重组蛋白包涵体表达的复性和纯化
- 格式:ppt
- 大小:1.52 MB
- 文档页数:24

包涵体纯化蛋白复性的方法操作流程返回:包涵体复性常见问题分析与解决包涵体的纯化和复性总结包涵体折叠复性的方法应具备以下几个特点:较高的活性蛋白质回收率;复性产物易于与错误折叠蛋白质分离;折叠复性后能够得到较高浓度的蛋白;折叠复性方法易于放大等。
蛋白复性的过程通常分为以下几个步骤:包涵体的洗涤包涵体在溶解之前需要进行洗涤,包涵体中主要含有重组蛋白,但也有一些杂质,如一些外膜蛋白、质粒DNA等,这些杂质会与包涵体粘连在一起,可以通过洗涤去除大多数杂质,但无法将杂蛋白去除干净。
包涵体的洗涤通常选用浓度较低的变性剂,如2M尿素在50mM Tris,pH7。
0—8。
5,1mM EDTA中洗涤包涵体。
此外可以用温和去垢剂TritonX—100洗涤去除膜碎片和膜蛋白。
同时,因为去垢剂的洗涤能力会随溶液离子强度升高而加强,所以在洗涤包涵体时可加入低浓度的尿素或高浓度的NaCl,使包涵体的纯度达到50%以上。
包涵体的溶解变性剂如尿素、盐酸胍,主要是通过离子间的相互作用,打断包涵体蛋白分子间的各种化学键,使多肽延伸.盐酸胍是较尿素强的变性剂,它能溶解尿素不溶的包涵体.SDS、正十六烷基三甲基铵氯化物、Sarkosyl等也可以破坏蛋白内的疏水键,溶解一些包涵体蛋白质。
另外,从某些含有半胱氨酸的蛋白质中分离出的包涵体,通常含有一些链间形成的二硫键和链内的非活性二硫键,这就还需加入还原剂,如巯基乙醇、二硫基苏糖醇(DTT)、二硫赤藓糖醇、半胱氨酸等。
这种还原性试剂能够同半胱氨酸形成各种二硫化物中间体,也会被复性用的二硫化物置换试剂所取代。
二硫键的形成和断裂是可逆的,直到最有利的蛋白二硫键形成,这个平衡才会被破坏。
常用变性剂对比尿素较盐酸胍慢而弱,溶解度为用尿素溶解具有不电离,呈中性,成(8-10M)70-90%本低,蛋白质复性后除去不会造成大量蛋白质沉淀溶解的包涵体可选用多种色谱法纯化盐酸胍(6-8M)溶解能力达95%以上,且溶解作用快而不造成重组蛋白质的共价修饰成本高、在酸性条件下易产生沉淀、复性后除去可能造成大量蛋白质沉淀对蛋白质离子交换色谱有干扰包涵体的复性复性是指通过去除变性剂使目标蛋白从完全伸展的变性状态恢复到正常的折叠结构,同时去除还原剂确保二硫键正常形成。


表达重组包涵体蛋白的变性溶解及复性研究概述高应瑞(译)目前,由于基因组序列数据库的快速扩增,重组蛋白的规模化生产已经成为迫切需要。
因此,通过基因重组生产蛋白质,必须考虑其蛋白特性和活性构象,以确定它们的生物学功能。
如果蛋白质的功能以及其基因的生物学功能未知,无论该蛋白质是在天然结构或在非天然结构,都不能通过生物活性评估来对其进行评估。
至今为止,有越来越多的异源重组蛋白成功得到表达。
其中大肠埃希氏菌(大肠杆菌)是最常使用的表达系统。
然而, 在大肠杆菌中异源表达外源基因往往会导致表达蛋白形成不溶性包涵体(IBS)。
包涵体必须经过变性溶解及复性过程后才会恢复其蛋白的生物活性。
包涵体复性过程的是一个非常复杂的过程,往往需要经过大量的反复试验才能确定其复性工艺参数。
在无活性的包涵体到具有生物活性蛋白的过程中,有两个最重要的影响因素:即包涵体的变性与复性。
包涵体蛋白的溶解必须使其蛋白肽链单分子进行分散并使最大程度减少蛋白质分子肽链内以及肽链之间的相互作用。
溶剂的选择,如尿素,盐酸胍,或洗涤剂等,在包涵体溶解过程中起关键作用,蛋白质变性溶解之后进行复性。
包涵体蛋白溶解后,即可进行蛋白复性。
蛋白复性过程不是单一反应,而蛋白的正确折叠与错误折叠和蛋白聚集的竞争过程。
在蛋白复性的竞争过程中,蛋白的复性率主要取决于变性剂的最终浓度以及溶剂的条件。
本文主要讨论的重点是复性工艺中对变性剂去除方式、溶剂、小分子添加剂以及工艺条件。
小分子添加剂的作用来自其在体内的功能定位。
小分子添加剂能够在生物体缺水的情况下维持体内蛋白质的稳定性,因此被命名为渗透剂[1,2]。
在另一个实例中,基因突变,蛋白的错误折叠常常导致蛋白的失活,从而导致机体生长的异常或某些细胞功能的异常。
实验已证明在某些小分子添加剂的存在时,这些失活蛋白质能够恢复正常功能,而且细胞能够生长正常。
由于小分子能够帮助蛋白质正确折叠,因此他们也被称为化学伴侣。
许多小分子添加剂都是渗透剂和化学伴侣。

一、菌体的裂解1、怎样裂解细菌?细胞的破碎方法1.高速组织捣碎:将材料配成稀糊状液,放置于筒内约1/3体积,盖紧筒盖,将调速器先拨至最慢处,开动开关后,逐步加速至所需速度。
此法适用于动物内脏组织、植物肉质种子等。
2.玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。
3.超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料,用大肠杆菌制备各种酶,常选用50-100毫克菌体/毫升浓度,在1KG至10KG 频率下处理10-15分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施,时间以及超声间歇时间、超声时间可以自己调整,超声完全了菌液应该变清亮,如果不放心可以在显微镜下观察。
对超声波及热敏感的蛋白和核酸应慎用。
4.反复冻融法:将细胞在-20度以下冰冻,室温融解,反复几次,由于细胞内冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。
5.化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好,我用的浓度一般为1mg/ml。
无论用哪一种方法破碎组织细胞,都会使细胞内蛋白质或核酸水解酶释放到溶液中,使大分子生物降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以抑制那些活性中心需要有疏基的蛋白水解酶的活性,加入苯甲磺酰氟化物(PMSF)也能清除蛋白水解酶活力,但不是全部,而且应该在破碎的同时多加几次;另外,还可通过选择pH、温度或离子强度等,使这些条件都要适合于目的物质的提取。
这是标准配方:裂解液:50mM Tris-HCl(pH8.5~9.0), 2mM EDTA, 100mM NaCl, 0.5% Triton X-100, 1mg /ml溶菌酶。

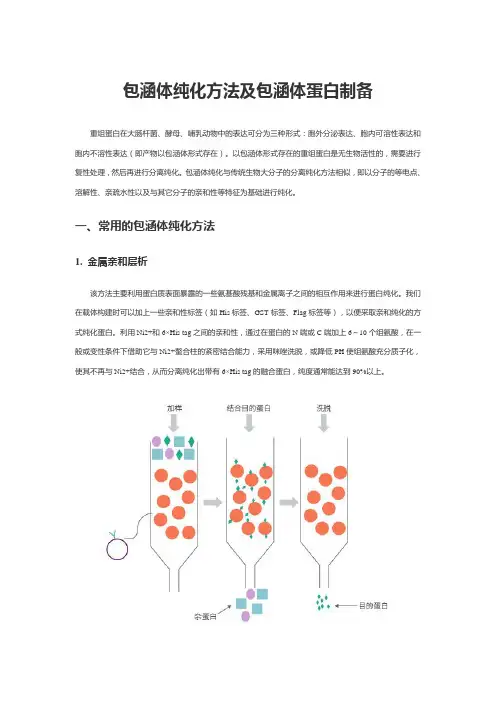
包涵体纯化方法及包涵体蛋白制备重组蛋白在大肠杆菌、酵母、哺乳动物中的表达可分为三种形式:胞外分泌表达、胞内可溶性表达和胞内不溶性表达(即产物以包涵体形式存在)。
以包涵体形式存在的重组蛋白是无生物活性的,需要进行复性处理,然后再进行分离纯化。
包涵体纯化与传统生物大分子的分离纯化方法相似,即以分子的等电点、溶解性、亲疏水性以及与其它分子的亲和性等特征为基础进行纯化。
一、常用的包涵体纯化方法1. 金属亲和层析该方法主要利用蛋白质表面暴露的一些氨基酸残基和金属离子之间的相互作用来进行蛋白纯化。
我们在载体构建时可以加上一些亲和性标签(如His标签、GST标签、Flag标签等),以便采取亲和纯化的方式纯化蛋白。
利用Ni2+和6×His tag之间的亲和性,通过在蛋白的N端或C端加上6~10个组氨酸,在一般或变性条件下借助它与Ni2+螯合柱的紧密结合能力,采用咪唑洗脱,或降低PH使组氨酸充分质子化,使其不再与Ni2+结合,从而分离纯化出带有6×His tag的融合蛋白,纯度通常能达到90%以上。
2.离子交换层析离子交换层析是根据在一定pH条件下,蛋白质所带电荷不同而进行的分离的方法。
常用的离子交换剂有羧甲基纤维素(阳离子交换剂,弱酸型)和二乙基氨基乙基纤维素(阴离子交换剂,弱碱型)。
阴离子交换基质结合带有负电荷的蛋白质,所以这类蛋白质被留在柱子上,需要通过提高洗脱液中的盐浓度等措施将其洗脱下来。
根据蛋白质结合能力不同,洗脱的速度会存在差异,通常结合较弱的蛋白质先被洗脱。
反之,阳离子交换基质会结合带有正电荷的蛋白质,可以通过提高洗脱液的PH或增加洗脱液中的盐浓度将蛋白洗脱下来。
3.凝胶过滤层析凝胶过滤层析通常又称为分子筛方法,主要是根据蛋白质的大小和形状达到分离和纯化的目的。
一般是大分子先流出来,小分子后流出来。
此方法的优点在于层析所用的凝胶属于惰性载体,不带电荷并且吸附力弱,可较广的温度范围内进行。

板块一、大肠杆菌(这里讨论的大肠杆菌为非分泌到培养基中的重组蛋白,是否有重组蛋白分泌到培养基中的工程菌我没有见过。
)一、关于菌体的量大肠杆菌表达的基因工程蛋白是纯化人员最方便获得的原料,对纯化工艺开发来说几乎没有原料方面的限制。
常看到有战友用个几毫升的菌液去做纯化,对此我十分不解,同样要做,为什么不多做点呢?很少的菌体会给纯化带来一些难以估计的问题,工艺的重复性和放大往往出现问题。
因此,要做个好工艺就多发酵表达一些菌体吧。
我做纯化时,初始工艺摸索用的菌体量一般为10g左右。
二、关于是否包涵体表达包涵体的定义我就不讲了。
我要讲的是,一个基因工程蛋白是否是包涵体表达的说法本身就不完全准确。
至于包涵体在电镜下的晶体颗粒表现等等对我们纯化来说毫无意义,我相信做纯化工艺的人没有谁去看这个电镜,也不关心。
我们判断的依据只是SDS-PAGE,目的蛋白在破菌沉淀中,我们就认为是包涵体表达,但这是一个似是而非的结论。
看着没问题,实际上是有毛病的。
关键在于你用的是什么破菌缓冲液!有些蛋白在用缓冲液A破菌时是在破菌沉淀中,而用缓冲液B破菌时却在破菌上清中。
缓冲液A和B的差别可能只是pH上相差1-2个单位。
那么它是包涵体表达还是可溶上清表达呢?说这个问题主要在于有些战友往往非常在意他的目标蛋白是否包涵体表达。
甚至还有包涵体表达就用专门的包涵体蛋白纯化方法等等。
我们应该关心的是目标蛋白在什么缓冲体系下是可溶的,在什么缓冲体系下是不溶的!不要让包涵体这个概念给你误导。
三、关于表达量我们常常在发表的文章上看到,我这个工程菌的目标蛋白的表达量达到菌体总蛋白的30%、50%等等。
我要说都是文章的作者在忽悠。
不知道他们是如何定量的,用的最多的大概就是SDS-PAGE的扫描分析吧。
且不说一个SDS-PAGE不能表现出所有的菌体蛋白,电泳的染色方法、染色脱色强度、照片的曝光强度、扫描分析时的条带选择等等无不对这个百分比影响巨大。
在公司的QC部门做的对此应该最有体验,20%的条带要它变成30%又有何难?我的观点是对待表达量的描述不可定量,只能定性。

包涵体纯化蛋白复性的方法操作流程返回:包涵体复性常见问题分析与解决包涵体的纯化与复性总结包涵体折叠复性的方法应具备以下几个特点:较高的活性蛋白质回收率;复性产物易于与错误折叠蛋白质分离;折叠复性后能够得到较高浓度的蛋白;折叠复性方法易于放大等。
蛋白复性的过程通常分为以下几个步骤:包涵体的洗涤包涵体在溶解之前需要进行洗涤,包涵体中主要含有重组蛋白,但也有一些杂质,如一些外膜蛋白、质粒DNA等,这些杂质会与包涵体粘连在一起,可以通过洗涤去除大多数杂质,但无法将杂蛋白去除干净。
包涵体的洗涤通常选用浓度较低的变性剂,如2M尿素在50mM Tris,pH7、0-8、5,1mM EDTA中洗涤包涵体。
此外可以用温与去垢剂TritonX-100洗涤去除膜碎片与膜蛋白。
同时,因为去垢剂的洗涤能力会随溶液离子强度升高而加强,所以在洗涤包涵体时可加入低浓度的尿素或高浓度的NaCl,使包涵体的纯度达到50%以上。
包涵体的溶解变性剂如尿素、盐酸胍,主要就是通过离子间的相互作用,打断包涵体蛋白分子间的各种化学键,使多肽延伸。
盐酸胍就是较尿素强的变性剂,它能溶解尿素不溶的包涵体。
SDS、正十六烷基三甲基铵氯化物、Sarkosyl等也可以破坏蛋白内的疏水键,溶解一些包涵体蛋白质。
另外,从某些含有半胱氨酸的蛋白质中分离出的包涵体,通常含有一些链间形成的二硫键与链内的非活性二硫键,这就还需加入还原剂,如巯基乙醇、二硫基苏糖醇(DTT)、二硫赤藓糖醇、半胱氨酸等。
这种还原性试剂能够同半胱氨酸形成各种二硫化物中间体,也会被复性用的二硫化物置换试剂所取代。
二硫键的形成与断裂就是可逆的,直到最有利的蛋白二硫键形成,这个平衡才会被破坏。
常用变性剂对比尿素较盐酸胍慢而弱,溶解度为用尿素溶解具有不电离,呈中性,成(8-10M)70-90%本低,蛋白质复性后除去不会造成大量蛋白质沉淀溶解的包涵体可选用多种色谱法纯化盐酸胍(6-8M)溶解能力达95%以上,且溶解作用快而不造成重组蛋白质的共价修饰成本高、在酸性条件下易产生沉淀、复性后除去可能造成大量蛋白质沉淀对蛋白质离子交换色谱有干扰包涵体的复性复性就是指通过去除变性剂使目标蛋白从完全伸展的变性状态恢复到正常的折叠结构,同时去除还原剂确保二硫键正常形成。

一、菌体的裂解1、怎样裂解细菌?细胞的破碎方法1.高速组织捣碎:将材料配成稀糊状液,放置于筒内约1/3体积,盖紧筒盖,将调速器先拨至最慢处,开动开关后,逐步加速至所需速度。
此法适用于动物内脏组织、植物肉质种子等。
2.玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。
3.超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料,用大肠杆菌制备各种酶,常选用50-100毫克菌体/毫升浓度,在1KG至10KG 频率下处理10-15分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施,时间以及超声间歇时间、超声时间可以自己调整,超声完全了菌液应该变清亮,如果不放心可以在显微镜下观察。
对超声波及热敏感的蛋白和核酸应慎用。
4.反复冻融法:将细胞在-20度以下冰冻,室温融解,反复几次,由于细胞内冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。
5.化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好,我用的浓度一般为1mg/ml。
无论用哪一种方法破碎组织细胞,都会使细胞内蛋白质或核酸水解酶释放到溶液中,使大分子生物降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以抑制那些活性中心需要有疏基的蛋白水解酶的活性,加入苯甲磺酰氟化物(PMSF)也能清除蛋白水解酶活力,但不是全部,而且应该在破碎的同时多加几次;另外,还可通过选择pH、温度或离子强度等,使这些条件都要适合于目的物质的提取。
这是标准配方:裂解液:50mM Tris-HCl(pH8.5~9.0), 2mM EDTA, 100mM NaCl, 0.5% Triton X-100, 1mg /ml溶菌酶。

包涵体蛋白的纯化及复性讨论专贴!zbbnet wrote:最近实验遇到了大麻烦,说出来大伙帮忙想想法。
实验所用菌株为BL21,载体pMAL-p2X,与MBP融合表达,目的蛋白有三对二硫键。
最初的设计思想是想通过MBP的信号肽实现可分泌表达,但实验发现主要是包涵体表达。
菌体超声破碎后12000rpm离心,所获得的沉淀不能很好的贴在离心管底,而是处于部分溶解状态,对沉淀再作如下处理:1、2M尿素洗涤,pH8.5,12000rpm离心,获得的上清不是澄清透明而是略显混浊,这次的沉淀能够牢固的贴在管底。
2、4M尿素洗涤,pH9.0,12000rpm离心,获得的上清不是澄清透明而是略显混浊,这次的沉淀为半溶状态的冻状物。
3、8M尿素溶解上述2中的沉淀,几乎不见不溶的颗粒,但溶液还是略显混浊。
上述三溶液取样电泳,均见较多的目的蛋白。
对3中的溶液取样调pH至12溶液依然混浊。
0.22um过滤,无法去除混浊物。
考虑到混浊物是多聚体,样品中加入1%SDS和0.2%β-Me,都无法改善。
做上述处理,主要考虑到混浊物是目的蛋白,因为目的蛋白设计了6HIS但不能成功的结合Ni柱。
主要想请教这样几个问题:1、有没有那位战友也碰上我上述的问题,这种问题是如何解决的?2、我所描述的包涵体的状态说明了什么问题?3、对与蛋白溶液混浊有什么好的处理方法?暂不讨论分泌表达的问题。
回答:1. 你的情况很正常,根据三次沉淀处理情况,应该可以肯定你的蛋白在8MUrea中是完全可以裂解的。
并且该蛋白如若作复性应该可溶性较佳。
但根据我个人的经验。
如果MBP融合蛋白还是以包含体表达的话是很没有意思的,建议作单独表达,除非单独表达不行。
否则即使完全复性,目的蛋白活性也大打折扣。
另可优化表达条件,有可能可容表达的,不要放弃,你的破菌上清也可以跑胶看看有没有目的蛋白。
2. 说明蛋白蔬水性不强。
3。
有些蛋白裂解后有浑浊非常正常,尤其是MBP融合的,即使做到可容也会有白乎乎浑浊的现象,只要不影响使用就行,没必要在意。
包涵体纯化蛋白复性得方法操作流程返回:包涵体复性常见问题分析与解决包涵体得纯化与复性总结包涵体折叠复性得方法应具备以下几个特点:较高得活性蛋白质回收率;复性产物易于与错误折叠蛋白质分离;折叠复性后能够得到较高浓度得蛋白;折叠复性方法易于放大等。
蛋白复性得过程通常分为以下几个步骤:包涵体得洗涤包涵体在溶解之前需要进行洗涤,包涵体中主要含有重组蛋白,但也有一些杂质,如一些外膜蛋白、质粒DNA等,这些杂质会与包涵体粘连在一起,可以通过洗涤去除大多数杂质,但无法将杂蛋白去除干净。
包涵体得洗涤通常选用浓度较低得变性剂,如2M尿素在50mM Tris,pH7。
0-8。
5,1mMEDTA中洗涤包涵体。
此外可以用温与去垢剂TritonX-100洗涤去除膜碎片与膜蛋白。
同时,因为去垢剂得洗涤能力会随溶液离子强度升高而加强,所以在洗涤包涵体时可加入低浓度得尿素或高浓度得NaCl,使包涵体得纯度达到50%以上。
包涵体得溶解变性剂如尿素、盐酸胍,主要就是通过离子间得相互作用,打断包涵体蛋白分子间得各种化学键,使多肽延伸。
盐酸胍就是较尿素强得变性剂,它能溶解尿素不溶得包涵体。
SDS、正十六烷基三甲基铵氯化物、Sarkosyl等也可以破坏蛋白内得疏水键,溶解一些包涵体蛋白质。
另外,从某些含有半胱氨酸得蛋白质中分离出得包涵体,通常含有一些链间形成得二硫键与链内得非活性二硫键,这就还需加入还原剂,如巯基乙醇、二硫基苏糖醇(DTT)、二硫赤藓糖醇、半胱氨酸等。
这种还原性试剂能够同半胱氨酸形成各种二硫化物中间体,也会被复性用得二硫化物置换试剂所取代。
二硫键得形成与断裂就是可逆得,直到最有利得蛋白二硫键形成,这个平衡才会被破坏。
常用变性剂对比尿素(8-1较盐酸胍慢而弱,溶解度为7用尿素溶解具有不电离,呈中性,成0M)0-90%本低,蛋白质复性后除去不会造成大量蛋白质沉淀溶解得包涵体可选用多种色谱法纯化盐酸胍(6-8M)溶解能力达95%以上,且溶解作用快而不造成重组蛋白质得共价修饰成本高、在酸性条件下易产生沉淀、复性后除去可能造成大量蛋白质沉淀ﻫ对蛋白质离子交换色谱有干扰包涵体得复性复性就是指通过去除变性剂使目标蛋白从完全伸展得变性状态恢复到正常得折叠结构,同时去除还原剂确保二硫键正常形成、一般在尿素浓度4M左右时复性过程开始,到2M左右时结束;盐酸胍可以从4M开始,到1、5M时结束。
IFN-α重组蛋白包涵体溶解和蛋白纯化复性一、表达产物处理1、表达菌液8000rpm 4℃离心10min2、菌体沉淀按10:1(菌重5g,加50ml)裂解缓冲液,冰上超声至清亮(250W,超声5s,间歇5s,功率35%)3、取样取100ul 超声菌液并离心,标记超声上清,超声沉淀4、12000g 4℃离心15min,上清备用,标记超声上清5、超声沉淀用含2M 尿素的裂解缓冲液,以20:1 比例重悬,继续超声5min6、12000g 4℃离心20min7、超声沉淀用含1% Triton X-100 的裂解缓冲液重悬,4℃放置10min8、12000g 4℃离心15min,获得包涵体9、获得包涵体用Binding Bufer 重悬,4℃放置过夜二、包涵体的纯化1、放置过夜包涵体4℃高速离心,收集上清备用2、取样取离心上清,标记柱前3、使用Ni-NTA 基质进行纯化,用Binding Buffer 平衡Ni 柱,柱子平衡后低流速上样,整个上样过程使样品处于冰上,上样后用3~5 个柱体积的Wash Buffer 进行漂洗,最后用Elution Buffer 洗脱4、取样取柱后,标记柱后三、包涵体复性1、透析袋处理方法:把透析袋剪成适当长度(10~20cm)小段,在大体积的2% (W/V)NaHCO 3 和1mM EDTA (PH8.0)中将透析袋煮沸10min。
用蒸馏水彻底清洗透析袋。
放在1mM EDTA(PH8.0)中将之煮沸10min。
冷却后存放于4℃,必须确保透析袋始终浸没在溶液内。
从此时起取用透析袋时必须戴手套保持清洁,用前在透析袋内装满水然后排出,将之清洗干净2、复性采用梯度透析法,纯化后包涵体用含4M 尿素的透析缓冲液稀释蛋白浓度至约100ug/ml,从4M、2M、2M、0.5M 依次降低透析液中尿素浓度,每个浓度均4℃透析,4~6h 换新鲜透析液一次,高速离心去除沉淀。
最后用PBS(PH7.2) 4℃透析过夜。
一、菌体的裂解1、怎样裂解细菌?细胞的破碎方法1.高速组织捣碎:将材料配成稀糊状液,放置于筒约1/3体积,盖紧筒盖,将调速器先拨至最慢处,开动开关后,逐步加速至所需速度。
此法适用于动物脏组织、植物肉质种子等。
2.玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。
3.超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料,用大肠杆菌制备各种酶,常选用50-100毫克菌体/毫升浓度,在1KG至10KG 频率下处理10-15分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施,时间以及超声间歇时间、超声时间可以自己调整,超声完全了菌液应该变清亮,如果不放心可以在显微镜下观察。
对超声波及热敏感的蛋白和核酸应慎用。
4.反复冻融法:将细胞在-20度以下冰冻,室温融解,反复几次,由于细胞冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。
5.化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好,我用的浓度一般为1mg/ml。
无论用哪一种方法破碎组织细胞,都会使细胞蛋白质或核酸水解酶释放到溶液中,使大分子生物降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以抑制那些活性中心需要有疏基的蛋白水解酶的活性,加入苯甲磺酰氟化物(PMSF)也能清除蛋白水解酶活力,但不是全部,而且应该在破碎的同时多加几次;另外,还可通过选择pH、温度或离子强度等,使这些条件都要适合于目的物质的提取。
这是标准配方:裂解液:50mM Tris-HCl(pH8.5~9.0),2mM EDTA,100mM NaCl,0.5%Triton X-100,1mg/ml溶菌酶。
(溶菌酶在这个pH围比较好发挥作用)但我个人的经验是:如果你裂解细菌是为了提取蛋白的话,而且蛋白的分子量又小于20kd的话,尽量减少溶菌酶的用量,会引入溶菌酶这种杂蛋白.一般配60ml裂解液用药匙匙柄盛一点就够.判断裂解好坏的标准是,溶液很粘.protocol是10ml-50ml缓冲液(菌体洗涤液,裂解液等)/1g湿菌体.如果只做一个鉴定,我觉得100-200ml菌就够了.但凡超声,我都用60ml裂解液,因为我们的超声仪(现代分子生物学实验技术录象里的那种)很适合用100ml小烧杯,装60ml裂解液,这样能让超声头离液面不高不低,不会冒泡泡,也不会洒出来.菌多我就延长超声时间.沉淀,也就是包涵体沉淀了,如果要上柱纯化,一定要先用4M尿素洗涤一下再用8M尿素溶解.如果不上柱,只是跑跑电泳,可以直接用8M尿素溶解以后,离心取上清,加入适量体积的loading buffer.loading buffer对于包涵体的溶解能力是较弱的."取200微升菌液,离心后直接加上样buffer,100度3分钟后上样,然后SDSPAGE.这个方法到底能不能溶解细菌中的包涵体?"而楼主的问题,虽然loading buffer对于包涵体的溶解能力是较弱的,但是我觉得你的做法只是在鉴定有无表达,用loading buffer是没有问题的.2、表达重组蛋白时,细菌裂解方法都有哪些?在表达重组蛋白时,诱导以后跑SDS-PAGE发现表达都很好,但是在裂解细胞时遇到问题。
包涵体蛋白的纯化和复性包涵体蛋白的纯化和复性1.菌体的收集和破碎用50 ml离心管分别收集诱导表达后的培养物,8000 rpm 4℃离心10 min。
沉淀用适量的超声破菌缓冲液重悬。
【备注】超声破菌缓冲液(20 mmol/L Tris-HCl pH8.0、0.5 mol/L NaCl、1 mmol/L EDTA、1 mg/mL溶菌酶)重悬后的菌液于冰浴中进行超声波破碎,其条件为:功率为300W,占空比50%,每个循环30 s,总时间20 min。
破碎后的匀浆,4℃、 12000 rpm离心15 min。
分别取上清液和沉淀进行12% SDS-PAGE分析,确定目的蛋白是否以包涵体的形式表达。
2.包涵体的处理洗涤:沉淀用适量的包涵体洗涤缓冲液重悬后,搅拌洗涤20~30 min,于4℃,12000 rpm离心15 min,弃去上清液。
重复一次。
再用适量的50 mmol/L Tris pH8.0溶液洗涤一遍(以去除残留的EDTA),于4℃ 12000 rpm离心15 min,弃去上清液。
【备注】包涵体洗涤缓冲液(20 mmol/L Tris-HCl pH 8.0,0.5 mol/L NaCl,2 mol/L尿素,2% Triton)2%Triton X-100溶液:量取2mlTriton X-100(聚乙二醇辛基苯基醚)液,加M缓冲液98ml即可。
溶解:沉淀用适量的包涵体溶解缓冲液重悬,室温搅拌5-6小时或过夜。
12000 rpm 4℃离心15 min,收集上清液。
【备注】包涵体溶解缓冲液(20 mmol/L Tris-HCl pH 8.0,0.5 mol/L NaCl,8 mol/L尿素,0.2 mmol/L DTT或100 mmol/L β-巯基乙醇,2%Triton)3.目的蛋白的纯化亲和层析基于目的蛋白与固相化的配基特异结合而滞留,其他杂蛋白会流过柱子。
谷胱甘肽S-转移酶(GST)是最常用的亲和层析纯化标签之一,带有此标签的重组蛋白可用交联谷胱甘肽的层析介质纯化,但本方法有以下缺点:首先,蛋白上的GST必须能合适的折叠,形成与谷胱甘肽结合的空间结构才能用此方法纯化;其次,GST标签多达220个氨基酸,如此大的标签可能会影响表达蛋白的可溶性,使形成包涵体,这会破坏蛋白的天然结构,难于进行结构分析,有时即便纯化后再酶切去除GST标签也不一定能解决问题。
板块一、大肠杆菌(这里讨论的大肠杆菌为非分泌到培养基中的重组蛋白,是否有重组蛋白分泌到培养基中的工程菌我没有见过。
)一、关于菌体的量大肠杆菌表达的基因工程蛋白是纯化人员最方便获得的原料,对纯化工艺开发来说几乎没有原料方面的限制.常看到有战友用个几毫升的菌液去做纯化,对此我十分不解,同样要做,为什么不多做点呢?很少的菌体会给纯化带来一些难以估计的问题,工艺的重复性和放大往往出现问题。
因此,要做个好工艺就多发酵表达一些菌体吧.我做纯化时,初始工艺摸索用的菌体量一般为10g左右.二、关于是否包涵体表达包涵体的定义我就不讲了。
我要讲的是,一个基因工程蛋白是否是包涵体表达的说法本身就不完全准确。
至于包涵体在电镜下的晶体颗粒表现等等对我们纯化来说毫无意义,我相信做纯化工艺的人没有谁去看这个电镜,也不关心.我们判断的依据只是SDS—PAGE,目的蛋白在破菌沉淀中,我们就认为是包涵体表达,但这是一个似是而非的结论。
看着没问题,实际上是有毛病的。
关键在于你用的是什么破菌缓冲液!有些蛋白在用缓冲液A破菌时是在破菌沉淀中,而用缓冲液B破菌时却在破菌上清中。
缓冲液A和B的差别可能只是pH上相差1-2个单位。
那么它是包涵体表达还是可溶上清表达呢?说这个问题主要在于有些战友往往非常在意他的目标蛋白是否包涵体表达。
甚至还有包涵体表达就用专门的包涵体蛋白纯化方法等等.我们应该关心的是目标蛋白在什么缓冲体系下是可溶的,在什么缓冲体系下是不溶的!不要让包涵体这个概念给你误导。
三、关于表达量我们常常在发表的文章上看到,我这个工程菌的目标蛋白的表达量达到菌体总蛋白的30%、50%等等。
我要说都是文章的作者在忽悠.不知道他们是如何定量的,用的最多的大概就是SDS-PAGE的扫描分析吧。
且不说一个SDS-PAGE不能表现出所有的菌体蛋白,电泳的染色方法、染色脱色强度、照片的曝光强度、扫描分析时的条带选择等等无不对这个百分比影响巨大。
在公司的QC部门做的对此应该最有体验,20%的条带要它变成30%又有何难?我的观点是对待表达量的描述不可定量,只能定性。