果蝇胚轴发育的机制
- 格式:docx
- 大小:17.03 KB
- 文档页数:4


果蝇胚轴发育的机制一、果蝇胚胎的极性果蝇早期胚轴形成涉及一个由母性影响基因产物构成的位置信息网络。
在这个网络中, 一定浓度的特异性母源性RNA 和蛋白质沿前- 后轴和背- 腹轴的不同区域分布, 以激活胚胎基因组的程序。
有4 组母性影响基因与果蝇胚轴形成有关, 其中3 组与胚胎前- 后轴的决定有关, 即前端系统( anterior system) 决定头胸部分节的区域, 后端系统( posterior system) 决定分节的腹部, 末端系统( terminal system) 决定胚胎两端不分节的原头区和尾节, 另一组基因决定胚胎的背- 腹轴, 即背腹系统( dorsovent ral system)。
控制发育的三类基因:1、母体效应基因2、影响身体分节的基因3、影响体节一致性的基因----同源异形框基因。
缺口基因表达区带宽大约相当于3个体节,其翻译的蛋白质以浓度效应调控成对控制基因(pair—rule genes)的表达。
成对控制基因为与前—后轴垂直的7条表达带,其翻译的蛋白质可激活体节极性基因(segment polarity genes)的转录。
体节极性基因的表达产物再进一步将胚胎划分成为14个体节。
同源异型基因的表达产物决定每个体节的发育命运果蝇胚胎的极性2.果蝇前—后轴(A-P)的形成1)前端系统:对于调节胚胎前- 后轴的形成有4 个非常重要的形态发生素: BICOID ( BCD ) 和HUNCHBACK( HB)调节胚胎前端结构的形成,NANOS( NOS) 和CAUDAL ( CDL ) 调节胚胎后端结构的形成。
Bicoid是控制头胸发育的一个关键母体效应基因,其不同浓度开启不同合子基因的表达。
在未受精卵中,bicoid mRNA定位在胞质前端;其受精后翻译出的蛋白质沿AP轴扩散,形成浓度梯度,为胚胎的后续分化提供位置信息。
bcd基因编码的BCD蛋白是一种转录调节因子,可与DNA 特异性结合并激活合子靶基因的表达。




果蝇的发育过程及其调控机制果蝇作为模式生物,其发育过程及调控机制已经被广泛研究。
果蝇的发育过程主要包括卵母形成、卵的受精、胚胎发育、幼虫期和蛹期等阶段。
这些阶段都受到不同的调控机制影响。
一、卵母形成和受精卵母形成过程在果蝇体内发生,一般从卵巢的端部开始,向基部发展。
在卵母形成早期,睾丸激素和卵泡刺激素对生殖细胞的发育起着重要作用。
随着卵母不断生长发育,卵母细胞核的复制和分裂也不断进行,最终形成完整的卵母。
成熟的卵母受到雄性精子的受精作用,形成受精卵。
在受精卵中,父本和母本细胞贡献的基因发挥不同的作用,影响胚胎的发育命运和表型表达。
二、胚胎发育受精卵受到一系列的调控因素作用,经过不同的胚胎发育阶段,最终形成成熟的幼虫体。
胚胎发育过程受到多个信号通路和基因网络的调控。
在果蝇的胚胎发育中,Wnt信号通路通过不同的基因调节细胞分裂和定向移动,对胚胎的头尾轴和背腹轴形成起着重要作用。
Hedgehog信号通路则调节胚胎前期的背腹轴形成。
同时,一个非常重要的因子是胰岛素样生长因子信号通路,其对幼虫的大小和发育有关键调节作用。
胚胎发育过程中也涉及到了一些胚胎基因调控系统,如TGFβ/Activin和Notch信号通路,调节着胚胎的细胞命运和分化。
三、幼虫期和蛹期在幼虫期和蛹期,果蝇表现出不同的形态和功能,同时也涉及到不同的代谢途径和调控机制。
在幼虫期,果蝇需要做出重要的生长和分化决策。
通过对食物和营养的感知和代谢途径的调控,果蝇可以根据生长的需要积累能量和物质,维持其生命活动的正常进行。
例如调节能量代谢的AMPK和Sirt1,以及调节食欲和代谢途径的Insulin/IGF1和TOR信号通路,都对幼虫的生长分化起着重要作用。
蛹期是果蝇生命周期中的一个转折点,幼虫期积累的营养和物质被重新配置为成虫身体的各种组织和器官。
在成虫器官分化和重构的过程中,也涉及到不同的调控机制。
例如调节上皮细胞形成和器官增殖的Notch信号通路,调节生殖腺发育的JAK/STAT信号通路,以及影响翅膀和大脑发育的Wnt和Hedgehog信号通路,都对蛹期的发育起着至关重要的作用。

果蝇胚胎形体模式构建的分子机制细胞发育的机制在不同物种中是不同的,但可以用果蝇作为模型来推论发育的基本原理,以期帮助人们了解其它生物发育的机制,即级联调节决定了胚胎细胞中基因表达的模式直至发育成为成体。
另外在不同种的相关生物中同源基因在发育中发挥的作用也是相关的。
虽然果蝇和哺乳动物的胚胎结构是不同的,但调控的方式相同。
和发育调控有关的基因是通过突变加以鉴别的。
有的突变导致在早期发育中致死,有的导致产生异常的结构。
影响到身体特殊部位发育的突变引起人们的注意,因为身体某部分的结构是复杂的,需要包括很多基因在内的一套特殊基因的表达。
一个基因的改变引起了整个身体某部分的结构的变化,因而可以推测这个基因可能是一个调控基因,担负着发育途经之间的“开关”或选择的作用。
在果蝇中体节是作为身体的一部分来进行分析的,在成蝇中能找到它的相应部分。
突变可以分为三组,通过它们对分节结构的作用来划分:(1)母体基因(maternal genes):通过母体在卵母细胞中表达,它们可能对成熟的卵母细胞起作用或者在早期胚中起作用;(2)分节基因(segmentation genes):在受精后表达,这些基因的突变会改变体节的数目或极性。
分节基因同分为3组,连续作用胚胎的较小区域;(3)同源异形基因(homeotic genes):控制体节的特征,而对其数目,大小和极性不起作用。
这些基因的突变会导致身体的一部分发育成另一部分的表型。
每一组基因的作用是使胚胎特异部分的特点逐步连续地显现出来。
母体基因作用卵中的广泛区域,其产物的分布差异控制了分节基因的表达;分节基因分划体节界线,同时同源异形基因控制体节的特征的发育。
首先起作用的是母体效应基因,这些基因决定胚胎的体轴,划分出胚胎形体模式或基因表达模式的总体格局。
接着表达的是负责身体分节的基因,这些基因在体轴确定的格局基础上,进一步将胚体划分为更细的基因表达区域。
再后表达的是同源异型基因,这些基因决定每一个体节的发育特征。

果蝇体轴形成的分子机制果蝇的卵、胚胎、幼虫、成虫都有明确的前后和背腹轴,在果蝇最初的发育中,由母源效应基因及其编码蛋白构成位置信息的基本网络,激活合子基因的表达,控制果蝇躯体模式的建立。
1、果蝇胚胎的极性果蝇早期胚轴形成设计由母源效应产物构成的位置信息网络,其中有3组与前-后轴形成有关,为前端系统;1组决定胚胎的背腹轴,即后端系统;还有一组决定背腹轴形成。
2、果蝇前-后轴的形成在果蝇前-后轴的形成中,有四个非常重要的形态发生素:Biociod(BCD)和Hunchback(HB)调节胚胎前端结构的形成;Nanos(NOS)和Candal(CDL)调节胚胎后端结构的形成。
1)前端组织中心: BCD蛋白浓度梯度前端系统至少包括4个主要的基因,其中起关键作用的是BCD,bcd是一种母源效应基因,在卵子发生时,bcd mRNA于滋养细胞中转录,再转运至卵子中并定位于卵子前极。
受精后迅速翻译,BCD具有决定胚胎极性和组织空间图示的功能。
受精后BCD蛋白在前端积累并向后端弥散,形成从前向后稳定的浓度梯度,主要覆盖胚胎前2/3区域。
bcd 基因也是同源异型框基因,BCD蛋白是一种转录调节因子,可与DNA特异性结合并激活合子靶基因的表达。
BCD蛋白浓度梯度可以同时特意新启动不同基因的表达,从而将胚胎划分为不同的区域。
2)后端组织中心:Nanos蛋白和Candal蛋白浓度梯度后端系统在控制图式形成中起到作用与前端系统相似。
决定胚胎后端的最初信息也是母源效应基因转录产物,在卵子发生过程中,后端决定子Nanos(NOS)的mRNA在卵室前端的滋养细胞中转录,通过转运定位到卵子后极,在成熟卵中定位于生殖质。
后端系统是通过抑制转录因子HB的翻译起作用,该系统包括约10个基因,这些基因都是腹部图示形成所必须的基因。
NOS活性从后端向前弥散形成浓度梯度,在胚胎后部抑制hb mRNA的翻译。
HB蛋白的分布区域主要位于胚胎前半部分。

果蝇胚胎发育和体轴形成的分子机制果蝇是一种十分常见的昆虫,它的胚胎期相当短暂,只需要不到两天的时间就能完成发育。
在这个过程中,许多重要的分子和信号通路都在起作用,对于了解发育和形态建成的分子机制具有重要意义。
本文将着重讨论果蝇胚胎发育和体轴形成的分子机制。
一、胚胎发育的基本过程果蝇的生命周期很短,在一般的实验室条件下,一只果蝇的寿命约为两个月。
雌性果蝇通常会在食物和水的混合物中产卵,大约在24小时内就会孵化。
在这之后的几天中,果蝇的发育非常迅速,逐渐从卵到幼虫,然后到成虫。
而在这个发育过程中,果蝇胚胎发育是非常关键的一步。
胚胎发育的过程可以分为五个连续的阶段。
第一阶段为卵细胞核形成,包括以交配卵产生的核和父母亲体细胞内孤雌生殖卵产生的核。
第二阶段是卵剖面形成,包括老二系统等。
通过定期锯切进行紧密环绕,则表明胚胎发展正常。
长时间僵滞不进则可能会在胚胎发展的后期出现显著的异常。
第三阶段是胚胎心脏形成,包括神经脑、背板神经织物、翼的最初生长和原胚内的胚芽形成。
第四阶段胚胎形成,则大半是身体器官、肌肉、皮肤等的形成。
卵子在此时分化为头部,胸部和腹部。
第五阶段是成虫形成,也就是最后的阶段。
它涉及更广泛的组织分化,包括瓢虫和蜜蜂等其他昆虫几乎都会经历的阶段。
以上这些阶段综合起来,构成了果蝇胚胎发育过程中的基本过程。
接下来我们将要探究果蝇胚胎形态的建立和维持所涉及的分子机制。
二、体轴形成的分子机制在果蝇发育的过程当中,体轴形成似乎是整个过程中最显著的阶段之一。
而体轴形成涉及到许多基因和信号通路的调控。
1. Wnt信号通路Wnt信号通路是胚胎发育中非常重要的一个通路。
Wnt分子在体轴形成中具有特殊作用,因为它们能够影响Dorsal基因的表达。
Dorsal基因在果蝇胚胎发育中发挥着严格的调控作用,在形成体胚层时非常重要。
Wnt拮抗剂能够使得胚胎中Dorsal基因表达出现异常,从而导致体轴形成不良。
2. Hedgehog信号通路Hedgehog信号通路也是在胚胎发育中重要的信号通路。

生命科学学院学院20 -20 学年第 学期《 发育生物学 》课程论文 课程号:2522080任课教师 成绩 论文题目:(可指定题目,也可说明题目范围。
)果蝇的发育及其发育的影响因素论文要求:(对论文题目、内容、行文、字数等作出判分规定。
) 1. 论文题目:准确得体,简短精炼,醒目2. 摘要:文字简练,字数不超过正文的5%;关键词不少于三个,关键词之间用分号间隔3. 正文:内容充实,论据充分、可靠,论证有力,主题明确语言流畅,条理清晰,字数不少于3000字4.字体:摘要、关键词宋体5号字;题目黑体三号字;正文宋体四号字 10分 教师评语:教师签字:年 月 日果蝇的发育及其发育的影响因素摘要:果蝇(Drosophila melanogaster)由于具有容易饲养、产卵多、生命周期短以及具有粗大的多线染色体而便于进行基因定位等优点,已成为遗传学和发育生物学等研究领域中重要的模式生物之一。
本文主要介绍了果蝇的发育过程及樟树叶片挥发物,紫外线照射,酵母粉和甲醛对果蝇生长发育的影响。
Abstract : Drosophila melanogaster is easy feeding, spaen many , shorter lifecycle and has athick multi-line chromosome.It has become such an important model organisms of Genetics and Developmental Biology field. This article mainly introduced the growth process of Drosophila melanogaster and some influence factor, such as essential oil produced by the leaves of camphor, ultraviolet rays, yeast powder and Formaldehyde. 关键词:果蝇;发育;樟树;紫外线;酵母粉;甲醛一、果蝇简介果蝇易于培养,生活周期短,基因组也比较小(1.4×108bp/每个单倍体基因组),只有4对染色体,且存在多线染色体,有关果蝇的遗传学知识已积累了80多年,遗传背景十分清楚,这些都是其重要的优势,故人们选择果蝇作为研究发育的模式动物。
果蝇胚胎发育过程中基因表达和细胞信号调节机制的研究果蝇是生物学史上最重要的模式生物之一,因为它们的基因组很小,很容易进行基因操作和研究,其生命周期短而且容易培养。
果蝇的生长和发育从受精卵开始到成熟昆虫,经历了一系列精细的调节和分化过程。
这些过程是由一系列基因的调控控制的,这些基因的表达和互作形成了非常复杂的基因调控网络。
这篇文章将探讨果蝇胚胎发育过程中的基因表达和细胞信号调控机制的研究。
果蝇受精卵发育最初的几个小时是决定其后的发育命运的关键时期。
在此期间,胚胎基因表达开始启动。
最早起始的是母系基因的表达,它们储存在成熟卵中并在胚胎发育过程中释放。
这些母系基因编码了一些重要的调控因子,如转录因子和剪接因子。
它们的表达为后续基因的调节控制奠定了基础。
在此之后,胚胎中出现了一些细胞信号分子,如趋化因子和生长因子。
这些信号分子在胚胎中施加影响,以确保细胞正确分化并形成特定的种类。
在果蝇胚胎中,基因调控网络的核心是由一系列转录因子组成的。
这些转录因子可以被分为两个主要类别:胚胎轴形成和胚胎分化因子。
胚胎轴形成因子如酸素和Toll等负责胚胎轴的定向和形成。
胚胎分化因子如Zerknüllt和Hunchback则负责细胞分化成特定的组织和器官。
根据基因调控网络的控制规律,胚胎轴形成和胚胎分化因子是相互影响的。
其中,胚胎轴形成因子优先出现并在分化之前确定了轴的方向和定向。
胚胎分化因子随着时间的推移,相继出现并开始对不同的组织和器官进行细胞分化。
大量的实验和基因编辑技术对果蝇胚胎发育的控制网络进行了深入的探索。
这包括详细的基因调控图谱,基因信号通路的鉴定和功能的研究。
现代技术大大提高了基因调控网络研究的速度和深度。
例如,近年来的高通量测序技术已经实现了对整个果蝇基因表达谱的鉴定和测定。
这种技术的应用使得我们对果蝇生长和分化的了解更加精确和全面。
细胞信号通路是基因调控网络的另一个核心组成部分。
通过这些信号通路,胚胎中的细胞可以相互作用并在发育和分化中进行调节。
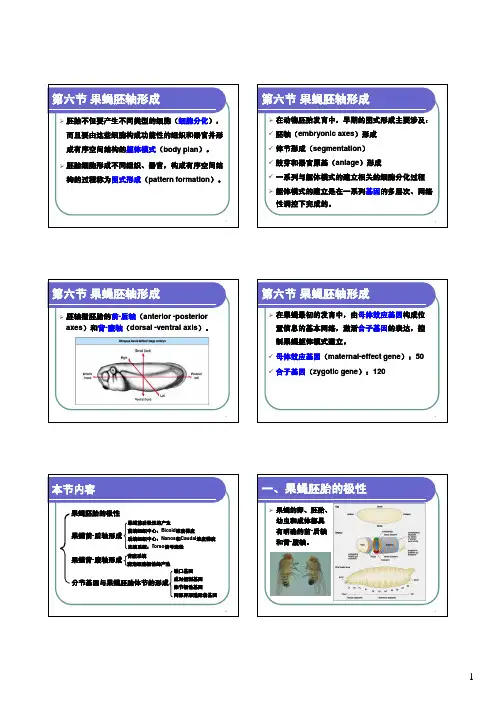
果蝇胚轴发育的机制一、果蝇胚胎的极性果蝇早期胚轴形成涉及一个由母性影响基因产物构成的位置信息网络。
在这个网络中, 一定浓度的特异性母源性RNA 和蛋白质沿前- 后轴和背- 腹轴的不同区域分布, 以激活胚胎基因组的程序。
有4 组母性影响基因与果蝇胚轴形成有关, 其中3 组与胚胎前- 后轴的决定有关, 即前端系统( anterior system) 决定头胸部分节的区域, 后端系统( posterior system) 决定分节的腹部, 末端系统( terminal system) 决定胚胎两端不分节的原头区和尾节, 另一组基因决定胚胎的背- 腹轴, 即背腹系统( dorsovent ral system)。
控制发育的三类基因:1、母体效应基因2、影响身体分节的基因3、影响体节一致性的基因----同源异形框基因。
缺口基因表达区带宽大约相当于3个体节,其翻译的蛋白质以浓度效应调控成对控制基因(pair—rule genes)的表达。
成对控制基因为与前—后轴垂直的7条表达带,其翻译的蛋白质可激活体节极性基因(segment polarity genes)的转录。
体节极性基因的表达产物再进一步将胚胎划分成为14个体节。
同源异型基因的表达产物决定每个体节的发育命运果蝇胚胎的极性2.果蝇前—后轴(A-P)的形成1)前端系统:对于调节胚胎前- 后轴的形成有4 个非常重要的形态发生素: BICOID ( BCD ) 和HUNCHBACK( HB)调节胚胎前端结构的形成,NANOS( NOS) 和CAUDAL ( CDL ) 调节胚胎后端结构的形成。
Bicoid是控制头胸发育的一个关键母体效应基因,其不同浓度开启不同合子基因的表达。
在未受精卵中,bicoid mRNA定位在胞质前端;其受精后翻译出的蛋白质沿AP轴扩散,形成浓度梯度,为胚胎的后续分化提供位置信息。
bcd基因编码的BCD蛋白是一种转录调节因子,可与DNA 特异性结合并激活合子靶基因的表达。
缺口基因hb 是其靶基因之一,是控制胚胎胸部及头部部分结构发育的重要基因。
3)后端组织中心:Nanos蛋白和Caudal蛋白浓度梯度决定胚胎后端图式的最初信息也是母体效应基因转录产物,在卵子发生过程中,nos mRNA在卵室前端的滋养细胞中转录,通过转运定位到卵子后极。
后端系统:mRNA的产物不能直接调节合子基因的表达,而是通过抑制一种转录因子的翻译来进行调节。
后端系统包括约10个基因,这些基因的突变都会导致胚胎腹部的缺失。
在这一系统中起核心作用的是nanos(nos) 基因。
其编码产物NOS 活性从后向前弥散形成浓度梯度。
NOS 的功能是在胚胎后端区域抑制母源性hb mRNA的翻译。
Nanos和Caudal蛋白梯度控制后区结构nanos ,Nanos决定后部区的发育,它在受精后形成P-A浓度梯度,其作用是与hunchback mRNA结合,阻止后者在后区的翻译,帮助形成Hunchback蛋白梯度。
Hunchback:母体mRNA在卵中均匀分布,受精后前区高浓度的 Bicoid蛋白激活hunchback基因的表达,Nanos控制hunchback mRNA的翻译从而帮助形成hunchback蛋白浓度梯度。
Caudal: 母体mRNA在卵中均匀分布,受精后bicoid蛋白抑制其在前区的表达,因而Caudal蛋白形成类似于nanos的浓度梯度。
末端系统:Torso信号途径卵膜表面受体的激活决定胚胎AP 轴的两个端点? Torso缺失突变体缺少原头区和尾区,torso蛋白为酪氨酸激酶类受体。
未受精前,torso已均匀地分布在卵的质膜上。
但其配体torsolike定位在两端的卵外膜(vitelline membrane) 上,不能与torso结合。
受精时,torsolike得以释放,torsolike与torso 结合, torso活化,启动信号传导。
Torsolike蛋白的存在量很低,受精后其扩散距离有限。
其突变体类似torso突变体分节基因与果蝇胚胎体节的形成分节基因的功能是把早期胚胎沿前—后轴分为一系列重复的体节原基3.背腹轴的形成背–腹系统的作用方式与末端系统有相似之处。
通过一种局部分布的信号分子,即定位于卵子腹侧卵黄膜上的配体激活分布于腹侧卵黄膜上的受体,进而调节下游合子基因的表达。
背-腹系统对合子靶基因表达的调节方式前端系统相似,通过一种转录因子的浓度前端系统相似,通过一种转录因子的浓度梯度来完成。
但背腹系统浓度梯度形成的方式却与前端系统完全不同,dl基因是这一信号传导途径的最后一个环节,它编码一种转录调节因子。
dl mRNA和DL蛋白在卵子中是均匀分子。
dl mRNA和DL蛋白在卵子中是均匀分布。
当胚胎发育到第9次细胞核分裂之后,细胞核迁移到达合胞体胚盘的外周皮质层,在腹侧的DL蛋白开始往核内聚集,但背侧的DL蛋白仍位于胞质中。
从而使DL蛋白在细胞核内的分布沿背腹轴形成一种浓度梯度。
1)DL蛋白定位于细胞核中的机制:cactus基因与DL蛋白能否进入细胞核这一调控过程有关。
CACTUS与DL结合时,DL蛋白不能进入细胞核。
toll基因在这一系统中具有极其重要的作用。
TOLL是一种跨膜受体蛋白,其配体分子也是母源性产物,是spǟtzle基因编码蛋白的裂是母源性产物,是spǟtzle基因编码蛋白的裂解片段。
Spǟtzle蛋白由卵室腹侧的特异性滤泡细胞产生,在胚胎发育的早期被释放定位于卵周隙中。
Spǟtzle蛋白与DL受体结合并使之活化,进而激发一系列细胞内信号传导,最终使CACTUS 蛋白降解,DL蛋白释放进而进入细胞核。
DL蛋白的浓度梯度通过对下游靶基因的调控,控制沿背-腹轴产生区域特异性的位置信息。
这种浓度梯度在腹侧组织中可活化信息。
这种浓度梯度在腹侧组织中可活化合子基因twist twi)和snail (sna)的表达,同时抑制dpp和zen基因的表达,进而指导腹部结构的发育。
dpp和zen基因在胚胎背侧表部结构的发育。
Toll蛋白的活化导致沿背腹轴方向细胞核之间dorsal蛋白梯度的形成。
果蝇核蛋白dorsal沿背腹轴的梯度将身体分为不同部分的模型。
twist和dpp等基因的激活解读dorsal蛋白的浓度梯度;Dpp蛋白的活性在sog蛋白的拮抗作用下局限于胚胎最背部的区域。
2)卵子发生过程中体轴的极化滋养细胞(滋养细胞(nurse cell)合成大量的蛋白和cell)合成大量的蛋白和mRNA通过胞质桥(mRNA通过胞质桥(cytoplasmic bridge)转bridge)转运至卵细胞中。
滤泡细胞在决定卵轴极性运至卵细胞中。
滤泡细胞在决定卵轴极性方面发挥这重要的作用。
果蝇卵子的发育果蝇卵子与15个滋养细胞相连,被一层滤泡细胞(700个左右)覆盖。
卵细胞和滤泡细胞协同作用确定将来卵子的背腹轴。
果蝇的卵细胞进入卵室(egg chamber)的后端,与后端的滤泡细胞建立联系。
但卵细胞与前端的滤泡细胞被滋养细胞隔开。
卵细胞中合成gurken mRNA,而gurken蛋白在局部分泌。
Gurken蛋白与后极滤泡细胞上表达的受体torpedo的结合引起相邻的滤泡细胞特化为后极滤泡细胞。
后极滤泡细胞发送信号至卵细胞,重排细胞骨架微管,从而将BCD和oskar蛋白分别定位于卵的前端和后端确定了卵的前后轴。
随后卵细胞的核运动到将来的背侧,随后卵细胞的核运动到将来的背侧,gurken蛋白的局部释放使相邻的滤泡细胞特化为背部的滤泡细胞,卵子将来的背方也得到确定。
3)信号调控在卵子发生过程中,滤泡细胞背腹极性的获得是由卵细胞的信号调控的。
这个过程与TOR途径有相似之处,但是信号传递的TOR途径有相似之处,但是信号传递的方式却相反。
这个信号传递途径至少包括7 个基因,这些基因的突变会影响卵壳和卵的极性。
grk、top或cni的突变产生腹部化的的极性。
grk、top或cni的突变产生腹部化的胚胎,而fs(1)k10、sqd、spir或capu的失活胚胎,而fs(1)k10、sqd、spir或capu的失活则产生背部化的卵和胚胎。
4.分节基因分节基因的突变可使胚胎缺失某些体节或体节的某些部分。
分节基因的可分为三类:缺口基因、成对控制基因和体节极性基因。
这三类基因的调控是逐级进行的,首先由母体效应基因控制缺口基因的活化,其次缺口基因之间互相调节彼此的转录并且共同调节成对控制基因的表达,然后成对控制基因之间相互作用把胚体分隔成为一系列重复的体节,并且进一步控制体节极性基因的表达。
缺口基因和成对控制基因再共同调控同源异型基因的表达。
1. 缺口基因缺口基因( gap gene)的表达区域为一些较宽的区域, 每个区域的宽度约相当于3个体节, 表达区之间可有部分重叠。
当缺口基因突变时胚胎缺失相应的区域。
缺口基因直接受母体效应基因的调控。
2. 成对控制基因成对控制基因( pair - rule gene )的表达区域以两个体节为单位且具有周期性, 在相互间隔的一个副体节中表达。
这些基因的功能是把缺口基因确定的区域进一步分成体节。
当这类基因突变时相应的表达区域缺失。
成对控制基因的表达是胚胎出现分节的最早标志。
它们在细胞化胚盘期第13 次核分裂时开始表达。
表达图式非常独特, 沿前- 后轴形成一系列斑马纹状的条带, 正好把胚胎分为预定体节。
3. 体节极性基因体节极性基因( segment polarity gene )在每一体节的特定区域细胞中表达。
当这类基因突变时每个体节都缺失一个特定的区域。
4. 同源异型选择者基因( homeotic selector gene)(1 ) 同源异型选择者基因的表达图式在体节界限确立之后每个体节的结构被进一步特化, 此过程由主调节基因(master regulatorygene )或称为同源异型选择者基因调控完成。
果蝇大部分同源异型选择者基因都位于第 3 号染色体相邻的两个区域, 其中一区域称为触角足复合体( antennapedia complex , Antp - C) , 另一个区域是双胸复合体( bithorax complex , BX - C) , 这两个复合体统称同源异形复合体( homeotic gene complex, HOM - C)。
(2 ) 同源异型选择者基因表达图式的建立与维持同源异型选择者基因表达图式的建立受成对控制基因和gap 基因的调控。