崔海莹 - 东北师范大学生命科学学院
- 格式:doc
- 大小:85.50 KB
- 文档页数:2
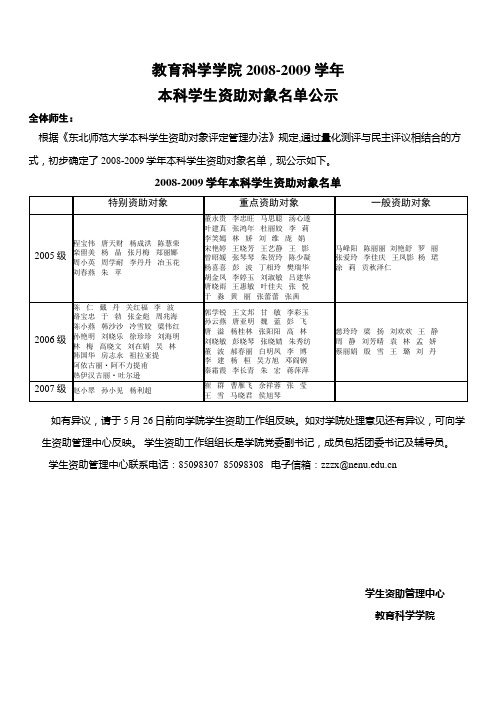
教育科学学院2008-2009学年本科学生资助对象名单公示全体师生:根据《东北师范大学本科学生资助对象评定管理办法》规定,通过量化测评与民主评议相结合的方式,初步确定了2008-2009学年本科学生资助对象名单,现公示如下。
2008-2009学年本科学生资助对象名单如有异议,请于5月26日前向学院学生资助工作组反映。
如对学院处理意见还有异议,可向学生资助管理中心反映。
学生资助工作组组长是学院党委副书记,成员包括团委书记及辅导员。
学生资助管理中心联系电话:85098307 85098308 电子信箱:zzzx@学生资助管理中心教育科学学院2008年5月23日政法学院2008-2009学年本科学生资助对象名单公示全体师生:根据《东北师范大学本科学生资助对象评定管理办法》规定,通过量化测评与民主评议相结合的方式,初步确定了2008-2009学年本科学生资助对象名单,现公示如下。
2008-2009学年本科学生资助对象名单如有异议,请于5月26日前向学院学生资助工作组反映。
如对学院处理意见还有异议,可向学生资助管理中心反映。
学生资助工作组组长是学院党委副书记,成员包括团委书记及辅导员。
学生资助管理中心联系电话:85098307 85098308 电子信箱:zzzx@学生资助管理中心政法学院2008年5月23日经济学院2008-2009学年本科学生资助对象名单公示全体师生:根据《东北师范大学本科学生资助对象评定管理办法》规定,通过量化测评与民主评议相结合的方式,初步确定了2008-2009学年本科学生资助对象名单,现公示如下。
2008-2009学年本科学生资助对象名单如有异议,请于5月26日前向学院学生资助工作组反映。
如对学院处理意见还有异议,可向学生资助管理中心反映。
学生资助工作组组长是学院党委副书记,成员包括团委书记及辅导员。
学生资助管理中心联系电话:85098307 85098308 电子信箱:zzzx@学生资助管理中心经济学院2008年5月23日商学院2008-2009学年本科学生资助对象名单公示全体师生:根据《东北师范大学本科学生资助对象评定管理办法》规定,通过量化测评与民主评议相结合的方式,初步确定了2008-2009学年本科学生资助对象名单,现公示如下。

卵母细胞及其胚胎发育潜能相关的生物标记物邵丽;崔毓桂【摘要】卵母细胞发育潜能是在卵泡发育过程中逐步获得的,卵丘细胞(CCs)与卵母细胞紧密接触,卵泡液(FF)为卵母细胞提供良好的微环境.因此,CCs功能及FF成分组成可以反映卵母细胞的发育潜能.CCs为卵母细胞提供代谢所需的营养物质如丙酮酸、丙氨酸、胆固醇等,并在卵母细胞的成熟、排卵、受精过程中发挥重要的作用.FF作为卵巢体壁细胞与血浆之间的媒介,其内的代谢相关分子、激素类分子、活性氧分子及一些细胞因子与卵母细胞的成熟及发育潜能密切相关.综述与卵母细胞及其胚胎发育潜能相关的生物标记物,主要是CCs、FF中的相关因子,两者结合为胚胎学家选择最佳的卵子提供参考.【期刊名称】《国际生殖健康/计划生育杂志》【年(卷),期】2014(033)005【总页数】6页(P379-383,395)【关键词】卵母细胞;生物学标记;卵丘细胞;卵泡液;胚胎发育【作者】邵丽;崔毓桂【作者单位】210029 南京医科大学第一附属医院生殖医学中心;210029 南京医科大学第一附属医院生殖医学中心【正文语种】中文欧洲人类生殖医学和胚胎学会(ESHRE)第13次调查统计显示,欧洲2009年体外受精(IVF)临床妊娠率为32.9%[1]。
为了提高临床妊娠率,临床上常在一个周期植入多个胚胎,使多胎妊娠率高,后者则可引起流产、难产、产后出血、胎儿生长受限等并发症增多。
单胚胎移植(singleembryo transfer,SET)可以有效地降低多胎妊娠率,SET达到满意临床妊娠率的关键是获得优质胚胎。
目前评价卵母细胞和胚胎质量主要依赖于相应的形态学参数,如卵子直径、卵丘扩张程度、胚叶个数及胚胎碎片率等,然而这些形态学检查方法缺乏客观性及准确性[2]。
因此,缺乏准确、敏感的评价卵子及其胚胎质量的方法,是SET尚未被临床广泛接受的原因之一。
因而,植入前通过无创性手段预测卵母细胞的发育潜能以获得优质卵子及其高质量的胚胎,是胚胎学家面临的挑战。

师范院校开设保护生物学通识课程的必要性收稿日期:2018-09-01基金项目:国家基础科学人才培养基金(J1210070);东北师范大学本科教学质量与教学改革工程建设项目(131004003);东北师范大学教师教学发展基金项目(15B1XZJ004)作者简介:李海燕,女,吉林公主岭人,副教授,博士,主要研究方向为植物种群生态学和保护生物学。
近些年来,由于人类社会经济的快速发展和人类活动的不断增强,导致人类赖以生存的自然环境持续恶化,许多动植物资源处于濒危状态,物种灭绝的速率也达到了前所未有的程度,生物多样性面临着史无前例的威胁。
目前,保护生物学(Conservation Biology )作为研究生物多样性保护的科学,相关研究已经持续了数十年之久。
联合国宣布2010年为国际生物多样性年。
为了不断增强人们的保护意识和采取更多的保护行动,科学而系统地宣传保护生物学的各方面知识则尤为重要和紧迫。
在我国教育体系中,通识课程作为普通高等教育中相对于专业教育而言的部分,与专业教育一起构成了普通高等教育的全部内容[1-2]。
它为面向各专业学生开设、以人文素质与科学素质教育为核心的综合素质类教育课程,是综合素质教育的重要环节,对于促进高等教育学生的全面科学发展具有重要的意义[3]。
师范类院校作为以师范教育为主的高等院校,师范教育为普及教育和国民素质的提高发挥了巨大的作用。
在生物多样性危机愈演愈烈的现状下,在师范院校开设保护生物学通识课程,对于全面提高我国基础教育中教师群体的生物保护素养,在未来的教学活动和日常生活中渗透生物保护的思想,科学广泛地普及生物保护知识、增强人们的保护意识,实现生物多样性的可持续发展和利用具有深远的影响。
一、我国生物多样性研究和保护的关注程度改革开放以来,我国的经济迅速发展。
然而,迅猛的经济发展和原本数量庞大的人口,使我国的生物多样性资源和动植物栖息环境遭受了过度的利用和严重的破坏。
近年来,不断有高等院校开始开设保护生物学课程,越来越多的保护生物学家、生态学家和生物学家从事生物多样性的研究和保护工作;同时,我国也积极地开展了各级各类自然保护区的建设工作以及诸多濒危物种如大熊猫、朱鹮、百山祖冷杉、银杉等的拯救和保护工作。

浒苔多糖EPⅢ的分离与鉴定
吴明江;焦丽丽;孙永旭;李天宝;张丽萍
【期刊名称】《东北师大学报:自然科学版》
【年(卷),期】2007(39)1
【摘要】利用乙醇分级对冻融后的浒苔热水提取物中的多糖进行了分级,得到了三个级分.采用凝胶层析法对其中一个级分进行纯化后得EPⅢ,经Sepharose CL-
6B,DEAE-SepharoseCL-6B检测均为单一对称洗脱峰,紫外检测未见有蛋白吸收峰.采用Sepharose CL-6B柱层析法测得其相对分子质量为38 000.经气相色谱分析该组分的单糖组成为鼠李糖、木糖、甘露糖、半乳糖、葡萄糖,其摩尔比为
12.70∶1.00∶0.64∶0.32∶19.03.
【总页数】4页(P97-100)
【关键词】浒苔;多糖;分离;组成分析
【作者】吴明江;焦丽丽;孙永旭;李天宝;张丽萍
【作者单位】东北师范大学生命科学学院
【正文语种】中文
【中图分类】Q538
【相关文献】
1.缘管浒苔粗多糖的气相色谱-质谱分析和初步分离 [J], 徐宝芹;张立新;王帆
2.4种不同来源浒苔中多糖的提取分离及理化性质 [J], 齐晓辉;李红燕;郭守东;陈荫;陈艳丽;徐健;侯玉娇;毛文君
3.浒苔多糖体外抗犬冠状病毒临床分离株的作用研究 [J], 孙秋艳;沈美艳;朱明恩;李舫
4.江苏产浒苔多糖的分离纯化和成分分析 [J], 潘晓慧; 吴皓; 潘旻; 张焱; 韦小翠; 程建明
5.浒苔多糖的分离纯化及其结构和抗氧化活性研究 [J], 王晔;娄永江;龚芳芳;陈淑敏;张玉琦;刘英丹;孟天宇;李勇勇
因版权原因,仅展示原文概要,查看原文内容请购买。

*通信作者,E -m a i l :s u n w e i @n e n u .e d u .c n收稿日期:2020-02-11;修回日期:2020-08-03基金项目:国家重点研发计划(2016Y F C 0500602);国家自然科学基金项目(31570470,31870456)作者简介:凌小莉(1994-),女,重庆人,硕士,研究方向为全球变化与草地碳循环,E -m a i l :l i n gx l 720@n e n u .e d u .c n .D O I :10.16742/j .z gc d x b .20200036氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响凌小莉,史宝库,崔海莹,宋文政,孙 伟*(东北师范大学草地科学研究所/植被生态科学教育部重点实验室,吉林 长春 130024)摘要:为探讨氮磷添加对草地土壤团聚体组成及碳含量分布特征的影响,在松嫩草地开展了为期4年(2015~2018年)的养分添加实验,包括4个氮添加水平和2个磷添加水平,共计8个处理组合㊂于2018年8月采集土样,采用湿筛法分析土壤团聚体结构,进而测量全土和不同粒径团聚体总碳(T C )㊁有机碳(S O C )㊁可溶性有机碳(D O C )和微生物量碳(M B C )含量㊂研究结果表明:随着氮添加水平的增加,0~10c m 和10~30c m 土层中大粒径团聚体比例和团聚体稳定性显著提高,小粒径团聚体比例显著降低;磷添加的影响与之相反㊂0~10c m 土层中,微团聚体占比高于淤泥+粘土团聚体㊂10~30c m 土层中,各团聚体组成所占比例大小为:微团聚体>淤泥+粘土团聚体>大团聚体㊂氮磷添加显著提高了土壤全土和团聚体S O C 和D O C 含量㊂随着氮添加水平的增加,土壤全土和团聚体M B C 含量显著降低;磷添加则显著提高了M B C 含量㊂在所有处理中,淤泥+粘土团聚体的碳含量显著大于微团聚体的碳含量㊂土壤团聚体稳定性与S O C 和D O C 含量存在正相关关系,与M B C 含量呈显著负相关关系㊂研究结果表明,连续氮磷添加可以改变草甸草原土壤结构,提高土壤固碳能力㊂关键词:草甸草原;氮磷添加;土壤团聚体;土壤结构;土壤有机碳中图分类号:S 812.2 文献标志码:A 文章编号:1673-5021(2021)02-0054-10土壤碳库是陆地生态系统中最大的有机碳(S o i l o r g a n i c c a r b o n ,S O C )库㊂全球的碳储量约为1.0ˑ108P g (1P g =1015g ),约1500P g SO C 以土壤有机质(S o i l o r ga n i cm a t t e r ,S OM )的形态储存于1m 深的土壤中㊂全球S O C 库储量约为大气碳库(750P g)的2倍,是陆地生物量碳库的2.5倍[1]㊂随着国际社会对气候变化和温室气体减排的日趋重视,土壤碳固存成为当今全球变化研究的热点之一㊂土壤固碳的主要机制是土壤团聚体的物理保护和土壤矿物对S O C 的吸附作用[2]㊂其中,土壤团聚体是通过矿物颗粒与有机和无机物质结合形成的二次颗粒,是土壤结构的基本单位[3]㊂土壤的固碳功能主要是以土壤团聚体为载体,S O C 不同形式的转化贯穿于土壤团聚体形成㊁稳定及周转过程㊂改善和提高土壤团聚体稳定性是增加土壤固碳潜力的主要途径㊂土壤团聚体结构和稳定性与土壤关键营养元素可利用性之间存在显著关系㊂例如,氮沉降和施肥会通过改变土壤理化性质㊁土壤微生物组成和土壤碳输入量等,影响土壤团聚体结构和稳定性,进而影响土壤固碳潜力㊂土壤团聚体在不同氮添加水平下的变化规律尚无定论㊂有研究发现氮添加对土壤团聚体组成没有显著影响[4],也有研究发现氮添加显著提高了大粒径团聚体比例和团聚体稳定性[5]㊂此外,氮添加会改变土壤的p H 值,导致土壤酸化或盐渍化,对土壤结构产生不利影响[6]㊂有关磷添加及氮磷交互添加对土壤团聚体影响的研究相对较少㊂氮磷添加如何影响草地土壤团聚体的组成㊁碳含量和稳定性,其影响是否存在交互作用?不同土层土壤团聚体的组成㊁碳含量和稳定性对氮磷添加的响应是否一致氮磷添加背景下土壤团聚体的组成㊁碳含量和稳定性的变化对土壤碳汇功能又有哪些指示作用这些问题仍需探讨㊂草地是我国面积最大的陆地生态系统类型㊂草地S O C 库是陆地生态系统碳库的重要组成部分㊂松嫩草地位于我国东北松嫩平原西部,处于欧亚草原的最东端,是东北黑土区重要的绿色生态屏障㊂全球变化导致氮磷元素变化对松嫩盐碱化草甸草原土壤团聚体结构和稳定性的影响程度和作用机制尚不明晰㊂本实验研究了不同氮磷添加组合对不同土壤层团聚体组成和团聚体碳的分布特征的影响,进而探讨氮磷添加对松嫩草地土壤固碳能力的影响,为保持和提升土壤固碳措施的研发提供参考㊂45 第43卷 第2期V o l .43 N o .2 中 国 草 地 学 报C h i n e s e J o u r n a l o fG r a s s l a n d2021年2月F e b .20211材料与方法1.1研究区域概况研究区域位于吉林省长岭县种马场(44ʎ30'~ 44ʎ45'N㊁123ʎ31'~123ʎ56'E),海拔约137.8~ 144.8m,属于典型的半干旱半湿润区域,气候类型为温带大陆性季风气候㊂研究区域年均降水量为280~644mm,季节间分配不均匀,降水主要集中在6~8月;年蒸发量在1500~2000mm之间,为降水量的3~4倍㊂年均气温为4.6~6.4ħ,无霜期约140d[7]㊂松嫩草地土壤以碱土和草甸土为主,p H 值变化范围为8.0~9.0,土壤S O C㊁总氮㊁总磷㊁无机氮和有效磷含量分别为16.14g/k g㊁1.59g/k g㊁0.32g/k g㊁5.93m g/k g和0.96m g/k g㊂生长季土壤含水量为3.7%~39.1%[8]㊂植被类型为羊草草甸草原,优势种为羊草(L e y m u s c h i n e n s i s),伴生种有芦苇(P h r a g m i t e s a u s t r a l i s)㊁拂子茅(C a l a m a g r o s-t i s e p i g e o s)㊁虎尾草(C h l o r i s v i r g a t a)等[9]㊂1.2样地设置于2010年围封2h m2羊草草甸草原㊂于2015年春,将围封草地划分为5个小区,每个小区包括8个样方,每个样方随机分配1个氮磷添加处理组合㊂不同处理样方的土壤和植被本底条件一致,每个样方面积为5mˑ5m,样方间隔2m,以保证各处理间无相互影响㊂于2015年5月开始进行氮磷添加处理,氮添加量分别为0㊁5㊁10和20g/(m2㊃a),磷添加量分别为0和5g/(m2㊃a),共8个处理组合㊂每个处理添加的氮磷量平均分为5份,于生长季(5~9月)每月中旬用喷壶喷施,对照样方喷施等量的水分,以减少外加水分带来的影响㊂氮添加处理使用硝酸铵ʒ尿素(7ʒ3)混合肥,磷添加处理使用过磷酸钙㊂1.3土样采集于2018年8月底,在每个小区利用多点采集法选取10点,用直径2c m的土钻分别采集0~10c m 和10~30c m土层土壤样品,剔除石块㊁根系㊁蚯蚓等动植物残体,充分混匀后,将土壤鲜样置于-20ħ冰箱中保存㊂1.4土壤团聚体筛分采用湿筛法[10]进行土壤团聚体分筛:称取约200g新鲜土样,水浸5m i n,利用湿筛仪(A S200 b a s i cB,R e t s c h G m b H,H a a n,G e r m a n y)以及250μm和53μm套筛,将土壤团聚体分为大团聚体(>250μm)㊁微团聚体(53~250μm)和淤泥+粘土团聚体(<53μm)三类;所获土壤团聚体38ħ烘干后保存于-20ħ冰箱,用于土壤团聚体碳含量分析㊂称取约50g自然风干土样分析土壤团聚体组成,方法同上㊂1.5土壤碳含量测定全土和筛分后的土壤团聚体用锡纸包样测量土壤总碳(T o t a l c a r b o n,T C)含量㊂土样盐酸酸化,除去无机碳后测量S O C含量㊂可溶性有机碳(D i s-s o l v e do r g a n i c c a r b o n,D O C)采用硫酸钾(K2S O4)浸提[11]㊂碳含量均用总有机碳分析仪(E l e m e n t a r v a r i oT O C,E l e m e n t a r a n a l y s e ns y s t e m eG m b H,H a n a u,G e r m a n y)测定㊂1.6团聚体稳定性土壤团聚体稳定性主要用平均重量直径(M e a n w e i g h t d i a m e t e r,MWD)和几何平均直径(G e o m e t-r i cm e a nd i a m e t e r,GM D)表示,计算公式为:MWD=ðn i=1X iˑW i(1) GM D=e x p[ðn i=1(W iˑl nX i)](2)公式中:X i为团聚体平均直径,W i为任一粒径团聚体百分比㊂1.7数据分析运用S P S S软件(I B M S P S S S t a t i s t i c s20, S P S S,C h i c a g o,U S A)进行数据统计分析㊂利用多因素方差分析氮添加(N)㊁磷添加(P)㊁土壤层(D)㊁粒径(A)及其交互作用对土壤团聚体组成㊁T C㊁S O C㊁D O C和M B C的影响㊂利用单因素方差分析氮添加(磷添加)对土壤团聚体组成㊁T C㊁S O C㊁D O C 和M B C的影响㊂利用线性模型分析MWD㊁GM D 与S O C㊁D O C和M B C之间的相关性㊂使用S i g m a-p l o t12.5软件作图㊂所有数值采用平均值ʃ标准误表示㊂2结果与分析2.1氮磷添加对土壤团聚体组成及稳定性的影响0~10c m土层湿筛收集到粒径53~250μm(微团聚体)和<53μm(淤泥+粘土团聚体)两类团聚体,10~30c m土层得到粒径>250μm(大团聚体)㊁53~250μm和<53μm三类团聚体㊂由于粒径> 250μm团聚体的收集量很少,不够用于土壤团聚体碳含量分析,因此分析了全土及53~250μm和< 53μm两类团聚体中的碳含量㊂55凌小莉史宝库崔海莹等氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响氮磷添加显著影响土壤团聚体组成(P <0.05,表1)㊂0~10c m 土层中,在P 0和P 5处理下,随着氮添加水平的增加,微团聚体的比例显著提高(图1-a ),淤泥+粘土团聚体的比例显著降低(图1-b )㊂10~30c m 土层中,在P 0处理下,大团聚体的比例随着氮添加水平的增加显著增加(图2-a ),淤泥+粘土团聚体结果与之相反(图2-c )㊂同一氮添加水平下,磷添加结果与氮添加结果相反(图1㊁图2)㊂土壤团聚体组成比例随着土壤层的变化而变化(P<0.05,表1)㊂0~10c m 土层中,微团聚体的比例为55%~67%,淤泥+粘土团聚体的比例为33%~45%(图1)㊂10~30c m 土层中:微团聚体>淤泥+粘土团聚体>大团聚体;其中大团聚体的比例为4%~5%,微团聚体的比例为63%~66%,淤泥+粘土团聚体的比例为29%~33%(图2)㊂ 氮磷添加显著影响土壤团聚体稳定性(P<0.05,表1)㊂在P 0和P 5处理下,随着氮添加水平的增加,各土壤层团聚体的MWD (图3-a ㊁图3-c)以及0~10c m 土层中团聚体的GM D 呈现显著增加的趋势(图3-b )㊂在P 0处理下,10~30c m 土层中团聚体的GM D 随着氮添加水平的增加而显著增加(图3-d )㊂与氮添加处理相比,氮磷添加交互处理降低了各土壤层团聚体的MWD 和GM D (图3)㊂土壤团聚体稳定性在不同土壤层存在显著差异(P <0.05,表1),10~30c m 土层中团聚体的MWD和GM D 均显著大于0~10c m 土层(图3)㊂表1 氮磷添加对松嫩草地土壤团聚体结构及其碳含量影响的方差分析T a b l e 1 A n a l y s i s o f v a r i a n c e (Fv a l u e )f o r t h e e f f e c t s o fNa n dPa d d i t i o no n s o i l a g g r e ga t e s t r u c t u r e a n d c a rb o nc o n t e n t i n t h e S o n gn e n g r a s s l a n d 影响因子F a c t o r s 自由度d f F 值 Fv a l u e微团聚体53~250μm M i c r o a g g r e g a t e 淤泥+粘土团聚体<53μm S i l t +c l a y a g g r e g a t e 平均重量直径M e a nw e i g h t d i a m e t e r 几何平均直径G e o m e t r i cm e a n d i a m e t e r 总碳T o t a l c a r b o n 有机碳S o i l o r ga n i c c a rb o n 可溶性有机碳D i s s o l v e do r ga n i c c a rb o n 微生物量碳M i c r o b i a l b i o m a s s c a r b o n氮310.33*11.70*17.76*13.18*2.133.45*15.79*24.64*磷14.91*5.84*10.89*7.03*65.09*0.072.4344.58*粒径1----5847.81*868.08*1276.77*1608.12*深度124.27*105.84*1791.25*274.23*118.08*1032.92*316.71*1014.68*氮ˑ磷30.590.560.590.586.883.43*0.621.48氮ˑ粒径3----51.74*1.501.777.13*氮ˑ深度34.11*3.26*0.491.6311.04*3.72*1.320.90磷ˑ粒径1----104.93*16.38*3.3010.20*磷ˑ深度10.580.300.810.0124.63*3.781.4114.74*粒径ˑ深度1----1069.21*24.72*34.08*115.74*氮ˑ磷ˑ粒径3----3.550.722.630.84氮ˑ磷ˑ深度30.150.150.460.145.930.460.100.87氮ˑ粒径ˑ深度3----1.580.551.100.29磷ˑ粒径ˑ深度1----1.4510.29*4.87*3.09氮ˑ磷ˑ粒径ˑ深度3----4.601.880.431.32 注:*表示在0.05水平上显著相关㊂N o t e :*m e a n s i gn i f i c a n t c o r r e l a t i o na t 0.05l e v e l.数据结果为平均值ʃ标准误(n =5)㊂不同字母表示在同一磷添加水平下氮添加处理之间存在显著差异,下同D a t a a r e r e p o r t e d a sm e a n ʃs t a n d a r d e r r o r s (n =5).D i f f e r e n t l e t t e r s r e p r e s e n t s i g n i f i c a n t d i f f e r e n c e s a m o n gd i f fe r e n t n i t r o g e na d d i t i o n r a t e s u n d e r t h e s a m e p h o s ph o r u s t r e a t m e n t ,t h e s a m e b e l o w 图1 0~10c m 土层土壤团聚体结构对氮磷添加的响应F i g .1 T h e r e s p o n s e s o f s o i l a g g r e g a t e s s t r u c t u r e i n t h e 0~10c ms o i l l a ye r t oNa n dPa d d i t i o n65 中国草地学报 2021年 第43卷 第2期图2 10~30c m 土层土壤团聚体结构对氮磷添加的响应F i g .2 T h e r e s p o n s e s o f s o i l a g g r e g a t e s s t r u c t u r e i n t h e 10~30c ms o i l l a ye r t oNa n dPa d d i t i on *表示在相同氮添加水平下,磷添加的显著差异;*㊁**和***分别表示显著水平P <0.05㊁P <0.01和P <0.001;下同*r e p r e s e n t s a s i g n i f i c a n t d i f f e r e n c e i n p h o s p h o r u s a d d i t i o na t t h e s a m e n i t r o ge na d d i t i o n l e v e l ;*,**a n d ***r e p r e s e n t s i g n if i c a n t d i f f e r e n c e a tP <0.05,P <0.01,a n dP <0.001l e v e l s ,r e s p e c t i v e l y.S a m e a s b e l o w 图3 不同土壤层的土壤团聚体稳定性对氮磷添加的响应F i g .3 T h e r e s p o n s e s o f s o i l a g g r e g a t e s s t a b i l i t y i n t h e d i f f e r e n t s o i l l a ye r s t oNa n dPa d d i t i o n 2.2 土壤碳含量分布特征对氮磷添加的响应氮磷添加显著影响全土及团聚体碳含量(P<0.05,表1)㊂在P 0和P 5处理下,两个土层中微团聚体T C 含量随着氮添加水平的增加而显著降低(图4-b ㊁图4-e )㊂在P 0处理下,随着氮添加水平的增加,0~10c m 土层中全土和10~30c m 土层中淤泥+粘土团聚体T C 含量呈现先增加后降低的趋势(图4-a ㊁图4-f )㊂同一氮添加水平下,磷添加提高各土壤层全土以及0~10c m 土层中微团聚体T C 含量,降低各土壤层淤泥+粘土团聚体以及0~30c m 土层中微团聚体T C 含量(图4)㊂在P 0和P 5处理下,随着氮添加水平的增加,0~10c m 土层中全土S O C 含量呈显著增加的趋势(图4-g)㊂在P 0处理下,0~10c m 土层中微团聚体S O C 含量随着氮添加水平的增加而显著增加(图4-h ),而氮添加显著降低10~30c m 土层中微团聚体S O C 含量(图4-k)㊂相同氮添加处理下,磷添加提高0~10c m 土层中全土及微团聚体S O C 含量,降低0~10c m 土层中淤泥+粘土团聚体以及10~30c m 土层中全土㊁淤泥+粘土团聚体S O C 含量(图4)㊂在P 0和P 5处理下,各土壤层淤泥+粘土团聚体D O C 含量随着氮添加水平的增加而逐渐增加(图5-c ㊁图5-f )㊂在P 5处理下,随着氮添加水平增加,各土壤层全土和0~10c m 土层中微团聚体D O C 含量显著增加(图5-a ㊁图5-b ㊁图5-d)㊂与氮添加处理相比,氮磷添加交互处理提高了各土壤层全75 凌小莉 史宝库 崔海莹等 氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响图4 氮磷添加处理对全土和不同粒径团聚体总碳和有机碳含量的影响F i g.4 E f f e c t s o fNa n dPa d d i t i o no n t h e c o n t e n t s o f s o i l t o t a l c a r b o na n d s o i l o r g a n i c c a r b o n i nb u l ks o i l a n dd i f f e r e n t a g g r e ga t e s 土㊁微团聚体及10~30c m 土层中淤泥+粘土团聚体D O C 含量,降低了0~10c m 土层中淤泥+粘土团聚体D O C 含量(图5)㊂在P 0和P 5处理下,随着氮添加水平的增加,10~30c m 土层中全土和各粒径团聚体M B C 含量呈显著降低的趋势(图5-j ㊁图5-k ㊁图5-l )㊂0~10c m 土层中全土(P 0)和淤泥+粘土团聚体(P 5)M B C 含量结果与之相同(图5-g ㊁图5-i )㊂在同一氮添加水平下,磷添加提高了各土壤层全土和各粒径团聚体M B C 含量(图5)㊂土壤全土及团聚体碳含量在不同土壤层中存在差异(P <0.05,表1)㊂0~10c m 土层中全土及各粒径团聚体S O C ㊁D O C 和M B C 含量大于10~30c m 土层(图4㊁图5)㊂不同粒径团聚体碳含量不同(P <0.05,表1),各土壤层微团聚体碳含量小于淤泥+粘土团聚体(图4㊁图5)㊂2.3 土壤团聚体稳定性与有机碳、可溶性有机碳和微生物碳的相关性在P 0处理下,0~10c m 土层中团聚体MW D 和G M D 与S O C 和D O C 含量呈显著正相关关系(P <0.05,图6-a ㊁图6-b ㊁图6-g㊁图6-h );10~30c m 土层中团聚体MWD 和GM D 与M B C 含量显著负相关(P <0.05,图6-f ㊁图6-l )㊂在P 5处理下,10~30c m 土层中团聚体MWD 和GM D 与D O C 含量存在显著正相关关系(P<0.05,图6-e ㊁图6-k );0~10c m 土层中团聚体MWD 和GM D 与M B C 含量呈现显著负相关关系(P <0.05,图6-c ㊁图6-i)㊂85 中国草地学报 2021年 第43卷 第2期图5氮磷添加处理对全土和不同粒径团聚体可溶性有机碳和微生物量碳含量的影响F i g.5 E f f e c t s o fNa n dPa d d i t i o no n t h e c o n t e n t s o f d i s s o l v e do r g a n i c c a r b o na n dm i c r o b i a l b i o m a s s c a r b o n i nb u l ks o i l a n dd i f f e r e n t a g g r e g a t e s3讨论3.1氮磷添加改变土壤有机碳和活性碳组分的潜在机制本研究结果表明0~10c m与10~30c m土层中土壤碳含量存在显著差异㊂0~10c m土层中,水热条件和通气状况较好,且聚集了较多植物根系,有较高的凋落物输入,土壤微生物活性高㊂因此,0~ 10c m土层中碳含量明显高于10~30c m土层(图4㊁图5),这与Z h a n g等[12]的研究结果相同㊂本研究发现氮添加提高了0~10c m和10~30c m土层中S O C含量;同一氮添加水平下,磷添加也提高了0~ 10c m土层中S O C含量(图4)㊂相比于典型草原和荒漠草原,松嫩草甸草原第一生产力受水氮共同限制㊂氮添加有效缓解了氮限制,显著增加植物生产力和土壤凋落物输入量,进而增加S O C含量,而氮㊁磷元素的耦合添加可以通过增加根系生物量和分泌物来增加S O C含量[13]㊂土壤活性碳是土壤有机碳的重要组成部分,主要包括D O C和M B C,其移动快㊁稳定性差㊁易氧化和矿化,与植物和土壤微生物功能活性密切相关㊂D O C是陆地和水生生态系统中非常重要的活性化学成分和土壤微生物最重要的能量和物质来源,其影响生态系统的养分循环和微生物的新陈代谢㊂与M c D o w e l l等[14]的研究结果一致,本研究发现氮磷添加显著提高了土壤D O C含量(图5),可能是因为养分添加增加了根系周转速率和凋落物的输入量,并且加速了有机质分解㊂作为草地生态系统中极为重要的部分,土壤微生物参与了土壤碳氮循环和土壤有机质矿化㊂它们在有机质分解,养分转化和供95凌小莉史宝库崔海莹等氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响图6 土壤团聚体稳定性与土壤有机碳,可溶性有机碳和微生物量碳的相关关系F i g .6 C o r r e l a t i o nb e t w e e n s o i l a g g r e g a t e s s t a b i l i t y a n d s o i l o r ga n i c c a rb o n ,d i s s o l v e do r ga n i c c a rb o na n dm ic r o b i a l b i o m a s s c a r b o n 应过程中起着主导作用,是草原土壤质量变化的重要指标㊂已有研究发现,氮添加会改变土壤微生物学特性,显著影响土壤微生物量[15],并且氮添加对土壤微生物的影响与植被类型㊁土壤类型㊁氮添加量和氮添加时间长短相关㊂但大多数研究结果表明,氮添加降低了土壤微生物量[16],这与本研究结果一致㊂此外,本研究发现在相同氮添加处理下,磷添加显著提高了土壤M B C 含量(图5),与B i e d e r b e c k等[17]的研究结果一致,这可能是因为磷添加改变了土壤关键元素化学计量特征,促进植物根系及部分微生物对无机氮的同化与吸收,从而减缓氮添加的不利影响㊂此外,磷添加增加的根系生物量及根系分泌物,为微生物提供充足的碳源,促进了微生物的生长[18]㊂3.2 土壤团聚体与土壤固碳功能的关系国内外关于土壤团聚体结构筛分的方法包括干筛和湿筛两种方法㊂众多研究表明,采用干筛法分析土壤团聚体结构,>250μm 团聚体质量比重较大;而采用湿筛法,<250μm 团聚体比重更大[19]㊂本研究采用湿筛法,因此0~10c m 土层中没有大团聚体(图1),10~30c m 土层中大团聚体的比例小于微团聚体和淤泥+粘土团聚体的比例(图2)㊂不同土壤层团聚体结构不同,可能与土壤母质以及土壤受干扰程度存在差异有关㊂土壤团聚体粒径越大,土壤团聚体的MWD 和GM D 越大,因此,土壤团聚体稳定性越高,土壤抗侵蚀能力越强㊂与Z h a n g等[12]的研究结果不同,本研究发现0~10c m 土层中团聚体稳定性小于10~30c m 土层,这是因为0~06 中国草地学报 2021年 第43卷 第2期10c m土层中没有大团聚体㊂土壤团聚体与S O C存在反馈调节机制㊂土壤有机碳促进团聚体聚集进而提高团聚体稳定性;而团聚体可为土壤有机碳提供物理保护,减少微生物降解和干扰[20]㊂相比于全土,团聚体孔隙小,孔隙弯曲度小,基质对微细胞的扩散慢,空气㊁水㊁养分㊁酶和外部微生物进入团聚体阻碍作用大,团聚体S O C不易被微生物降解㊂因此,土壤团聚体中有更大的S O C储量[21]㊂研究表明,表层土壤中约90%的S O C存在于团聚体中[22]㊂根据土壤团聚体的层次模型理论,土壤有机质组分在团聚体形成过程中会发生分异㊂与D o n g等[23]的研究结果相同,本研究发现小粒径团聚体中S O C含量大于大粒径团聚体(图4)㊂团聚体粒径越小,比表面积越大,其表面吸附能力越高,致使小粒径团聚体保留更多S O C含量㊂大粒径团聚体中S O C以易分解㊁矿化的活性S O C为主,活性S O C的分解与矿化导致总S O C含量不断降低,而小粒径团聚体中的S O C相对稳定,不易被微生物分解,因此被认为是土壤中稳定的碳库[23]㊂也有研究发现土壤中S O C增加主要发生在大粒径团聚体中[22],造成研究结果不同的原因可能是研究地域土壤类型,母质及物种种类差异㊂尽管M B C在土壤有机质总量中的占比很小,但它在调节能量流动和养分循环中起着至关重要的作用[24]㊂土壤团聚体中M B C的分布特征也可在一定程度上表征土壤质量的变化㊂本研究结果发现小粒径团聚体中M B C含量大于大粒径团聚体,这可能与团聚体中微生物群落组成及微生物总量有关㊂但也有研究发现大粒径团聚体中M B C值最大[25],导致微生物碳在土壤团聚体中分布结果差异与土壤类型㊁植被类型和土地管理措施等密切相关㊂本研究结果表明,随着氮添加水平的增加,土壤团聚体MWD和GM D显著增加(图3),土壤团聚体稳定性显著提高㊂此外,我们还发现团聚体稳定性与土壤中碳含量显著相关(图6)㊂S O C作为土壤团聚体胶结剂之一[26],其浓度反映了S O C分解速率㊁阳离子在土壤溶液中的释放及其与土壤溶液中阳离子络合的能力[3],进而对土壤团聚体聚集有着直接影响㊂有研究发现,土壤团聚体稳定性随S O C 浓度的增加而增加[3]㊂氮添加显著增加土壤全土S O C含量(图4),促进小粒径团聚体聚集形成大粒径团聚体,进而增加土壤团聚体稳定性(图3)㊂然而,在相同氮添加处理下,磷添加削弱了氮添加的效应,可能是因为磷添加改变了土壤团聚体的其他影响因子,例如土壤动物种类或代谢活动,进而大粒径团聚体裂解增加,小粒径团聚体的比例增加,最终土壤团聚体稳定性降低㊂土壤团聚体稳定性除了与S O C含量相关,我们还发现MWD和GM D与M B C 呈显著负相关关系(图6)㊂微生物对土壤团聚体的影响可能主要体现在两个方面:(1)丛枝菌根真菌和放线菌等微生物的菌丝将土壤颗粒彼此机械地缠绕在一起而聚集形成土壤团聚体;(2)依靠微生物的细胞分泌物(如多糖等)对土壤颗粒的胶结作用而形成稳定性土壤团聚体㊂本研究发现土壤团聚体稳定性与M B C呈显著负相关关系,表明氮磷添加条件下,微生物对土壤团聚体的影响可能小于S O C对土壤团聚体的影响,具体的解释还需要深入研究微生物群落结构组成变化㊂氮磷添加不仅影响土壤团聚体稳定性,还影响土壤团聚体碳含量,各粒径团聚体S O C含量随氮磷添加而显著增加(图4㊁图5),这与已有研究结果一致[5,12,27]㊂因此,本研究结果表明氮添加促进大粒径团聚体聚集,提高土壤团聚体稳定性;各粒径团聚体S O C含量也显著提高,最终改善土壤结构,提高土壤固碳能力㊂4结论松嫩草地土壤不同层次团聚体结构组成存在差异:0~10c m土层中微团聚体的比例>淤泥+粘土团聚体的比例;10~30c m土层中土壤团聚体的比例大小为微团聚体>淤泥+粘土团聚体>大团聚体㊂10~30c m土层中团聚体稳定性大于0~10c m土层㊂随着氮添加水平的增加,土壤大粒径团聚体比例和团聚体稳定性显著提高,小粒径团聚体比例显著降低㊂相同氮添加处理下,磷添加提高了小粒径团聚体所占的比例,降低了大粒径团聚体比例和团聚体稳定性㊂氮磷添加显著提高了土壤全土和团聚体S O C和D O C含量㊂土壤全土和团聚体M B C含量随着氮添加水平的增加而显著降低㊂土壤团聚体稳定性与土壤S O C含量存在显著正相关关系㊂总体上,氮添加增加了土壤团聚体S O C含量,提高了土壤团聚体稳定性,增强了土壤固碳能力㊂参考文献(R e f e r e n c e s):[1] R a i c h JW,P o t t e r CS.G l o b a l p a t t e r n s o f c a r b o n d i o x i d e e m i s-s i o n s f r o m s o i l s[J].G l o b a lB i o g e o c h e m i c a lC y c l e s,1995,9(1):23-36.[2] B a l e s d e n t J,C h e n uC,B a l a b a n eM.R e l a t i o n s h i p o f s o i l o r g a n-16凌小莉史宝库崔海莹等氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响i cm a t t e r d y n a m i c s t o p h y s i c a l p r o t e c t i o na n d t i l l a ge [J ].S o i l a n dT i l l a g eR e s e a r c h ,2000,53(3-4):215-230.[3] B r o n i c kCJ ,L a l R.S o i l s t r u c t u r e a n dm a n a g e m e n t :Ar e v i e w [J ].G e o d e r m a ,2005,124(1-2):3-22.[4] E g a n G ,C r a w l e y M J ,F o r n a r a D A.E f f e c t so f l o n g-t e r m g r a s s l a n dm a n a g e m e n t o n t h e c a r b o n a n d n i t r o ge n p o o l s of d i f -f e r e n t s o i l ag g r e g a t e f r a c t i o n s [J ].S c i e n c e o f th eT o t a lE n v i -r o n m e n t ,2018,613-614.[5] G h o s h A ,B h a t t a c h a r y y aR ,M e e n a M C ,e ta l .L o n g-t e r m f e r t i l i z a t i o ne f f e c t so ns o i l o r g a n i cc a r b o ns e qu e s t r a t i o ni na n i n c e p t i s o l [J ].S o i la n d T i l l a g eR e s e a r c h ,2018,177:134-144.[6] H a yn e sRJ ,N a i d uR.I n f l u e n c e o f l i m e ,f e r t i l i z e r a n dm a n u r e a p p l i c a t i o n so ns o i lo r g a n i c m a t t e rc o n t e n ta n ds o i l p h ys i c a l c o n d i t i o n s :A r e v i e w [J ].N u t r i e n tC y c l i n g i n A g r o e c o s y s -t e m s ,1998,51(2):123-137.[7] C u iH ,W a n g Y ,J i a n g Q ,e t a l .C a r b o n i s o t o p e c o m po s i t i o n o f n i g h t t i m e l e a f -r e s p i r e dC O 2in t h e a g r i c u l t u r a l -p a s t o r a l z o n e o f t h eS o n g n e nP l a i n ,n o r t h e a s t C h i n a [J ].P L o SO N E ,2015,10(9):e 0137575.[8] S h iB ,X u W ,Z h u Y ,e ta l .H e t e r o g e n e i t y of g r a s s l a n ds o i l r e s p i r a t i o n :A n t a g o n i s t i c e f f e c t so f g r a z i n g a n dn i t r o ge na d d i -t i o n [J ].A g r i c u l t u r a la n d F o r e s t M e t e o r o l o g y ,2019,268:215-223.[9] 曲同宝,朱悦,于淼,等.不同放牧强度松嫩草地土壤细菌碳代谢指纹差异[J ].中国草地学报,2016,38(4):68-75.Q uT o n g b a o ,Z h uY u e ,Y uM i a o ,e t a l .C a r b o nm e t a b o l i c f i n -g e r p r i n t o f s o i l b a c t e r i a a f f e c t e d b y g r a z i n g i n t e n s i t y i n S o n g n e ns t e p p e [J ].C h i n e s eJ o u r n a l o f G r a s s l a n d ,2016,38(4):68-75.[10] W a n g R ,C r e a m e rC A ,W a n g X ,e t a l .T h eef f e c t so f a9-y e a r n i t r og e n a n dw a t e r a d d i t i o n o n s o i l a g g r e g a t e ph o s ph o r u s a n d s u l f u r a v a i l a b i l i t y i n a s e m i -a r i d g r a s s l a n d [J ].E c o l o g i c a l I n d i c a t o r s ,2016,61:806-814.[11] W e iC ,Y u Q ,B a iE ,e ta l .N i t r o g e nd e po s i t i o n w e a k e n s p l a n t -m i c r o b e i n t e r a c t i o n s i n g r a s s l a n d e c o s ys t e m s [J ].G l o b -a lC h a n g eB i o l o g y ,2013,19(12):3688-3697.[12] Z h a n g L ,W e i X ,H a oM ,e t a l .C h a n g e s i n a g g r e ga t e -a s s o c i -a t e do r g a n i c c a rb o n a n dn i t r o ge n af t e r 27y e a r s o f f e r t i l i z a t i o n i nad r y l a n da l f a l f ag r a s s l a n do nth eL o e s sP l a t e a uo fC hi n a [J ].J o u r n a l o f A r i dL a n d ,2015,7(4):429-437.[13] T r i b e r t i L ,N a s t r iA ,G i o r d a n i G ,e t a l .C a nm i n e r a l a n do r -g a n i c f e r t i l i z a t i o n h e l p s e q u e s t r a t e c a r b o n d i o x i d e i n c r o p l a n d [J ].E u r o p e a nJ o u r n a l o f A g r o n o m y ,2008,29(1):13-20.[14] M c D o w e l l W H ,C u r r i e W S ,A b e rJD ,e ta l .E f f e c t so fc h r o n i c n i t r o g e na m e nd me n t so n p r o d u c t i o nof d i s s o l v e do r -g a n i c c a r b o na n dn i t r o ge ni nf o r e s ts o i l s [J ].W a t e r ,A i r ,a n dS o i lP o l l u t i o n ,1998,105(1-2):175-182.[15] C h e nD ,L a nZ ,H uS ,e t a l .E f f e c t so f n i t r o ge ne n r i c h m e n t o nb e l o w g r o u n dc o mm u n i t i e s i n g r a s s l a n d :R e l a t i v er o l eof s o i l n i t r og e n a v a i l a b i l i t y v s .s o i l a c i d i f i c a t i o n [J ].S o i l B i o l o g ya n dB i o c h e m i s t r y ,2015,89:99-108.[16] C h a o W ,L i uD ,B a i E .D e c r e a s i n g s o i lm i c r o b i a l d i v e r s i t yi s a s s o c i a t e d w i t hd e c r e a s i n g m i c r o b i a lb i o m a s su n d e rn i t r o g e n a d d i t i o n [J ].S o i l B i o l o g y a n d B i o c h e m i s t r y ,2018,120:126-133.[17] B i e d e r b e c kV O ,C a m p b e l l CA ,Z e n t n e rRP .E f f e c t o f c r o pr o t a t i o na n df e r t i l i z a t i o no ns o m eb i o l o g i c a l p r o pe r t i e so fa l o a mi ns o u t h w e s t e r nS a s k a t c h e w a n [J ].C a n a d i a nJ o u r n a l of So i l S c i e n c e ,1984,64(3):355-367.[18] G o ya l S ,C h a n d e rK ,M u n d r a M C ,e t a l .I n f l u e n c eo f i n o r -g a n i c f e r t i l i z e r s a n do r g a n i c a m e n d m e n t s o n s o i l o r ga n i cm a t -t e r a n d s o i lm i c r ob i a l p r o p e r t i e s u n d e r t r o p ic a l c o nd i t i o n s [J ].B i o l o g y a n dFe r t i l i t y of S o i l s ,1999,29(2):196-200.[19] 田浩.土壤团聚体有机碳研究现状[J ].安徽农学通报,2012,18(7):112-113.T i a n H a o .R e s e a r c hs t a t u so fs o i la g g r e g a t eo r g a n i cc a r b o n [J ].A n h u i A gr i c u l t u r a lS c i e n c e B u l l e t i n ,2012,18(7):112-113.[20] S i x J ,E l l i o t tET ,P a u s t i a nK.S o i lm a c r o a g g r e ga t e t u r n o v e r a n dm i c r o a g g r e g a t e f o r m a t i o n :A m e c h a n i s mf o r C s e qu e s t r a -t i o nu n d e rn o -t i l l a g ea g r i c u l t u r e [J ].S o i lB i o l o g y a n d B i o -c h e m i s t r y ,2000,32(14):2099-2103.[21] Z h o n g XL ,L i JT ,L i XJ ,e t a l .P h y s i c a l p r o t e c t i o nb y so i l a g g r e g a t e ss t a b i l i z e ss o i lo r g a n i cc a r b o nu n d e rs i m u l a t e d N d e p o s i t i o n i nas u b t r o pi c a lf o r e s to fC h i n a [J ].G e o d e r m a ,2017,285:323-332.[22] A o y a m a M ,A n g e r sD A ,N 'D a y e g a m i yeA.P a r t i c u l a t ea n d m i n e r a l -a s s o c i a t e do r g a n i c m a t t e r i n w a t e r -s t a b l ea g g r e ga t e s a s a f f e c t e db y m i n e r a l f e r t i l i z e r a n d m a n u r ea p pl i c a t i o n s [J ].C a n a d i a nJ o u r n a l o f So i l S c i e n c e ,1999,79(2):295-302.[23] D o n g X ,H a o Q ,L iG ,e ta l .C o n t r a s te f f e c to f l o n g-t e r m f e r t i l i z a t i o no nS O Ca n dS I Cs t o c k s a n d d i s t r i b u t i o n i n d i f f e r -e n t s o i l p a r t i c l e -s i z e f r a c t i o n s [J ].J o u r n a l o f S o i l s a n dS e d i -m e n t s ,2017,17(4):1054-1063.[24] J i a n g X ,S h i X ,W r i g h tAL .S e a s o n a l v a r i a b i l i t y ofm i c r o b i a l b i o m a s s a s s o c i a t e dw i t ha g g r e g a t e s i na r i c e -b a s e de c o s y s t e m [J ].E u r o p e a nJ o u r n a l o f S o i lB i o l o g y ,2013,56:84-88.[25] W a n g S ,L iT ,Z h e n g Z .D i s t r i b u t i o no fm i c r o b i a lb i o m a s s a n d a c t i v i t y w i t h i ns o i l a g g r e g a t e sa sa f f e c t e db y te a p l a n t a -t i o na ge [J ].C a t e n a ,2017,153:1-8.[26] 时龙,郭艳菊,于双,等.不同补播模式对荒漠草原土壤团聚体稳定性的影响[J ].中国草地学报,2019,41(3):83-89.S h i L o n g ,G u oY a n j u ,Y uS h u a n g ,e t a l .E f f e c t s o f d i f f e r e n t r e s e e d i n gp a t t e r no ns o i l a g g r e g a t es t a b i l i t y i nd e s e r ts t e p pe [J ].C h i n e s eJ o u r n a l of G r a s s l a n d ,2019,41(3):83-89.[27] W a ng R ,D o r o d n i k o vM ,D i jk s t r aFA ,e t a l .S e n s i t i v i t i e s t o n i t r o g e na n d w a t e ra d d i t i o n v a r y a m o n g m i c r o b i a l g r o u ps w i t h i ns o i la g g r e g a t e s i nas e m i a r i d g r a s s l a n d [J ].B i o l o g ya n dF e r t i l i t y o f So i l s ,2016,53(1):129-140.26 中国草地学报 2021年 第43卷 第2期E f f e c t s o fN i t r o g e na n dP h o s p h o r u sA d d i t i o no n S o i lA g g r e ga t e s S t r u c t u r e a n dC a rb o nC o n t e n t i n S o n gn e nG r a s s l a n d L I N G X i a o -l i ,S H IB a o -k u ,C U IH a i -y i n g ,S O N G W e n -z h e n g,S U N W e i (I n s t i t u t e o f G r a s s l a n dS c i e n c e ,N o r t h e a s tN o r m a lU n i v e r s i t y /K e y L a b o r a t o r y o fV e g e t a t i o nE c o l o g y ,M i n i s t r y o f E d u c a t i o n ,C h a n gc h u n 130024,C h i n a )A b s t r a c t :T o e x p l o r e t h e e f f e c t s o f n u t r i e n t s ad d i t i o no n g r a s s l a n d s o i l a g g re g a t e c o m po s i t i o na n d c a r -b o nc o n t e n t d i s t r i b u t i o n ,d i f f e r e n t l e v e l s o f n i t r o g e n a n d p h o s p h o r u sw e r e a d d e d i n a f i e l d e x p e r i m e n t f o r 4y e a r s (2015~2018)i nS o n g n e n g r a s s l a n d .T h e r ew e r e e i g h t t r e a t m e n t s i n t h e e x p e r i m e n t ,i n c l u d i n g f o u r a n d t w o d i f f e r e n t l e v e l s o f n i t r o g e n a n d p h o s p h o r u s ,r e s p e c t i v e l y .S o i l a g g r e g a t e sw e r e c l a s s i f i e du s i n g we t s i e v i n g m e t h o d .T h ec o n t e n t so ft o t a lc a r b o n (T C ),o r g a n i cc a r b o n (S O C ),d i s s o l v e do r g a n i cc a r b o n (D O C ),a n dm i c r o b i a l b i o m a s s c a r b o n (M B C )of b u l k s o i l a n d ag g r e g a t e sw i t hd i f f e r e n t pa r t i c l e s i z e sw e r e a n a l y z e d .T h e r e s u l t s s h o w e d t h a t t h e p r o p o r t i o n o f a g g r e g a t e sw i t h l a r g e p a r t i c l e s i z e a n d a g g r e ga t e s t ab i l -i t y (0~10c ma n d10~30c m s o i l l a y e r s )w e r es i g n i f i c a n t l y i n c r e a s ed w i t ht he i n c r e a s i n g n i t r o g e nl e v e l ,w h i l e t h e a d d i t i o nof p h o s p h o r u s h a d t h eo p p o s i t ee f f e c t .I nt h es o i l l a y e r (0~10c m ),t h e p r o po r t i o no f m i c r o a g g r e g a t e sw a s h i g h e r t h a n t h a t o f s i l t +c l a y a g g r e g a t e s .I n t h e s o i l l a ye r (10~30c m ),t h e r a n kof i n d i v i d u a l ag g r e g a t e p r o p o r t i o nw a s sh o w na s f o l l o wi n g :m i c r o a g g r e g a t e s >s i l t +c l a y a g g r e g a t e s >m a c r o -a g g r e g a t e s .T h e a d d i t i o no f n i t r o g e na n d p h o s p h o r u s s i g n i f i c a n t l yi n c r e a s e dS O Ca n dD O Cc o n t e n t s i n t h e b u l ks o i l a n d a g g r e g a t e s .W i t h t h e i n c r e a s i n g n i t r o g e n l e v e l ,M B Cc o n t e n t o f t h eb u l ks o i l a n da g g r e g a t e s w a s s i g n i f i c a n t l y d e c r e a s e d ,w h e r e a s t h e a d d i t i o no f p h o s p h o r u sh a d t h eo p p o s i t e e f f e c t .A m o n g al l t r e a t -m e n t s ,c a r b o n c o n t e n t s i n t h e s i l t +c l a y a g g r e g a t e sw e r e s i g n i f i c a n t l y h i g h e r t h a n t h a t o fm i c r o a g g r e ga t e s .S o i l a g g r e g a t e s s t ab i l i t y w a s s i g n i f ic a n t l y co r r e l a t e dw i t hS O C ,D O Ca n dM B C c o n t e n t s .I n c o n c l u s i o n ,t h e a d d i t i o no f n i t r o g e n a n d p h o s p h o r u s c a n c h a n g e t h e s t r u c t u r e o f s o i l a n d i m p r o v e t h e s o i l c a r b o n f i x a t i o n a -b i l i t yi n t h e p r e s e n t a r e a .K e y wo r d s :M e a d o ws t e p p e ;N i t r o g e na n d p h o s p h o r u sa d d i t i o n ;S o i l a g g r e g a t e s ;S o i l s t r u c t u r e ;S o i l o r ga n i c c a rb o n ʌ责任编辑 吴 尧ɔ36 凌小莉 史宝库 崔海莹等 氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响。

核糖体RNA基因在海洋动物分子系统学中的应用
毕相东;侯林;刘晓惠;王雪;刘芳;阎晗;杨雷
【期刊名称】《应用与环境生物学报》
【年(卷),期】2005(11)6
【摘要】随着分子遗传标记在分子系统学中应用的发展,核糖体RNA基因因其特殊的组成结构和演化方式被广泛应用于海洋动物分子系统学研究中.本文系统地介绍了真核生物核糖体RNA基因的组成和性质及其近年来在海洋动物系统进化、分类和种质鉴定、遗传多样性研究等方面研究中的应用.从中可以看出,核糖体RNA 基因已经广泛应用于海洋动物的分子系统学研究中,但主要利用的是核糖体RNA基因的一级结构.
【总页数】5页(P779-783)
【关键词】核糖体RNA基因;海洋动物;分子系统学;真核生物
【作者】毕相东;侯林;刘晓惠;王雪;刘芳;阎晗;杨雷
【作者单位】辽宁师范大学生命科学学院
【正文语种】中文
【中图分类】Q178.53;Q75
【相关文献】
1.核糖体RNA基因在蕈菌分子系统学研究中的应用 [J], 盛伟;方晓阳;潘传奇
2.核糖体RNA基因在丝核菌分子系统学研究中的应用 [J], 于金凤;伍恩宇;夏海波
3.双壳纲动物核糖体RNA 18S-ITS1序列及其在分子系统发育研究中的应用 [J],
丁小雷;何毛贤;邓凤姣;张锡元
4.COⅠ基因在海洋动物分子系统学研究中的应用 [J], 毕相东;杨雷;侯俊利;白东清;董少杰;孙金辉;李永仁;侯林
5.RNA的二级结构在动物分子系统学研究中的应用 [J], 谢强;卜文俊
因版权原因,仅展示原文概要,查看原文内容请购买。
实验教学示范中心在人才培养模式体系中的作用与教学改革实践崔喜艳(),李海燕吉林农业大学生命科学学院,农业生物学与工程技术实验教学示范中心,长春,130118摘 要:实验教学示范中心的建设和运行,是加强高等学校本科教学工作提高教学质量的重要举措,在高素质人才的综合培养中起到举足轻重的作用。
农业生物学与工程技术省级实验教学示范中心经过十多年的建设和发展,在师资队伍、培养模式、管理机制、实验教学和仪器管理等方面,理顺管理体制,实现资源共享;建立可行的管理机制,保证人才培养的顺利实施;建立规范化、科学化的管理方法,整合优化各种教学资源,形成了符合现代农业生物学实验教学示范中心内涵的运行模式,在构建多层次、多方位和多元化人才培养模式体系中提供了重要的保障。
关键词:实验教学示范中心,运行模式,人才培养模式体系The Function of Experimental Teaching Demonstration Center in Talent Cultivation Mode and Teaching Reform PracticeCUI Xi -yan (),LI Hai -yanSchool of Life Sciences ,Jilin Agricultural University ,Experiment Center for Agri -biology and Engineering ,Changchun 130118,China高校生物学教学研究(电子版)2020年10月,10(5):37-42ISSN 2095-1574 CN 11-9307/R DOI 10.3868/j.issn 2095-1574.2020.05.009实践教学收稿日期:2020-03-18;修回日期:2020-06-22基金项目:国家级精品资源共享课建设项目;吉林省教育科学“十三五”一般规划课题(GH170252,GH170253);吉林省教育厅省级优秀教学团队项目(吉教高字〔2016〕35号)。
生命科学学院研究生学业奖学金评定细则2019根据《吉林大学研究生学业奖学金评审办法》(校发[2018]23号)精神的有关规定,结合生命科学学院的实际情况,特制定本细则。
一、学院评审委员会(按姓氏拼音排序)主任委员:于湘晖副主任委员:刘丽慧委员:崔帅、崔银秋、高波、高仁钧、李正强、孔维、马俊锋、滕利荣、吴永革、邢述、徐力、邹丽云、张永锋(学生代表)二、评选对象档案已调入我校的纳入全国研究生招生计划的全日制研究生(有固定工资收入除外)。
研究生有下列情形之一的,取消参评资格:1.在学校规定的时间内欠缴学费和住宿费的(已办理绿色通道的新生除外),在学校规定的时间内未注册的;2.学期考核不合格的;3.保留学籍的;4.受纪律处分仍在处分期的。
三、评选办法及组织程序研究生学业奖学金的评审采取评审委员会分配名额并监督、各部门评审小组组织初评推荐复审,最后学院评审委员会确定并公示上报的方式进行。
1、博士研究生奖励额度为100%。
2、硕士研究生一年级优先奖励保送研究生,剩余名额按考研总成绩排序(按学术型生物、学术型药学、学位型专业分别进行排序),按成绩高低分给考入我院的非专业调剂研究生。
3、硕士研究生二年级、三年级学业奖学金的评定根据学生人数按比例分配到各系或实验室,换算公式:(名额总数/学生总数)×各部门研究生数。
研究生指导教师以研究生管理系统里的指导教师为准。
4、评审流程:(1) 10.17-10.21公示《生命科学学院研究生学业奖学金评定细则2019》;(2) 10.17-10.21研究生登录“吉林大学研究生教育管理信息系统”(/login.jsp),在申请“研究生学业奖学金”模块中进行网上申请;《吉林大学研究生学业奖学金审批表》填写好,导师签字后,交给班长。
(3) 10.22前,各部门评审小组组织初评推荐复审,并将拟获奖名单报到生命科学学院研究生工作办公室邹丽云老师处;(4) 10.22-10.28对生命科学学院2019年研究生学业奖学金名单进行公示。