[药学]第九章MAPK信号转导通路
- 格式:ppt
- 大小:3.68 MB
- 文档页数:122
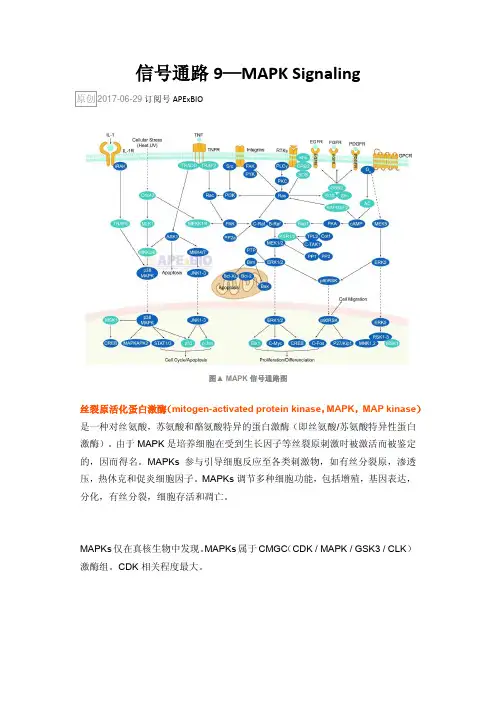
信号通路9—MAPK SignalingAPExBIO图▲ MAPK信号通路图丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK, MAP kinase)是一种对丝氨酸,苏氨酸和酪氨酸特异的蛋白激酶(即丝氨酸/苏氨酸特异性蛋白激酶)。
由于MAPK是培养细胞在受到生长因子等丝裂原刺激时被激活而被鉴定的,因而得名。
MAPKs参与引导细胞反应至各类刺激物,如有丝分裂原,渗透压,热休克和促炎细胞因子。
MAPKs调节多种细胞功能,包括增殖,基因表达,分化,有丝分裂,细胞存活和凋亡。
MAPKs仅在真核生物中发现。
MAPKs属于CMGC(CDK / MAPK / GSK3 / CLK)激酶组。
CDK相关程度最大。
MAPK链由3类蛋白激酶组成:上游激活蛋白→MAPK激酶激酶(MAPKKK)→MAPK激酶(MAPKK)→MAPK,通过依次磷酸化将上游信号传递至下游应答分子。
经典的MAPK通路激活开始于细胞膜,在这里,小GTP酶和各种蛋白激酶磷酸化并激活MAPKKK(MAP kinase kinase kinase,MAP3K或MKKK,MAPK激酶激酶)。
随后,MAPKKK直接磷酸化MAPKK(MAP kinase kinase,MAP2K 或MKK,MAPK激酶),MAPKK一旦被激活就会磷酸化并激活MAPK。
MAPK 的激活导致特异性MAPK激活蛋白激酶(MAPKAPK,MAPK-activated protein kinase)的磷酸化和活化,例如RSK,MSK或MNK家族成员和MK2/3/5。
MKKK的4个亚族已得到鉴定:A. Raf亚族。
研究的最为透彻,包括B-Raf、A-Raf、Raf1。
B. MEKK亚族。
由4种MEKK构成:MEKK1~MEKK4。
C. 第三个亚族:ASK1和Tpl2。
D. 第四个亚族与上述三个有较大不同,它包括MST(mammalian sterile 20-like)、SPRK、MUK(MAPK upstream kinase)、TAK1,以及相关程度最小的MOS (molony sarcoma oncoprotein)。

第九章细胞信号转导细胞通讯:一个信号产生细胞发出的信息通过介质(又称配体)传递到另一个靶细胞并与其相应的受体相互作用,然后通过信号转导产生靶细胞内一系列的生理生化变化,最终表现为靶细胞整体的生物学效应。
信号传导:是指信号分子从合成的细胞中释放出来,然后进行传递。
信号传导强调信号的产生、分泌与传送。
信号转导:是指信号的识别、转移与转换,包括配体与受体的结合、第二信使的产生及其后的级联反应等。
信号转导强调信号的接收与接收后信号转换的方式与结果。
受体:是一类能够结合细胞外特异性信号分子并启动细胞反应的蛋白质。
第二信使:细胞外信号分子不能进入细胞,它作用于细胞表面受体,经信号转导,在细胞内产生非蛋白类小分子,这种细胞内信号分子称为第二信使。
分子开关:细胞信号传递级联中,具有关闭和开启信号传递功能的分子。
信号通路:细胞接受外界信号,通过一整套特定机制,将胞外信号转化为胞内信号,最终调节特定基因表达,引起细胞的应答反应,这种反应系列称为细胞信号通路。
G蛋白偶联受体:指配体-受体复合物与靶细胞的作用是要通过与G蛋白的偶联,在细胞内产生第二信使,从而将细胞外信号跨膜传递到胞内影响细胞行为的受体。
cAMP信号通路:细胞外信号与细胞相应受体结合,导致细胞内第二信使cAMP水平的变化而引起细胞反应的信号通路。
(磷脂酰肌醇信号通路)双信使系统:胞外信号分子与细胞表面G蛋白偶联受体结合,激活膜上的磷脂激酶C,使质膜上的PIP2分解成IP3和DAG两个第二信使,将胞外信号转导为胞内信号,两个第二信使分别激活两种不同的信号通路,即IP3-Ca2+和DAG-PKC途径,实现对胞外信号的应答,因此将这种信号通路称为“双信使系统”。
钙调蛋白:真核细胞中普遍存在的Ca2+应答蛋白。
Ras蛋白:Ras基因的产物,分布于质膜胞质侧,结合GTP时为活化状态,结合GDP时失活状态,因此Ras蛋白属于GTP结合蛋白,具有GTP酶活性,具有分子开关的作用。

MAPK信号通路2008-06-04 21:50MAPK,丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是细胞内的一类丝氨酸/苏氨酸蛋白激酶。
研究证实,MAPKs信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。
研究表明,MAPKs信号转导通路在细胞内具有生物进化的高度保守性,在低等原核细胞和高等哺乳类细胞内,目前均已发现存在着多条并行的MAPKs信号通路,不同的细胞外刺激可使用不同的MAPKs信号通路,通过其相互调控而介导不同的细胞生物学反应。
1并行MAPKs信号通路的组成及其活化特点在哺乳类细胞目前已发现存在着下述三条并行的MAPKs信号通路[1]。
1.1ERK(extracellular signal-regulated kinase)信号通路1986年由Sturgill等人首先报告的MAPK。
最初其名称十分混乱,曾根据底物蛋白称之为MAP2K、ERK、MBPK、RSKK、ERTK等。
此后,由于发现其具有共同的结构和生化特征,而被命名为MAPK。
近年来,随着不同MAPK家族成员的发现,又重新改称为ERK。
在哺乳类动物细胞中,与ERK相关的细胞内信号转导途径被认为是经典MAPK信号转导途径,目前对其激活过程及生物学意义已有了较深入的认识。
研究证实,受体酪氨酸激酶、G蛋白偶联的受体和部分细胞因子受体均可激活ERK信号转导途径。
如:生长因子与细胞膜上的特异受体结合,可使受体形成二聚体,二聚化的受体使其自身酪氨酸激酶被激活;受体上磷酸化的酪氨酸又与位于胞膜上的生长因子受体结合蛋白2(Grb2)的SH2结构域相结合,而Grb2的SH3结构域则同时与鸟苷酸交换因子SOS(Son of Sevenless)结合,后者使小分子鸟苷酸结合蛋白Ras的GDP解离而结合GTP,从而激活Ras;激活的Ras进一步与丝/苏氨酸蛋白激酶Raf-1的氨基端结合,通过未知机制激活Raf-1;Raf-1可磷酸化MEK1/MEK2(MAP kinase/ERK kinase)上的二个调节性丝氨酸,从而激活MEKs;MEKs为双特异性激酶,可以使丝/苏氨酸和酪氨酸发生磷酸化,最终高度选择性地激活ERK1和ERK2(即p44MAPK和p42MAPK)。

MAPK 信号通路2008-06-04 21:50 MAPK, 丝裂原活化蛋白激酶( mitogen-activatedprotein kinases,MAPKs )是细胞内的一类丝氨酸/苏氨酸蛋白激酶。
研究证实,MAPKs 信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。
研究表明,MAPKs 信号转导通路在细胞内具有生物进化的高度保守性,在低等原核细胞和高等哺乳类细胞内,目前均已发现存在着多条并行的MAPKs 信号通路,不同的细胞外刺激可使用不同的MAPKs 信号通路,通过其相互调控而介导不同的细胞生物学反应。
1 并行MAPKs 信号通路的组成及其活化特点在哺乳类细胞目前已发现存在着下述三条并行的MAPKs 信号通路 [1]。
1.1 ERK (extracellular signal-regulated kinase)信号通路1986 年由Sturgill 等人首先报告的MAPK 。
最初其名称十分混乱,曾根据底物蛋白称之为MAP2K 、ERK、MBPK 、RSKK 、ERTK 等。
此后,由于发现其具有共同的结构和生化特征,而被命名为MAPK 。
近年来,随着不同MAPK 家族成员的发现,又重新改称为ERK 。
在哺乳类动物细胞中,与ERK 相关的细胞内信号转导途径被认为是经典MAPK 信号转导途径,目前对其激活过程及生物学意义已有了较深入的认识。
研究证实,受体酪氨酸激酶、G 蛋白偶联的受体和部分细胞因子受体均可激活ERK 信号转导途径。
如:生长因子与细胞膜上的特异受体结合,可使受体形成二聚体,二聚化的受体使其自身酪氨酸激酶被激活;受体上磷酸化的酪氨酸又与位于胞膜上的生长因子受体结合蛋白2( Grb2)的SH2 结构域相结合,而Grb2 的SH3 结构域则同时与鸟苷酸交换因子SOS( Son of Sevenless)结合,后者使小分子鸟苷酸结合蛋白Ras的GDP 解离而结合GTP,从而激活Ras;激活的Ras进一步与丝/苏氨酸蛋白激酶Raf-1 的氨基端结合,通过未知机制激活Raf-1;Raf-1 可磷酸化MEK1 /MEK2 (MAP kinase/ERK kinase)上的二个调节性丝氨酸,从而激活MEKs ;MEKs 为双特异性激酶,可以使丝/苏氨酸和酪氨酸发生磷酸化,最终高度选择性地激活ERK1和ERK2(即p44MAPK 和p42MAPK )。

“mapk信号转导通路”资料合集目录一、MAPK信号转导通路在肝细胞癌中的作用研究二、MAPK信号转导通路在肝细胞癌中的作用研究三、糖肾平胶囊对STZ诱导糖尿病肾病大鼠肾脏保护及其对TGF1p38MAPK信号转导通路的影响四、MAPK信号转导通路与神经损伤研究进展五、P,pDDE诱导ROS在线粒体和MAPK信号转导通路中的作用六、P38MAPK信号转导通路在大蒜素诱导THP1细胞凋亡中的作用七、MAPK信号转导通路中ERK、JNK和P38在大鼠肝脏缺血再灌注和缺血后处理中表达的变化八、MAPK信号转导通路及凋亡蛋白在子痫前期中的研究MAPK信号转导通路在肝细胞癌中的作用研究肝纤维化动物实验模型的研究进展肝纤维化是一种常见的慢性肝病,其特征是肝脏中胶原蛋白的过度积累。
为了更好地研究肝纤维化的发病机制和寻找有效的治疗方法,建立动物实验模型是至关重要的。
本文将综述近年来肝纤维化动物实验模型的研究进展。
一、肝纤维化动物实验模型概述肝纤维化动物模型主要用于模拟人类肝纤维化的发生和发展过程,以便更深入地了解其病理生理机制。
这些模型可以通过不同的方法建立,包括化学物质诱导、基因工程和无菌炎症等。
二、肝纤维化动物实验模型的建立方法1、化学物质诱导模型:通过给动物注射化学物质,如四氯化碳、二甲基亚硝胺等,来诱导肝脏损伤和纤维化。
这种方法操作简单,但化学物质对肝脏的损伤程度和纤维化进程的调控不够精确。
2、基因工程模型:通过基因工程技术,如转基因或基因敲除技术,来改变动物体内相关基因的表达,以模拟肝纤维化的发生。
这些模型具有更好的可控性和可重复性,但制备过程较为复杂。
3、无菌炎症模型:通过向动物体内注射无菌炎症因子,如脂多糖等,来模拟慢性炎症环境下的肝纤维化。
这种方法可以在一定程度上模拟人类肝纤维化的自然病程。
三、肝纤维化动物实验模型的应用肝纤维化动物实验模型在研究肝纤维化的发病机制、药物筛选和评价等方面具有广泛的应用。



MAPK/ERK信号通路图及简介日期:2013-01-23 来源:互联网标签:信号通路MAPK相关专题:MAPK信号通路专题摘要: MAPK,丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是细胞内的一类丝氨酸/苏氨酸蛋白激酶。
研究证实,MAPKs信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。
研究表明,MAPKs 信号转导通路在细胞内具有生物进化的高度保守性,在低等原核细胞和高等哺乳类天隆科技NP968自动核酸提取仪,产品试用进行中!佛山泰尔健生物细胞培养器材诚征代理MAPK,丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是细胞内的一类丝氨酸/苏氨酸蛋白激酶。
研究证实,MAPKs信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。
研究表明,MAPKs 信号转导通路在细胞内具有生物进化的高度保守性,在低等原核细胞和高等哺乳类细胞内,目前均已发现存在着多条并行的MAPKs信号通路,不同的细胞外刺激可使用不同的MAPKs信号通路,通过其相互调控而介导不同的细胞生物学反应。
ERK(extracellular signal-regulated kinase)信号通路1986年由Sturgill等人首先报告的MAPK。
最初其名称十分混乱,曾根据底物蛋白称之为MAP2K、ERK、MBPK、RSKK、ERTK等。
此后,由于发现其具有共同的结构和生化特征,而被命名为MAPK。
近年来,随着不同MAPK家族成员的发现,又重新改称为ERK。
哺乳类动物细胞中,与ERK相关的细胞内信号转导途径被认为是经典MAPK 信号转导途径,目前对其激活过程及生物学意义已有了较深入的认识。

肿瘤细胞的信号转导通路信号传导通路是将胞外刺激由细胞表面传入细胞内,启动了胞浆中的信号转导通路,通过多种途径将信号传递到胞核内,促进或抑制特定靶基因的表达。
一、MAPK信号通路MAPK信号通路介导细胞外信号到细胞内反应。
丝裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)主要位于细胞浆,很多生长因子所激活,活化后既可以磷酸化胞浆内的靶蛋白,也能进入细胞核作用于对应的转录因子,调节靶基因的表达。
调节着细胞的生长、分化、分裂、死亡各个阶段的生理活动以及细胞间功能同步化过程,并在细胞恶变和肿瘤侵袭转移过程中起重要作用,阻断MAPK途径是肿瘤侵袭转移的治疗新方向。
MAPK信号转导通路是需要经过多级激酶的级联反应,其中包括3个关键的激酶,即MAPK激酶激酶(MKKK)→MAPK激酶(MKK)→MAPK。
(一)MKKK:包括Raf、Mos、Tpl、SPAK、MUK、MLK和MEKK等,其中Raf又分为A-Raf、B-Raf、Raf-1等亚型;MKKK是一个Ser/Thr蛋白激酶,被MAPKKKK、小G蛋白家族成员Ras、Rho激活后可Ser/Thr磷酸化激活下游激酶MKK。
MKK识别下游MAPK分子中的TXY序列(“Thr-X-Tyr”模序,为MAPK第Ⅷ区存在的三肽序列Thr-Glu-Tyr、Thr-Pro-Tyr或Thr-Gly-Tyr),将该序列中的Thr和Tyr分别磷酸化后激活MAPK。
注:TXY序列是MKK活化JNK的双磷酸化位点,MKK4和MKK7通过磷酸化TXY 序列的第183位苏氨酸残基(Thr183)和第185位酪氨酸残基(Tyr185)激活JNK1。
(二)MKK:包括MEK1-MEK7,主要是MEK1/2;(三)MAPK:MAPK是一类丝氨酸/苏氨酸激酶,是MAPK途径的核心,它至少由4种同功酶组成,包括:细胞外信号调节激酶(Extracellular signal Regulated Kinases,ERK1/2)、C-Jun 氨基末端激酶(JNK)/应激激活蛋白激酶(Stress-activated protein kinase,SAPK)、p38(p38MAPK)、ERK5/BMK1(big MAP kinase1)等MAPK亚族,并根据此将MAPK 信号传导通路分为4条途径。

细胞生物综述MAPK/ERK信号转导通路与学习记忆丝裂素活化蛋白激酶(MAPK)是细胞内的一类丝氨酸/苏氨酸蛋白激酶,是多种细胞外信号从细胞表面传导到细胞内的重要传递者。
细胞外信号调节激酶(ERK)包括ERKl和 ERK2,又称p44 MAPK和p42 MAPK,相对分子质量分别为4400和4200,是MAPK家族中的重要成员。
ERK最初被认为和细胞的生长、发育、分化有关,然而在已经不需要增殖和分化的成熟神经元中发现仍然有大量ERK上游调控子和下游靶蛋白存在,因此人们开始探索ERK的其他作用。
1997年English和Sweatt首次发现了ERK和长时程增强(LTP) 效应的关系,他们采用ERK上游激酶的抑制剂PD 098059阻止了ERK 的激活,同时也显著阻止了海马CA1区LTP 的诱导。
该发现随后又被其他研究小组证实并扩展,从而确定ERK与LTP的关系。
大量研究表明,LTP和学习、记忆过程密切相关,被许多学者命名为“学习、记忆的突触模型”,因此确定了ERK与学习、记忆的关系。
1 MAPK/ERK三级级联反应及与学习记忆的关系MAPK/ERK信号转导通路采用高度保守的三级激酶级联传递信号即Raf/MEK/ERK,活化的Raf使MEKⅧ亚区的两个丝氨酸磷酸化而激活,MEK 激活后使 ERK的苏氨酸和酪氨酸双位点磷酸化而激活。
研究表明,经水迷宫训练后的大鼠,背侧海马CA1/CA2区ERK被激活,使用PD098059抑制背侧海马 MAPK/ERK级联反应则磷酸化ERK蛋白(即活性形式)含量降低,并且长期空间记忆的形成受损。
海马内注入PD098059的大鼠精细定位信息受损。
利用转基因方法使小鼠前脑MEK1突变而突变的MEK1仍然能够和ERK相互作用,但却不能使它激活,这些小鼠表现为空间参考记忆及长期关联性记忆受损。
另一研究小组发现,利用转基因方法使小鼠脑中的MEK1突变,则这些小鼠海马区ERK活性降低,同时表现出特殊的恐惧性条件反射损害,此为海马依赖性的学习形式之一。

MAPK信号通路2008-06-04 21:50MAPK,丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是细胞内的一类丝氨酸/苏氨酸蛋白激酶。
研究证实,MAPKs信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。
研究表明,MAPKs信号转导通路在细胞内具有生物进化的高度保守性,在低等原核细胞和高等哺乳类细胞内,目前均已发现存在着多条并行的MAPKs信号通路,不同的细胞外刺激可使用不同的MAPKs信号通路,通过其相互调控而介导不同的细胞生物学反应。
1并行MAPKs信号通路的组成及其活化特点在哺乳类细胞目前已发现存在着下述三条并行的MAPKs信号通路[1]。
1.1ERK(extracellular signal-regulated kinase)信号通路1986年由Sturgill等人首先报告的MAPK。
最初其名称十分混乱,曾根据底物蛋白称之为MAP2K、ERK、MBPK、RSKK、ERTK等。
此后,由于发现其具有共同的结构和生化特征,而被命名为MAPK。
近年来,随着不同MAPK家族成员的发现,又重新改称为ERK。
在哺乳类动物细胞中,与ERK相关的细胞内信号转导途径被认为是经典MAPK信号转导途径,目前对其激活过程及生物学意义已有了较深入的认识。
研究证实,受体酪氨酸激酶、G蛋白偶联的受体和部分细胞因子受体均可激活ERK信号转导途径。
如:生长因子与细胞膜上的特异受体结合,可使受体形成二聚体,二聚化的受体使其自身酪氨酸激酶被激活;受体上磷酸化的酪氨酸又与位于胞膜上的生长因子受体结合蛋白2(Grb2)的SH2结构域相结合,而Grb2的SH3结构域则同时与鸟苷酸交换因子SOS(Son of Sevenless)结合,后者使小分子鸟苷酸结合蛋白Ras的GDP解离而结合GTP,从而激活Ras;激活的Ras进一步与丝/苏氨酸蛋白激酶Raf-1的氨基端结合,通过未知机制激活Raf-1;Raf-1可磷酸化MEK1/MEK2(MAP kinase/ERK kinase)上的二个调节性丝氨酸,从而激活MEKs;MEKs为双特异性激酶,可以使丝/苏氨酸和酪氨酸发生磷酸化,最终高度选择性地激活ERK1和ERK2(即p44MAPK和p42MAPK)。
信号通路9—MAPK Signaling订阅号APExBIO图▲ MAPK信号通路图丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK, MAP kinase)是一种对丝氨酸,苏氨酸和酪氨酸特异的蛋白激酶(即丝氨酸/苏氨酸特异性蛋白激酶)。
由于MAPK是培养细胞在受到生长因子等丝裂原刺激时被激活而被鉴定的,因而得名。
MAPKs参与引导细胞反应至各类刺激物,如有丝分裂原,渗透压,热休克和促炎细胞因子。
MAPKs调节多种细胞功能,包括增殖,基因表达,分化,有丝分裂,细胞存活和凋亡。
MAPKs仅在真核生物中发现。
MAPKs属于CMGC(CDK / MAPK / GSK3 / CLK)激酶组。
CDK相关程度最大。
MAPK链由3类蛋白激酶组成:上游激活蛋白→MAPK激酶激酶(MAPKKK)→MAPK激酶(MAPKK)→MAPK,通过依次磷酸化将上游信号传递至下游应答分子。
经典的MAPK通路激活开始于细胞膜,在这里,小GTP酶和各种蛋白激酶磷酸化并激活MAPKKK(MAP kinase kinase kinase,MAP3K或MKKK,MAPK激酶激酶)。
随后,MAPKKK直接磷酸化MAPKK(MAP kinase kinase,MAP2K 或MKK,MAPK激酶),MAPKK一旦被激活就会磷酸化并激活MAPK。
MAPK 的激活导致特异性MAPK激活蛋白激酶(MAPKAPK,MAPK-activated protein kinase)的磷酸化和活化,例如RSK,MSK或MNK家族成员和MK2/3/5。
MKKK的4个亚族已得到鉴定:A. Raf亚族。
研究的最为透彻,包括B-Raf、A-Raf、Raf1。
B. MEKK亚族。
由4种MEKK构成:MEKK1~MEKK4。
C. 第三个亚族:ASK1和Tpl2。
D. 第四个亚族与上述三个有较大不同,它包括MST(mammalian sterile 20-like)、SPRK、MUK(MAPK upstream kinase)、TAK1,以及相关程度最小的MOS (molony sarcoma oncoprotein)。
MAPK信号转导途径及其功能蛋白质调节其功能/活性的方式有很多种,包括磷酸化-去磷酸化,乙酰化,蛋白质切割如酶原激活和caspases激活等。
蛋白质的磷酸化和去磷酸化是蛋白质调节其功能/活性的一种重要方式,有些蛋白质在磷酸化状态时具有活性,而在非磷酸化状态时没有活性,如激酶MAPK和转录因子CREB,Jun等,而有些蛋白质相反,在磷酸化状态时没有活性,而在非磷酸化状态时具有活性,如转录因子IκBα的抑制活性。
蛋白质通过磷酸化-去磷酸化调节功能/活性并进而影响细胞的很多生命过程。
我们在这里仅仅重点介绍MAPK超家族蛋白质的磷酸化-去磷酸化状态对一些重要生命过程的调节。
MAPK超家族信号转导途径MAPK超家族包括三个亚家族:ERK1/2,JNK/SAPK和p38 MAPK。
近10年来,利用培养的可分裂/分化的细胞系对MAPK超家族的信号转导途径和功能进行了详细深入的研究,其中相当一部分论文发表在诸如NATURE,SCIENCE,CELL,JBC,PNAS等著名杂志上,是近年来生命科学的研究热点之一。
研究表明ERK1/2信号转导途径主要对细胞的生长,分裂和分化信号进行传导,而JNK/SAPK信号转导途径和p38 MAPK信号转导途径主要对炎性细胞因子和多种类型的细胞应激信号进行传导。
当然这种划分是不很严格的,尤其是对于动物的神经系统。
一般地说,一种细胞外信号可以通过一种或一种以上的信号转导途径传导,而一种信号转导途径可以传导一种或一种以上细胞外信号。
信号转导途径间还存在crosstalk。
1.SAPK/JNK途径SAPK/JNK途径传导细胞应激,炎性细胞因子,紫外线,蛋白质合成抑制剂,渗透压应激等信号。
MEKK1,SEK1/MKK4/JNKK(Ser219/Thr223),SAPK/JNK(Thr183/Tyr185)激酶依次被激活。
活化的SAPK/JNK 激活转录因子c-Jun(Ser63/Ser73),c-Jun结合到TRE/AP-1元件启动转录;活化的SAPK/JNK也可激活转录因子ATF-2/CRE-BP1(Thr69/Thr71),ATF-2结合到AP-1和CRE DNA反应元件启动转录;活化的SAPK/JNK还可激活转录因子Elk1。
MAPK级联激活信号通路丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)是一组能被不同的细胞外刺激,如细胞因子、神经递质、激素、细胞应激及细胞黏附等激活的丝氨酸-苏氨酸蛋白激酶。
MAPK信号通路是利用逐级磷酸化过程将信号放大传至细胞核内,调节转录因子的活性,调控相应基因的表达,进而引起细胞反应的一类重要信号系统。
MAPK级联激活是多种信号通路的中心,参与到细胞的分裂、分化、凋亡等多种生命过程,且在一些骨关节组织炎症,癌细胞转移等病症中发挥关键性作用。
MAPK信号转导是以3 级激酶级联激活的方式进行的,如图所示,首先MAPKKK受有丝分裂原刺激磷酸化而激活,在此基础上MAPKKK转而磷酸化激活MAPKK,最后由MAPKK磷酸化MAPK,使其活化,完成信号放大和传递。
这是一种从酵母到人类都保守的三级激酶模式,信号最终被传递到细胞核,调节转录因子和相关酶的活性,参与细胞增殖、分化、转化及凋亡的调节,并与炎症、肿瘤等多种疾病的发生密切相关。
MAPK家族的信号通路主要包括细胞外信号调控的蛋白激酶(ERK)、c- Jun N端激酶(JNK)、P38MAPK以及ERK5四条途径。
ERK、JNK、P38、ERK5可以由不同的信号激活,形成不同的转导通路,激活各不相同的转录因子,介导不同的生物学效应,但这几条通路存在广泛的“cross talk”,从而导致通路间产生相互协同或抑制作用。
如在骨关节炎软骨损伤病症中,JNK、P38、ERK5协同参与细胞内的信号转导,介导软骨细胞的增殖和肥大分化,导致软骨钙化和骨赘的形成,最终形成骨关节炎。
从上个世纪开始,对细胞内信号转导通路的研究就已被广泛关注,有关细胞信号转导研究的技术和方法也是层出不穷,如免疫沉淀,RNA干扰,蛋白质组学,ELISA等,都可以用来检测信号转导过程中差异表达的信号分子以及关键蛋白的磷酸化。
Cloud-Clone Corp.已研发了针对MAPK级联激活信号通路的各种蛋白抗体,且研发出多种MAPK家族蛋白的ELISA 试剂盒供广大研究者选择使用。
细胞生物综述MAPK/ERK信号转导通路与学习记忆丝裂素活化蛋白激酶(MAPK)是细胞内的一类丝氨酸/苏氨酸蛋白激酶,是多种细胞外信号从细胞表面传导到细胞内的重要传递者。
细胞外信号调节激酶(ERK)包括ERKl和 ERK2,又称p44 MAPK和p42 MAPK,相对分子质量分别为4400和4200,是MAPK家族中的重要成员。
ERK最初被认为和细胞的生长、发育、分化有关,然而在已经不需要增殖和分化的成熟神经元中发现仍然有大量ERK上游调控子和下游靶蛋白存在,因此人们开始探索ERK的其他作用。
1997年English和Sweatt首次发现了ERK和长时程增强(LTP) 效应的关系,他们采用ERK上游激酶的抑制剂PD 098059阻止了ERK 的激活,同时也显著阻止了海马CA1区LTP 的诱导。
该发现随后又被其他研究小组证实并扩展,从而确定ERK与LTP的关系。
大量研究表明,LTP和学习、记忆过程密切相关,被许多学者命名为“学习、记忆的突触模型”,因此确定了ERK与学习、记忆的关系。
1 MAPK/ERK三级级联反应及与学习记忆的关系MAPK/ERK信号转导通路采用高度保守的三级激酶级联传递信号即Raf/MEK/ERK,活化的Raf使MEKⅧ亚区的两个丝氨酸磷酸化而激活,MEK 激活后使 ERK的苏氨酸和酪氨酸双位点磷酸化而激活。
研究表明,经水迷宫训练后的大鼠,背侧海马CA1/CA2区ERK被激活,使用PD098059抑制背侧海马 MAPK/ERK级联反应则磷酸化ERK蛋白(即活性形式)含量降低,并且长期空间记忆的形成受损。
海马内注入PD098059的大鼠精细定位信息受损。
利用转基因方法使小鼠前脑MEK1突变而突变的MEK1仍然能够和ERK相互作用,但却不能使它激活,这些小鼠表现为空间参考记忆及长期关联性记忆受损。
另一研究小组发现,利用转基因方法使小鼠脑中的MEK1突变,则这些小鼠海马区ERK活性降低,同时表现出特殊的恐惧性条件反射损害,此为海马依赖性的学习形式之一。