的主要物理—机械性能纤维的拉伸性能与织物耐用性及服用性能之间有着密切的联系,而纤维的断裂强度、断裂伸长率、应力应变曲线、弹性均与纤维拉伸性能有关。
一纤维素纤维的断裂强度、断裂伸长率断裂强度:纤维在拉伸至断裂时所能承受的最大外力。
表示方法有:1 抗张强度(σ)又称断裂应力,极限强度,是指纤维或纱线单位截面上能承受的最大拉力。
2 断裂强度(相对强度)指单位线密度或纱线所能承受的最大应力,单位为N/tex,又有干强和湿强之分。
3 断裂长度(L R)指将纤维一端固定悬挂,由于纤维本身质量而发生断裂时的长度,单位km4 断裂伸长率断裂伸长与纤维原长之比一般在结晶度相同的情况下,取向度越低,断裂伸长率越高,韧性越大。
二纤维的初始模量初始模量也称为杨氏模量或弹性模量指材料所受应力与其相应形变之比。
纤维的初始模量是指纤维产生1%伸长所需的应力,以kg/mm2或 g/tex表示,反映纤维在外力作用下变形的难易程度。
三纤维的应力-应变曲线又称纤维的负荷-延伸曲线,是将纤维随着应力的增大逐渐发生应变的情况绘成的曲线 (具体见P47的图3-9)四纤维素纤维的断裂机理与纤维超分子结构的关系纤维在外力作用下发生断裂,是因为外力破坏了分子内共价键力或分子间作用力的结果。
断裂机理有两种解释:1 纤维大分子链在受外力作用时,由于不能承受外力的作用而发生大分子链的断裂,从而导致纤维材料的断裂。
2 纤维在受外力作用时,大分子间的作用力不足以抵抗外力的作用,使得大分子链间发生相对位移,甚至滑脱,从而导致纤维的断裂。
纤维素纤维的断裂机理不是由单纯的分子链断裂或分子链间的相对滑移造成,还可能是由于超分子中存在缺口、弱点,在受外力作用拉伸时,弱点出现应力集中,首先断裂,缺口逐渐增大,进而分子链断裂,导致纤维断裂。
棉、麻湿强大于干强的原因因为在潮湿状态下水的增塑作用,可以部分消除纤维中的弱点,使得大分子中的缺陷结构得到改善,使应力分布更加均匀,从而增大了纤维的强力。
粘胶湿强小于干强的原因因为粘胶纤维大分子的聚合度低,只有250-500左右,结晶度低,取向度也不高,本身分子间力小,其断裂主要是由于大分子链或其他结构单元之间相对滑移形成,而湿强低则是由于吸湿后水的溶胀作用,降低了纤维的分子间力,有利于分子链间或结构单元间的相对滑移。
五纤维的弹性弹性是指纤维从形变中回复原状的能力。
弹性高的纤维所组成的织物外观比较挺括,不易起皱,如毛织物及涤纶织物。
可复弹性形变(弹性形变)不可复形变(塑性形变)弹性大小表示方法:形变回复率和功回复率形变回复率=弹性形变/总形变功回复率=回复时的回缩功(回复功)/形变时的总功§3.5 纤维素纤维的主要化学性质一纤维的吸湿和溶胀吸湿性:纺织纤维在空气中吸收和放出水分的能力表示方法:吸湿率(回潮率)R 和含水率M吸湿率(回潮率):纤维内所含水分的质量与绝对干燥纤维的质量之比。
含水率:纤维内所含水分的质量与未经烘干纤维质量之比R=(W/D) × 100%M=W/(D+W) × 100%式中:W——试样吸收水分的质量 D——绝对干燥试样的质量影响纤维吸湿性的有关因素1 纤维上的亲水性基团,纤维大分子中,亲水性基团的多少和亲水性的强弱均能影响其吸湿能力的大小。
2 纤维的超分子结构,无定形区越大,吸湿能力越强,如粘胶大于棉。
3 纤维的比表面积(单位质量的纤维所具有的表面积),如细纤维大于粗纤维。
4 纤维内的空隙越多,水分子越容易进去,如粘胶纤维比棉纤维疏松,吸湿能力也就越强。
5 纤维中各种伴生物和杂质对吸湿也有影响。
6 与空气温度、湿度有关。
二碱对纤维素纤维的作用1 纤维素对碱的稳定性稀碱低温条件下,纤维素对碱还是比较稳定。
但高温、浓碱时纤维素降解迅速,在高温且有空气存在时,即使较稀的碱液,也会使纤维素氧化。
2 浓碱对纤维素的作用在常温下,浓NaOH溶液会使天然纤维素纤维、溶胀,纵向收缩,直径增大。
如果施加张力,可防止收缩,及时洗除碱液可达到丝光效果。
如不施加张力,则发生碱缩。
对于针织物,增加弹性和厚实的手感。
碱与纤维素作用后的产物叫碱纤维素,一种不稳定的化合物,水洗可回复原来结构,通常成为纤维素Ⅱ,但结晶度下降,无定形区增加,对染料的吸附能力和化学反应能力大大提高。
三纤维素与酸的作用酸对纤维素分子中的甙键水解起催化作用,导致纤维素大分子聚合度降低,而使纤维受到损伤。
1酸与纤维素的作用原理纤维素大分子的1,4甙键具有缩醛的性质,对碱稳定,对酸敏感,酸对甙键水解起催化作用,导致纤维素纤维聚合度的降低和潜在的醛基增加。
得到水解纤维素。
酸与纤维素的反应首先发生在纤维无定形部分和晶区表面。
2 影响纤维素酸性水解的因素(1)主要是酸的性质、浓度、水解反应的温度、作用时间。
但在适当条件下,还是有一定的稳定性。
(2)与纤维素的种类有关,纤维中无定形区越多,越易水解。
四氧化剂对纤维素的作用纤维素一般不受还原剂的影响,而氧化剂则能使纤维素氧化成为氧化纤维素,使纤维受到损伤。
1 纤维素的氧化主要发生在葡萄糖剩基的三个羟基和大分子末端的潜在醛基上剧烈氧化的最终产物为CO2和H2O,具体的氧化反应见P57-58。
2 氧化剂的种类选择性氧化剂:对纤维素的某一位置上的基团进行专一氧化。
如NaClO2非选择性氧化剂:能对纤维素不同位置上的基团进行氧化作用的氧化剂,如NaClO、H2O2、KMnO4等3 氧化纤维素的性质还原型氧化纤维素:指分子中含有大量醛基的氧化纤维素酸型氧化纤维素:分子中含有大量羧基的氧化纤维素。
潜在损伤:纤维素氧化后生成还原型氧化纤维素时,只是葡萄糖环发生破裂,并没有使纤维素大分子断裂,纤维的强度变化不大,但不稳定,经碱煮后,强力下降非常大,这种现象称为潜在损伤氧化纤维素与水解纤维素的性能比较:见P60 表3-7五光、热及微生物对纤维素的作用1 光对纤维素的作用光和大气使纤维素纤维发生氧化和裂解反应。
光解作用:在波长较短的紫外线照射下,直接引起C-C键 C-O键的断裂,与空气无关。
光敏作用:指在波长靠近紫外光及可见光区,同时有光敏剂、氧及水分的存在,使纤维发生光氧化。
2 热对纤维素的作用纤维素对热稳定性较好,但在高温时,纤维素的稳定性下降,发生明显热退化现象,伴随氧化及水解反应。
3 微生物对纤维素的作用细菌和霉菌均属微生物,在其分泌物酶的作用下,纤维素易发生水解,生成较简单的糖,而使其强度下降。
§3.6纤维素共生物及棉籽壳纤维素共生物主要有果胶物质、含氮物质、蜡质、灰分、色素等,而棉纤维则还有伴生物棉籽。
共生物在染整加工中影响纤维的吸水、染色、白度等性能。
一果胶物质存在位置:纤维的初生胞壁中。
主要成分:果胶酸的衍生物化学组成:多半乳糖醛酸,具有链状结构。
含量:成熟棉纤维小于0.9%~1%,不成熟棉纤维则高达6%虽然果胶酸含有大量的亲水性的羟基羧基,但在棉纤维中部分以钙盐、镁盐和甲酯的形式存在,所以亲水性比纤维素本身要低。
果胶物质对纤维的色泽和润湿性有一定的影响。
二含氮物质存在位置:主要以蛋白质的形式存在于纤维的胞腔中。
小部分存在于初生胞壁和次生胞壁中。
含量:0.2~0.4%含氮物质分为两部分:一部分为无机盐类,如硝酸盐或亚硝酸盐可溶于60℃热水、常温稀酸稀碱。
另一部分主要成分为蛋白质,在烧碱溶液中长时间煮沸才能除去若以蛋白质形式存在,则加工或服用过程中,与有效氯接触很容易形成氯胺,引起织物泛黄。
三蜡质不溶于水而能被有机溶剂提取的物质存在位置:纤维的初生胞壁中。
主要成分:脂肪族高级一元醇含量:0.5~06%对纤维的润湿性能影响很大,可借助皂化和乳化作用去除。
四灰分主要成分:无机盐含量:1~2%对纤维的吸水性、白度和手感有一定影响,其中某些盐类和氧化铁等对漂白剂的分解有催化作用,加速漂白剂对纤维的损伤。
可通过水洗和酸洗去除。
五色素有色物质,影响织物的白度,可通过漂白作用去除。
六棉籽壳籽棉在轧花过程中,少量的残片附在纤维上,影响织物的外观。
化学组成:木质素为主,还有单宁、纤维素、半纤维素及其他多糖。
在高温烧碱液的长时间作用下,棉籽壳发生溶胀,变的松软而解体通过水洗和受机械摩擦作用而脱落。