NF-kB信号转导通路简介.pptx
- 格式:ppt
- 大小:2.04 MB
- 文档页数:15

NF-KB与微循环障碍核因子-KB(nuclear factor-kappa B,NF-KB)•蛋白家族是一种多效性的转录因子,可以与多种基启动子部位的KB位点发生特异性的结合从而促进其转录表达。
其受氧化应激、细菌脂多糖,细胞因子等多种刺激而活化后,能调控前炎症性细胞因子、细胞表面受体、转录因子、粘附分子等的生成。
而这些刺激因素及其调控的因子与微循环障碍的发生、发展均有着密切的关系。
本文就NF-KB的组成结构,•活化调节及与微循环障碍的关系等方面做一综述,以期从一新的角度阐述微循环障碍发生的机制及改善的途径。
1.NF-KB的概述1.1 NF-KB/Rel蛋白家族及结构1986年,Sen 等首次从鼠B淋巴细胞核提取物中,发现一种能与免疫球蛋白K轻链基因增强子KB序列(GGGACTTTCC)特异结合,调节其基因表达的核蛋白因子,•称之为NF-KB。
随后大量的研究又陆续发现了NF-KB•家族的其它成员,•其构成亚基分别是NF-•KB1 (P50)、NF-KB2(P52)、P65(RelA)、c-Rel(Rel)、RelB等,因这些亚基的N-末端均崐有约300个氨基酸残基的Rel同源区(rel homology domain ,RHD)•,•故统称为NF-KB/Rel蛋白家族。
其RHD内含DNA 结合区,二聚体化区和核定位序列,分别具有与DNA KB序列结合、与同源或异源亚基二聚体化以及与NF-KB抑制蛋白(IKB)家族成员相互作用并携带核定位信号(NLS),参与活化的NF-KB由细胞质向细胞核的迅速移动等功能。
又根据结构、功能和合成方式的不同,Rel蛋白分为两类。
•一类为P50(•NF-•KB1)和P52(•NF-•KB2),•分别由含有C-末端锚蛋白重复序列(ahkrin ••repeat motif)的前体蛋白p105和p100通过ATP依赖蛋白水解过程裂解而形成。
该类蛋白含有RHD,但缺乏转录活性区,无独立激活基因转录的功能。



01定义02意义信号转导是指细胞外信号通过细胞膜上的受体,经过一系列细胞内信号分子的传递和放大,最终引起细胞生理反应的过程。
信号转导是细胞对外界刺激作出应答的关键环节,参与调控细胞生长、分化、凋亡等多种生理过程,对于维持机体稳态具有重要意义。
信号转导定义与意义信号分子与受体分类信号分子包括激素、神经递质、生长因子、细胞因子等多种类型,它们通过与受体结合来传递信息。
受体分类根据信号分子的不同,受体可分为离子通道型受体、酶联型受体和G蛋白偶联型受体等类型。
每种受体都有其特定的结构和功能,能够识别并结合相应的信号分子。
03通过G 蛋白偶联受体介导的信号转导途径,包括cAMP 信号通路、磷脂酰肌醇信号通路等。
G 蛋白偶联受体信号转导途径通过酶联受体介导的信号转导途径,包括酪氨酸激酶受体信号通路、丝氨酸/苏氨酸激酶受体信号通路等。
酶联受体信号转导途径通过细胞因子受体介导的信号转导途径,包括JAK-STAT 信号通路、NF-κB信号通路等。
细胞因子受体信号转导途径信号转导途径简介信号的逐级放大细胞内信号传递过程中,信号分子通过级联反应逐级放大,使微弱的细胞外信号能够引起强烈的细胞生理反应。
信号的可调性细胞内信号传递过程受到多种因素的调节,包括受体表达水平、信号分子的合成与降解、信号转导蛋白的活性与定位等,这些调节机制使细胞能够对外界刺激作出精确而灵活的应答。
信号的整合性细胞内存在多种信号转导途径,这些途径之间通过交叉对话和相互调控,实现对细胞生理功能的整体协调和控制。
信号的特异性细胞内信号传递具有高度的特异性,不同的信号分子只能激活特定的信号转导途径,引起特定的细胞生理反应。
细胞内信号传递特点G蛋白偶联受体介导通路G蛋白偶联受体(GPCR)是一大类膜蛋白受体的统称,介导细胞对多种信号分子的响应。
GPCR与G蛋白结合后,通过激活或抑制下游效应器酶,将信号传递至细胞内。
常见的GPCR介导的信号转导通路包括cAMP信号通路、磷脂酰肌醇信号通路等。


NF-KB与微循环障碍核因子-KB(nuclear factor-kappa B,NF-KB)•蛋白家族是一种多效性的转录因子,可以与多种基启动子部位的KB位点发生特异性的结合从而促进其转录表达。
其受氧化应激、细菌脂多糖,细胞因子等多种刺激而活化后,能调控前炎症性细胞因子、细胞表面受体、转录因子、粘附分子等的生成。
而这些刺激因素及其调控的因子与微循环障碍的发生、发展均有着密切的关系。
本文就NF-KB的组成结构,•活化调节及与微循环障碍的关系等方面做一综述,以期从一新的角度阐述微循环障碍发生的机制及改善的途径。
1.NF-KB的概述1.1 NF-KB/Rel蛋白家族及结构1986年,Sen 等首次从鼠B淋巴细胞核提取物中,发现一种能与免疫球蛋白K轻链基因增强子KB序列(GGGACTTTCC)特异结合,调节其基因表达的核蛋白因子,•称之为NF-KB。
随后大量的研究又陆续发现了NF-KB•家族的其它成员,•其构成亚基分别是NF-•KB1 (P50)、NF-KB2(P52)、P65(RelA)、c-Rel(Rel)、RelB等,因这些亚基的N-末端均崐有约300个氨基酸残基的Rel同源区(rel homology domain ,RHD)•,•故统称为NF-KB/Rel蛋白家族。
其RHD内含DNA结合区,二聚体化区和核定位序列,分别具有与DNA KB序列结合、与同源或异源亚基二聚体化以及与NF-KB抑制蛋白(IKB)家族成员相互作用并携带核定位信号(NLS),参与活化的NF-KB由细胞质向细胞核的迅速移动等功能。
又根据结构、功能和合成方式的不同,Rel蛋白分为两类。
•一类为P50(•NF-•KB1)和P52(•NF-•KB2),•分别由含有C-末端锚蛋白重复序列(ahkrin ••repeat motif)的前体蛋白p105和p100通过ATP依赖蛋白水解过程裂解而形成。
该类蛋白含有RHD,但缺乏转录活性区,无独立激活基因转录的功能。
NF-KB与微循环障碍核因子-KB(nuclear factor-kappa B,NF-KB)•蛋白家族是一种多效性的转录因子,可以与多种基启动子部位的KB位点发生特异性的结合从而促进其转录表达。
其受氧化应激、细菌脂多糖,细胞因子等多种刺激而活化后,能调控前炎症性细胞因子、细胞表面受体、转录因子、粘附分子等的生成。
而这些刺激因素及其调控的因子与微循环障碍的发生、发展均有着密切的关系。
本文就NF-KB的组成结构,•活化调节及与微循环障碍的关系等方面做一综述,以期从一新的角度阐述微循环障碍发生的机制及改善的途径。
1.NF-KB的概述1.1 NF-KB/Rel蛋白家族及结构1986年,Sen 等首次从鼠B淋巴细胞核提取物中,发现一种能与免疫球蛋白K轻链基因增强子KB序列(GGGACTTTCC)特异结合,调节其基因表达的核蛋白因子,•称之为NF-KB。
随后大量的研究又陆续发现了NF-KB•家族的其它成员,•其构成亚基分别是NF-•KB1 (P50)、NF-KB2(P52)、P65(RelA)、c-Rel(Rel)、RelB等,因这些亚基的N-末端均崐有约300个氨基酸残基的Rel同源区(rel homology domain ,RHD)•,•故统称为NF-KB/Rel蛋白家族。
其RHD内含DNA 结合区,二聚体化区和核定位序列,分别具有与DNA KB序列结合、与同源或异源亚基二聚体化以及与NF-KB抑制蛋白(IKB)家族成员相互作用并携带核定位信号(NLS),参与活化的NF-KB由细胞质向细胞核的迅速移动等功能。
又根据结构、功能和合成方式的不同,Rel蛋白分为两类。
•一类为P50(•NF-•KB1)和P52(•NF-•KB2),•分别由含有C-末端锚蛋白重复序列(ahkrin ••repeat motif)的前体蛋白p105和p100通过ATP依赖蛋白水解过程裂解而形成。
该类蛋白含有RHD,但缺乏转录活性区,无独立激活基因转录的功能。


(完整版)NF-kb信号通路NF-KB与微循环障碍核因子-KB(nuclear factor-kappa B,NF-KB)?蛋白家族是一种多效性的转录因子,可以与多种基启动子部位的KB位点发生特异性的结合从而促进其转录表达。
其受氧化应激、细菌脂多糖,细胞因子等多种刺激而活化后,能调控前炎症性细胞因子、细胞表面受体、转录因子、粘附分子等的生成。
而这些刺激因素及其调控的因子与微循环障碍的发生、发展均有着密切的关系。
本文就NF-KB的组成结构,?活化调节及与微循环障碍的关系等方面做一综述,以期从一新的角度阐述微循环障碍发生的机制及改善的途径。
1.NF-KB的概述1.1 NF-KB/Rel蛋白家族及结构1986年,Sen 等首次从鼠B淋巴细胞核提取物中,发现一种能与免疫球蛋白K轻链基因增强子KB序列(GGGACTTTCC)特异结合,调节其基因表达的核蛋白因子,?称之为NF-KB。
随后大量的研究又陆续发现了NF-KB?家族的其它成员,?其构成亚基分别是NF-?KB1 (P50)、NF-KB2(P52)、P65(RelA)、c-Rel (Rel)、RelB等,因这些亚基的N-末端均崐有约300个氨基酸残基的Rel同源区(rel homology domain ,RHD)?,?故统称为NF-KB/Rel 蛋白家族。
其RHD内含DNA 结合区,二聚体化区和核定位序列,分别具有与DNA KB序列结合、与同源或异源亚基二聚体化以及与NF-KB抑制蛋白(IKB)家族成员相互作用并携带核定位信号(NLS),参与活化的NF-KB由细胞质向细胞核的迅速移动等功能。
又根据结构、功能和合成方式的不同,Rel蛋白分为两类。
?一类为P50(?NF-?KB1)和P52(?NF-?KB2),?分别由含有C-末端锚蛋白重复序列(ahkrin ??repeat motif)的前体蛋白p105和p100通过ATP依赖蛋白水解过程裂解而形成。

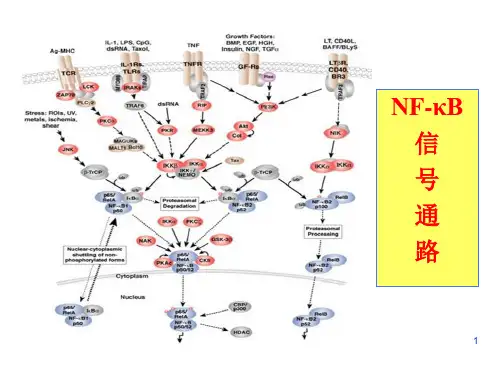
NF-kB信号通路-信号通路-生命维客——自由开放的生物医学交流平台NF-KB 信号通路图NF-κB信号通路实验解决方案(赞助商链接)NF-κB是一组重要的转录因子,参与细胞中多种生理过程,具有复杂的调节机制。
近年来的研究发现NF-κB在在肿瘤发生发展过程中具有重要而且复杂的作用,总体来说,NF-κB通路的异常激活可导致一系列与肿瘤相关基因的异常表达,从而抑制肿瘤细胞凋亡,促进正常细胞转化及肿瘤血管形成和转移等,直接影响恶性肿瘤的发生和发展,其抑制剂的开发亦为肿瘤治疗提供了一种新的选择。
NF-κB是一组具有特殊DNA结合序列的转录因子,主要调节炎症和自身免疫反应。
最近在基因工程小鼠中进行的研究表明,NF-κB对某些上皮来源器官的精确发育也具有重要作用[1]。
近些年很多研究发现NF-κB能够控制细胞增殖和癌变,调控细胞周期及凋亡,影响细胞分化,促进肿瘤转移。
在多数白血病和实体肿瘤中,NF-κB或IκB(Inhibitor of kappaB)家族成员往往通过遗传学改变或信号转导异常,而持续性激活或出现异常的核定位。
目录•NF-κB-Rel家族•NF-κB通路激活对肿瘤发生发展的促进作用•NF-kB通路激活对肿瘤发生发展的抑制作用•结语NF-κB-Rel家族编辑本段回目录NF-κB-Rel家族包括5个成员,即RelA(p65)、RelB、c-Rel、p105-p50(NF-κB1)和p100-p52(NF-κB2),在所有细胞中都有表达。
它们的N-末端均包含一个约300个氨基酸的高度同源序列,称为Rel 同源结构域(RHD),介导其与DNA结合及二聚化。
p105和p100通过蛋白酶解加工产生具有活性的DNA结合形式——p50 和 p52。
在未受到刺激的正常细胞中,NF-κB亚基与I-κBα、β或γ结合,以非活性形式被阻滞在胞浆中。
p105 和 p100的未加工形式的C-末端含有锚蛋白重复序列,此序列也能和I-κBs一样将p105 和 p100的二聚体阻滞在胞浆中。