2011CB944000-G哺乳动物后肾发育重要环节及关键调控因子的研究
- 格式:doc
- 大小:488.50 KB
- 文档页数:25

哺乳动物生殖系统的分子调控机制研究哺乳动物生殖系统的正常发育和功能受到复杂的分子调控机制的影响,包括激素、细胞因子、信号转导通路等多种因素。
这些因素参与在精子和卵子生成、排出和受精过程中,以及胚胎着床、胚胎早期发育、性成熟以及生殖周期等各个方面。
本文将重点介绍几种重要的信号通路和关键分子,在哺乳动物生殖系统中发挥重要作用的机理和研究进展。
激素调控激素是哺乳动物生殖系统中重要的调控因素,包括促性腺激素(GnRH)、促卵泡素(FSH)、黄体生成素(LH)等,在雌性和雄性生殖系统中均起到关键作用。
GnRH 是促性腺激素释放激素,是垂体前叶释放促性腺激素的重要调节因子。
GnRH 受体(GnRHR)是局部调节促性激素合成和释放的重要分子,它的表达水平与生殖周期、排卵和受孕机会等密切相关。
其中,在雌性动物中存在两种GnRHR亚型(GnRHR1 和 GnRHR2),并在卵巢和子宫等组织中表达。
目前的研究发现,GnRHR 参与了雌性生殖系统中多种重要生理过程,如卵泡发育、卵巢周期与黄体形成等。
而在雄性生殖系统中,GnRHR 在精子发生中也发挥作用。
除此之外,FSH 和 LH 是垂体前叶的两种重要的促性腺激素。
它们被释放后可以刺激卵巢和睾丸的细胞分化和成熟,从而参与生殖细胞生成和调控。
这些激素通过 G 蛋白偶联的受体介导信号传导,激活下游分子的转录因子,这些传递下来的信号通路为了研究生殖细胞发育有着重要意义。
信号转导在激素作用的基础上,细胞内外的信号传导过程也对哺乳动物生殖发育起到至关重要的作用。
其中,细胞膜上的受体包括 G 型蛋白偶联受体和酪氨酸激酶受体等,如上述的GnRHR 就属于前者。
G 型蛋白偶联受体活化后,由GTP 与其结合,从而激活腺苷酸酶(AC)、磷脂酰肌醇(PI)3-激酶、蛋白激酶C(PKC)等信号分子,最终促进生殖细胞发育。
而另一类受体-酪氨酸激酶受体,则能够启动一条tyrosine kinase(TK)的信号通路。
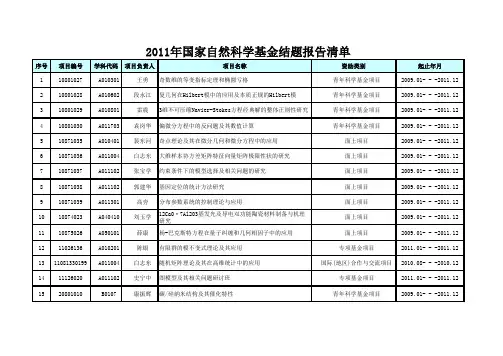
序号项目编号学科代码项目负责人项目名称资助类别起止年月110801027 A010301 王勇奇数维的等变指标定理和椭圆亏格 青年科学基金项目2009.01- - -2011.12 210801028 A010602 段永江复几何在Hilbert模中的应用及本质正规的Hilbert模 青年科学基金项目2009.01- - -2011.12 310801029 A010801 雷震3维不可压缩Navier-Stokes方程经典解的整体正则性研究 青年科学基金项目2009.01- - -2011.12 410801030 A011703 袁岗华偏微分方程中的反问题及其数值计算 青年科学基金项目2009.01- - -2011.12 510871035 A010401 裴东河奇点理论及其在微分几何和微分方程中的应用 面上项目2009.01- - -2011.12 610871036 A011004 白志东大维样本协方差矩阵特征向量矩阵极限性状的研究 面上项目2009.01- - -2011.12 710871037 A011102 张宝学约束条件下的模型选择及相关问题的研究 面上项目2009.01- - -2011.12 810871038 A011102 郭建华基因定位的统计方法研究 面上项目2009.01- - -2011.12 910871039 A011301 高夯分布参数系统的控制理论与应用 面上项目2009.01- - -2011.121010874023 A040410 刘玉学12CaO·7Al2O3基发光及导电双功能陶瓷材料制备与机理研究面上项目2009.01- - -2011.121110875026 A050101 薛康杨-巴克斯特方程在量子纠缠和几何相因子中的应用 面上项目2009.01- - -2011.12 1211026136 A010201 陈银有限群的模不变式理论及其应用 专项基金项目2011.01- - -2011.12 1311081330199 A011004 白志东随机矩阵理论及其在高维统计中的应用 国际(地区)合作与交流项目2010.08- - -2010.12 1411126020 A011102 史宁中图模型及其相关问题研讨班 专项基金项目2011.01- - -2011.12 1520801010 B0107 康振辉碳/硅纳米结构及其催化特性 青年科学基金项目2009.01- - -2011.122011年国家自然科学基金结题报告清单序号项目编号学科代码项目负责人项目名称资助类别起止年月2011年国家自然科学基金结题报告清单1620803008 B030702 刘阳ZnO的一维纳米材料及量子点:合成、组装及光化学性质研究青年科学基金项目2009.01- - -2011.121720804008 B0403 孙海珠环境友好纳米粒子表面接枝共轭聚合物的复合材料光伏性质研究青年科学基金项目2009.01- - -2011.121820805005 B050104 杨丽光快门漂白进样-电泳媒介微分析法连用的毛细管电泳在线酶反应研究平台青年科学基金项目2009.01- - -2011.121920871026 B01 王晓红同时具有B酸和L酸双重酸中心固体杂多酸催化剂的合成及催化性能研究面上项目2009.01- - -2011.122020871027 B010204 刘术侠HDACi多酸前导化合物的设计合成和抗肿瘤活性 面上项目2009.01- - -2011.122120872015 B0201 刘群基于多官能团化插烯式硫代羧酸衍生物的多取代异喹啉和萘啶的多样性合成面上项目2009.01- - -2011.122220873017 B030201 仇永清开壳层有机/配合物分子NLO性质的理论研究 面上项目2009.01- - -2011.12 2320873018 B0303 郭伊荇固体多酸非均相光催化反应机理研究 面上项目2009.01- - -2011.12 2420875012 B050206 郭黎平基于有序介孔碳及其功能化材料的电化学传感平台研究 面上项目2009.01- - -2011.12 2520877017 B070403 徐镜波硝基甲苯暴露鲤鱼的基因表达及能量分配 面上项目2009.01- - -2011.12 2630800119 C030102 唐占辉西双版纳地区果蝠与植物的协同进化 青年科学基金项目2009.01- - -2011.12 2730800557 C0703 王秀莉精氨酸甲基化对p16功能的影响及其分子机制研究 青年科学基金项目2009.01- - -2011.122830870178 C020303 刘宝松嫩草原三种重要牧草--短芒野大麦、芦苇和羊草天然种群的表观遗传多样性及其与逆境适应的可能关系面上项目2009.01- - -2011.122930870222 C020409 朱长甫玉米类胡萝卜素生物合成基因的克隆与功能分析 面上项目2009.01- - -2011.12 3030870238 C020601 郭继勋松嫩草地C3、C4植物光合特性对CO2和温度升高的响应 面上项目2009.01- - -2011.12序号项目编号学科代码项目负责人项目名称资助类别起止年月2011年国家自然科学基金结题报告清单3130870318 C0404 高玮两种洞巢鸟巢址选择中对公共信息的利用 面上项目2009.01- - -2011.12 3230870371 C0302 冯江同域共存的马铁菊头蝠和大趾鼠耳蝠生境选择机制 面上项目2009.01- - -2011.123330871301 C0709 李晓萌hZimp10对Smad3/4复合物转录活性的调控在前列腺癌细胞凋亡中的作用机制研究面上项目2009.01- - -2011.123430871302 C0709 朱筱娟肌球蛋白Ⅹ参与侵入性伪足形成并介导肿瘤细胞侵袭的研究面上项目2009.01- - -2011.123530872365 H1005 王丽白色念珠菌天冬氨酸蛋白酶表位的噬菌体展示及其产物的免疫学研究面上项目2009.01- - -2011.123640871005 D0101 刘惠清城市沟谷地污染元素迁移的机理研究 面上项目2009.01- - -2011.123740871120 D010504 殷秀琴松嫩草地土壤动物多样性及其在凋落物分解中作用和物质能量收支研究面上项目2009.01- - -2011.123840871236 D011002 张继权基于多源信息融合的草原火灾风险评价体系构建及其在应急管理中的应用研究面上项目2009.01- - -2011.123950725205 E020702 刘益春发光及显示材料 国家杰出青年科学基金2008.01- - -2011.124050802014 E0207 张昕彤p-型CuI/窄带隙半导体薄层/n-型氧化物纳米线异质结固态太阳电池材料与器件青年科学基金项目2009.01- - -2011.124150873019 E030901 王广萘酚吡喃聚合物光致变色材料的设计和性能 面上项目2009.01- - -2011.124250873020 E030901 张景萍宽光谱强吸收有机太阳能电池给体材料的分子设计与性能研究面上项目2009.01- - -2011.124350878041 E080402 袁星多酸-二氧化钛复合膜的制备及其光催化降解水中有机污染物研究面上项目2009.01- - -2011.124460803102 F020509 殷明浩开放式规划研究 青年科学基金项目2009.01- - -2011.12 45A0724101 郭建华现代统计暑期讲习班 专项基金项目2008.01- - -2008.12。


维生素C和E与哺乳动物生殖的关系生理科学进展2005年第36卷第1期维生素C和E与哺乳动物生殖的关系木陶勇张美佳(中国农业大学生物学院,北京100094;谢辉蓉夏国良安徽农业大学畜牧水产学院,合肥230036)摘要维生素在哺乳动物各个系统的生长发育中和正常的生理功能发挥过程中起着举足轻重的作用,具有调节代谢,调节渗透压,调节免疫力,维持酸碱平衡等功能.同样,维生素对哺乳动物的生殖也具有非常重要的作用,尤其是维生素E(生育酚)和维生素C(抗坏血酸),在抗氧化,抗自由基,抗不育,抗衰老,抗癌变等过程中发挥重要作用.关键词维生素E;维生素C;卵母细胞;胚胎;生殖中图分类号Q954.4维生素是哺乳动物维持正常代谢所必需的一类低分子的有机营养物,维生素在动物体内含量很少,大多数维生素必须从食物或饲料中取得.维生素在生理上不是构成各种组织的主要原料,也不是体内能量的来源,但是却有很重要的生理功能,是许多酶的辅酶或辅基的组成成分,起着调节免疫力和控制代谢的作用,还具有抗氧化,抗自由基,抗凋亡和抗衰老等多方面的功能.维生素对生殖性能的影响也十分明显,几乎所有的维生素都与哺乳动物的生殖行为密切相关.在几十种维生素中,维生素c和E与生殖系统的作用相对更加明显和直接.维生素c又称抗坏血酸,溶于水,化学结构中有2个具有氧化和还原功能的烯二醇基团,既可参与氧化反应,又不失还原的作用,大多是辅酶的组成部分,在人体内参与许多重要的代谢或反应,它是细胞外最重要的抗氧化剂,在清除自由基方面发挥重要的作用¨.维生素E又称生育酚,是一组具有生物活性的化学结构相似的酚类化合物,天然的维生素E有8种.维生素E为浅黄色,为粘性油状物,几乎无臭,不溶于水,可溶于醇和油脂,遇光色泽变深,易被氧化,故在体内可保护其它可被氧化的物质,如不饱和脂肪酸. 维生素E具有捕捉自由基的作用,是哺乳动物细胞膜上清除自由基的首要因子.这两种维生素与哺乳动物的生殖系统的功能活动密切相关,缺乏就会引发多种机能障碍,生殖能力减退甚至丧失.一,维生素C在哺乳动物生殖中的作用Chinoy等(1986)就发现,睾丸和精液对抗坏血酸的浓度下降极其敏感.Jacob等(1992)发现,精液中抗坏血酸的浓度是血液中的8~l0倍,提示它可能具有重要的生化功能.Dawson等又发现,不育男性的精液中抗坏血酸的水平果然显着低于生育健康男性.维生素c是生殖过程中不可或缺的生化因子',通过在食物中增加维生素c,能够提高25岁以上男性的精子的存活力和运动力,增加成熟精子数,降低精子粘附性,减少精子畸形率..对于维生素c影响雄性生殖性能的原因,Fraga等认为,精液中维生素c可能对精子的DNA具有保护作用.精液中高浓度的维生素c可使精子内的重要成分免遭氧自由基的损伤,保护精子免受有害物质引起的基因损伤,这种损伤极可能使其子女罹患可怕的遗传病或癌症.如果减少维生素c的摄人量,男性精子中的脱氧核糖核酸的损伤就会成倍增加.雌性动物血浆中维生素c的水平在受孕期问上升到最高,这提示维生素c在这一过程中的重要作用.大鼠卵巢排卵前卵泡中含有大量的抗坏血酸,研究表明,高水平的抗坏血酸能防止卵泡的凋亡.体外培养卵母细胞或胚胎时,一个重要的问题是高浓度的氧环境,这时会产生许多自由基,对卵母细胞和胚胎的发育不利.为此,可以通过在培养基中添加维生素c的方法对抗自由基的作用.维生素c为酸性物质,中性pH环境中性质不稳定,而卵母细胞适宜的生长pH范围是7.2~7.4,因此用人工合成的方法,在抗坏血酸的分子中连接糖苷,成了对热稳定的化合物AA一2G,更适用于培养卵母细胞或胚胎.Tatemoto等用AA一2G研究了维生素c对猪裸卯(cumulus—denudedoocytes,DOs)的体外培国家杰出青年基金(30025032)和北京市自然科学基金(6992019)资助课题通讯作者生理科学进展2005年第36卷第1期养成熟的作用,发现250p~mol/LAA一2G虽然对DOs 的成熟率,精子穿透率和多精子受精没有明显影响,但能够提高雄原核的形成率,并进一步提高囊胚发育率.Tao等用次黄嘌呤模拟机体的生理成熟状态,分别研究了维生素c对DOs和卵丘包被的卵母细胞(cumulus—enclosedoocytes,CEOs)体外成熟的作用,发现对CEOs卵丘扩展和减数分裂成熟没有明显影响,但能延缓卵丘细胞的凋亡;对于DOs,维生素c能促进生发泡破裂(germinalbreakdown, GVBD),进一步研究还发现,它还可促进DOs由第一次减数分裂中期(metaphaseI,MI)发育到第二次减数分裂中期(metaphaseII,MII),作用的最佳浓度仍是250p~mol/L.由于CEOs中卵匠细胞能通过升高卵母细胞的GSH水平来保护卵母细胞在体外培养时因环境高氧而产生的氧化刺激作用,而维生素C又可以提高DOs内谷胱甘肽(glutathione, GSH)的水平,降低了DNA的损伤,因此维生素c对于DOs可以部分替代卵丘细胞的功能,促进DOs 的细胞胞质成熟.现已找到转运维生素C的物质SVCT1和SVCT2,尤其是前者的亲和性很高,其基因SLC23A2 位于人第5号染色体上(5q31.2—31.3),SVCT1主要分布于肝脏和肾脏内,而SVCT2则广泛分布在大脑,视网膜,内分泌和神经内分泌器官内,这为研究健康或疾病状态下维生素c的代谢提供了方便. 二,维生素E在哺乳动物生殖中的作用在维生素E发现的最初,就知道它对动物的生殖和发育都有明显的影响.在饲料中给种公猪补允维生素E,能够提高公猪的精子浓度和一次射精的精子总数.雄鼠缺乏维生素E后,可导致永久性的不可逆的不育,雌鼠则导致流产.活性氧基(ROS)的产生和精子膜的过氧化会损伤精子的运动力,导致精子中段畸形和精卵融合障碍,维生素E能保护精子不受这种损伤.维生素E还能保护重金属对动物繁殖性能的损害作用,Rao和Sharma给雄性小鼠强行饲喂氯化汞,会导致附睾中精子减少,精子运动力和活力下降,但同时饲喂维生素E则能逆转这些变化,并使睾丸,附睾和输精管中的氯化汞下降Y ousef等通过饮水给公兔补充维生素C和E,发现这两种维生素能够显着增加射精量,精子浓度,精子总数,精子活力和果糖浓度,降低pH值,同时还显着降低畸形精子和死精子的比率,显着降低精液中天门冬氨酸转氨酶,丙氨酸转氨酶和乳酸脱氢酶的活性,而谷胱甘肽转移酶(GSH)的活性显着提高,尤其以维生素E的作用更为明显,他们还发现这两种维生素能够显着提高公兔的摄食量但体重增加不明显.Salem等发现维生素c能增加兔体重,但Shetaewi报道,维生素c和E能降低兔的采食量,增加体重,即提高了饲料的利用效率.关于维生素c能够提高精液品质的原因,许多报道都认为,是由于其降低了机体自由基的生成.维生素E与雌性哺乳动物的繁殖性能有着密切的关系,可刺激垂体前叶的功能活动,促进分泌细胞分泌促性腺激素增加,从而调节性腺的生长发育和功能活动,如促进精子的生成与活动,增强卵巢的生理机能,使卵泡增加和黄体细胞增大并使孕酮的作用增强,防止流产,提高产仔数,降低幼仔断奶前死亡率.连续流产的妇女,用激素治疗无效的不孕妇女,服用维生素E可获治愈,子宫内膜增厚,胚胎移植后妇女成功受孕.不过也有不同的报道,Tarin等给小鼠从断奶开始饲喂含维生素c和维生素E 的混合食物,至l0周龄,结果发现对受精率,囊胚率,囊胚细胞数,有丝分裂和核凋亡都没有显着影响,卵母细胞的MPF活性不变,而MAPK的活性降低,取出的卵母细胞经孤雌激活后,有更多的卵发育形成原核一.Gardner等也发现维生素是通过改变代谢酶的活性提高绵羊卵裂率和胚胎发育率. Gabryszuk和Klewiec研究发现,给2~3岁的母羊肌肉注射维生素E和硒酸钠,并不能提高母羊的繁殖性能(包括发情和产羔数).雌性哺乳动物缺乏维生素E会导致一系列生殖系统的病症,如不易受精,习惯性流产,不可逆的不育症,雌鼠缺乏维生素E后妊娠约在l0天内终止,公猪缺乏维生素E会引起睾丸退化,雄鼠长期缺乏维生素E会发生上皮变性.在禽类,维生素缺乏可引起繁殖机能紊乱,精子数量减少,种蛋孵化率低.体外培养卵母细胞和胚胎时,发现在培养体系中加入维生素E,会对卵母细胞和胚胎的发育产生影响.由于维生素E是脂溶性的,体外培养时用95%的乙醇配制成2000倍的浓贮液,培养前用稀释2000倍,培养基中乙醇的浓度为0.05%,不会对卯母细胞或胚胎产生影响.发现维生素E对CEOs的卵丘扩展和减数分裂成熟没有明显影响,但能延缓卵丘细胞的凋亡;对于DOs,维生素E能促进生发泡破裂,并促进DOs由MI期发育到MII期,作用的最佳浓度为10/~mol/L_6'.Tsujii等在体外培养小鼠胚胎时加入维生素E,能够增加胚胎对葡萄糖的利用,提高囊胚发育率.Olson和Seidel也报道了维生素E可促进牛胚胎体外发育.维生素E能有效保护大鼠的胚胎体外发育时免受氧化性刺激的损伤,保护小鼠体外培养的胚胎在热应激时仍能发育,并且还能提高胚胎的存活力.这些体外实验的结果提示维生素E可能在体内发挥相似的作用. 有趣的是,无论是维生素C或维生素E,单独使用都能促进胚胎发育,提高囊胚形成率,但同时使用却抑制牛胚胎的发育.三,维生素C和E的毒副作用饲料和食物是维生素C的最佳来源,例如人每天需要摄入60mg维生素C,只相当于一个桔子中的维生素C的含量,就可以保护男性精子免受损伤.膳食平衡的动物或人无需另行补充.吸烟会损伤男性的精子,从而影响生殖健康,需要增加维生素C的摄入.维生素C的补充相对安全,大量补充的不良反应的发生率较低.但这并不意味着就可以长期大剂量食用维生素C的药物.由于维生素C具有酸性,直接的刺激作用可使胃肠蠕动增加而导致头痛恶心,呕吐腹泻,胃痉挛,尿频尿痛和尿结石等症状,还有可能产生依赖性,减量时会发生回跳性坏血病.大剂量长期补充维生素C也会干扰凝血.维生素C还会影响维生素B的吸收和利用,与维生素K可发生氧化还原反应,因此不宜长期大量补充.此外,长期补充过量维生素C对生殖功能也会产生不良影响,但大多是间接的.口服避孕药的妇女,血浆中维生素C的含量亦低,如突然停用维生素C,可因相互作用引起破溃性出血而使避孕失败. 我们还发现,在猪卵母细胞体外成熟培养体系中加入维生素C,可不同程度地损伤DOs和CEOs的存活力[6Z.维生素E的需要随着饲料中其他成分的含量多少而有所变化,不饱和脂肪酸,蛋白质,胆固醇,维生素A和硒等成分的含量均会影响维生素E的摄入量.补充维生素E相对安全,多数动物摄入超过其日常供应量的100倍都不发生不良反应.但过量补充维生素E也会对动物的繁殖性能产生不良的影响.Tarin等给雌性小鼠饲喂大剂量的维生素E, 结果发现其生殖性能下降,产仔间隔延长,产仔数下降,断奶时存活雄性仔鼠减少;卵巢和子宫的功能受损,卵巢黄体数减少,左侧子宫角隐性流产胎儿数增加¨….过量摄取维生素E可能会抑制维生素A及维生素K的吸收,摄入量大于1200mg/d生育酚当量时,可干扰维生素K的代谢,降低血小板的粘附能力,增强某些抗凝剂的作用,引起术后出血.高剂生理科学进展2005年第36卷第1期量维生素E会引起仓鼠睾丸萎缩和精子产生量减少.胎儿体内高浓度的维生素E可能会抑制嗜中性粒细胞对细菌及真菌的杀伤能力,从而增加早产胎儿患脓毒血症的危险.虽然成人每日服用100~800mg,连续半年后进行20多项血液检查,未发现任何异常,但是,长期大量服用可引起恶心,呕吐,眩晕,头痛,视力模糊,皮肤皲裂,唇炎,口角炎,胃肠功能紊乱,腹泻,乳腺肿大或乏力软弱等症状,超量服用(每日超过800mg)可能引起凝血时间延长,内分泌代谢紊乱(甲状腺,垂体和肾上腺)和免疫机制的改变,妇女可引起月经过多,闭经,性功能紊乱等,并有引起血栓性静脉炎或栓塞的危险.此外,维生素E的代谢物具有拮抗维生素K的作用,能降低血液凝固性,维生素E与雌激素长期并用可诱发血栓性静脉炎.四,研究展望到目前为止,维生素对生殖健康以及配子/胚胎体外发育作用和影响的报道仍然不多,研究工作不够系统和完整,许多问题没有得到深入研究和解决.例如,有实验表明,在培养基中添加多种维生素能促进仓鼠胚胎发育,促进山羊卵母细胞发育能力;但究竟是哪一种维生素在起作用,或者是哪几种维生素协同作用?如果是协同作用,最佳的剂量关系如何?这些仍没有研究清楚.此外,尚缺乏对维生素作用机制的深入认识,这仍是今后很长一段时间内需要做的工作.随着对维生素认识的不断深入,维生素C和维生素E对哺乳动物的繁殖乃至人类的生殖健康和配子/胚胎体外发育,必将会作出更多的贡献.参考文献1RoseRC,BodeAM.Biologyoffreeradicalscavengers:ane,'aluationofascorbate.FASEBJ.1993.7:1135~1142.2ChowCK.VitaminEandoxidativestress.FreeRadicalBiol Med.1991.11:215~232.3DawsonEB,HalTiSW A.|reterMC,eta1.Efrectofaseorbie acidsupplementationonthespermqualityofsmokers.Fertil Steril.1992.58:1034~1039.4LuckMR,JeyaseelanI.ScholesRA.Minireview:ascorbic acidandfertility.BiolReprod,1995.52:262~266.5TatemotoH,OkudaT.SokoN.eta1.Malepronuelealfor. mationandblastocystformationaleimprovedbysupplemen—tationofAscorbicAeid2一O—d—Glucosideduringinvitromat—urationcultureofdenudedporcineooeytes.JReprodDev,2001.47:329~339.生理科学进展2005年第36卷第哺乳动物精子与卵子质膜粘附和融合的分子基础冯伯森闰一琳(辽宁师范大学生命科学学院,大连116029)摘要精子与卵子质膜粘附并发生融合是哺乳动物完成受精过程所必需的步骤.近年来,学者们以现代分子生物学理论为基础,对参与精卵质膜粘附,融合过程的分子进行了研究,特别是精子表面的去整合素金属蛋白酶基因家族(ADAM)和卵子表面的整合素蛋白.本文通过对精子表面的受精素(It,受精素B,cyritestin,卵子表面的ot6131,CD9等蛋白分子的研究,揭示了这些分子对粘附,融合的重要作用,为提高受精率提供了重要的依据.关键词受精;粘附;融合;分子基础中图分类号Q492哺乳动物受精时精子进入雌性生殖道与卵子相遇,精子最先与卵子质膜接触的部位是顶体内膜,然后是赤道段/顶体后区的质膜,卵子参与精卵质膜融合的部位一般是微绒毛丰富的区域.根据对精卵质膜融合模式的研究推测,卵子表面的整合素和精子表面的去整合素是粘附所需要的.一,精子表面去整合素蛋白受精素(fertilin)是精子表面去整合素的一种,它是在筛选抗豚鼠精子质膜融合的单克隆抗体时得到的,位于精子头的后区,即精子参与融合的部位.在该研究中发现单抗PH一3O可以降低豚鼠受精率并呈现出剂量的依赖性,当PH一3O浓度为1401~g/ml时可以达到阻断质膜融合的最大效率73%(Prima—koff等.1987).PH-30是由两个结构非常相似的亚基组成的异二聚体:fertilinot(ADAM1)和fertilin13(ADAM2).分析成熟蛋白及其前体的一级结构,确定了一个以fertilinot和fertilin13为代表的去整合素一金属蛋白酶基因家族(adisintegrinandmetallopro—teasegenefamily,ADAM).ADAM家族成员中有保守型结构域,包括信号序列,前区,金属蛋白酶结构域,去整合素结构域,富含半胱氨酸结构域,EGF样重复序列,跨膜结构域和短的胞质尾部¨.Cyrites—tin也是ADAM家族成员,定位于精子顶体内膜或精子赤道段,由其中的去整合素结构域参与精卵之间的反应(Barker等.1994).对fertilinot,fertilin13和cyritestin很多功能性的研究都表明,它们是参与精卵质膜粘附,融合作用的候选分子.在人工体外受精(IVF)实验中,利用抗体,肽及单独的蛋白等多种反应物来检测去整合素在精卵质膜反应中的作用.用精子fertilinot,fertilin13和cyritestin重组体与卵子人工体外受精,粘附和融合都被抑制;当fertilin13去整合素结构域合成肽参与受精作用时,对质膜间粘附和融合反应只产生微弱抑制作用,而cyritestin肽对质膜粘附和融合的阻断程度却很高,从其他蛋白(fertilinot,AD—AM4和ADAM5)中提取的肽的阻断作用较低或不发生阻断.这种阻断作用可能由于肽结合在卵受体上(可能是整合素)的过程中有特定的步骤被阻断.6TaoY,ZhouB,XiaGL,eta1.ExposuretoL—ascorbicacid ord—tocopherolfacilitatesthedevelopmentofporcinedenu—dedoocytesfrommetaphaseItometaphaseIIandprevents cumuluscellsfromfragmentation.ReprodDomestAnim, 2004.39:52~57.7Y ousefMI,AbdallahGA,KamelKI.Effectofascorbicacid andVitaminEsupplementationonsemenqualityandbio—chemicalparametersofmalerabbits.AnimReprodSci,2003.76:99~111.8TarinJJ,Perez—MbalaS,Garcia—PerezMA,eta1.Effectof dietarysupplementationwithamixtureofVitaminsCandE onfertilizationoftertiarybutylhydroperoxide—treatedoocytes andparthenogeneticactivationinthemouse.Theriogenology, 2002.57:869~881.9TsujiiH,MuranakaM,HamanoK.Cultureofinvitro mouseembryoswithvitaminEimprovesdevelopment.JRe—prodDev.2002.48:25~29.10TarinJJ,Perez—AlbalaS,PertusaJF,eta1.Oraladminis—trationofpharmacologicaldosesofvitaminsCandEreduces reproductivefitnessandimpairstheovariananduterine functionsoffemalemice.Theriogernology,2002,57:】539~】550.。

辛建增,唐婷,刘盛.PGC-lα调控畜禽肌肉脂肪生长代谢及其与肉品质研究进展[J].畜牧与兽医,2024,56(5):138-145.XINJZ,TANGT,LIUS.Progressinresearchonrelationshipbetweenregulationofperoxisomeproliferator-activatedreceptorγ-coactivator-1αongrowthandmetabolismofmuscleandfatandmeatqualityinlivestockandpoultry[J].AnimalHusbandry&VeterinaryMedicine,2024,56(5):138-145.PGC-lα调控畜禽肌肉脂肪生长代谢及其与肉品质研究进展辛建增1,唐婷1,刘盛2∗(1.烟台大学生命科学学院,山东烟台㊀264000;2.烟台大学药学院,山东烟台㊀264000)摘要:过氧化物酶体增殖物激活受体γ辅激活因子1α(PGC-lα)是一种具有广泛功能的转录调节因子,其在动物体内参与线粒体生物合成㊁肌纤维类型转化㊁脂肪分化㊁肌内脂肪沉积㊁糖脂代谢㊁能量代谢等多项生理过程,其中,肌纤维类型和肌内脂肪含量与肉品质密切相关㊂因此,在分子水平深入探究PGC-1α调控肌肉和脂肪的生长代谢过程将为改善肉品质提供新的研究思路㊂本文系统概述了PGC-lα的结构特点及PGC-1α调控肌肉线粒体增生㊁脂肪分化㊁能量代谢等过程的机制,重点介绍了PGC-lα调控肌纤维类型转化㊁肌内脂肪沉积㊁糖类代谢及其与肉品质形成之间的可能关系,以期为今后通过PGC-1α调控畜禽肌肉脂肪生长代谢,进而改善肉品质提供参考㊂关键词:过氧化物酶体增殖物激活受体γ辅激活因子1α;肌纤维类型;肌内脂肪沉积;能量代谢;肉品质中图分类号:S826㊀㊀㊀文献标志码:A㊀㊀㊀文章编号:0529-5130(2024)05-0138-08Progressinresearchonrelationshipbetweenregulationofperoxisomeproliferator-activatedreceptorγ-coactivator-1αongrowthandmetabolismofmuscleandfatandmeatqualityinlivestockandpoultryXINJianzeng1,TANGTing1,LIUSheng2∗(1.CollegeofLifeSciences,YantaiUniversity,Yantai264000,China;2.CollegeofPharmacy,YantaiUniversity,Yantai264000,China)Abstract:Peroxisomeproliferator-activatedreceptorγ(PPAR-γ)coactivator1α(PGC-lα)isaversatiletranscriptionalregulator.Thisregulatorisinvolvedinmanyphysiologicalprocessessuchasmitochondrialbiosynthesis,musclefibertypetransformation,adiposedifferenti⁃ation,intramuscularadiposedeposition,glycolipidmetabolism,andenergymetabolisminanimals.Musclefibertypeandintramuscularfatcontentarecloselyrelatedtomeatquality.Therefore,exploringtheregulationofPGC-1αonthegrowthandmetabolismofmuscleandfatatthemolecularlevelwillprovidenewresearchideasforimprovingmeatquality.Inthispaper,thestructuralcharacteristicsofPGC-lαandthemechanismofPGC-1αregulatingmusclemitochondria,adiposedifferentiationandenergymetabolismaresystematicallyreviewed.Theregu⁃lationofPGC-lαonmusclefibertypetransformation,intramuscularfatdeposition,carbohydratemetabolismanditspossiblerelationshipwiththeformationofmeatqualityareemphasized;whichprovidesreferenceforimprovingmeatqualitybyregulatingthegrowthandmetabo⁃lismofmuscleandfatbyPGC-1αinlivestockandpoultry.Keywords:PGC-1α;musclefibertype;intramuscularfatdeposition;energymetabolism;meatquality㊀㊀畜禽肉品质包括肉色㊁嫩度㊁系水力㊁风味㊁多汁性等多个方面㊂因此,肉品质性状是一个复杂的综合性状㊂肉品质受宰前和宰后多种因素的影响,例如遗传(品种㊁性别㊁年龄㊁基因)㊁营养水平㊁饲养管理㊁宰前运输㊁屠宰方式㊁宰后成熟方式等,其中㊀收稿日期:2023-05-25;修回日期:2024-03-20基金项目:烟台大学博士启动基金项目(SM20B113)第一作者:辛建增,男,博士,讲师∗通信作者:刘盛,讲师,研究方向为食品化学,E-mail:liush⁃eng87@126 com㊂遗传因素起决定性作用㊂然而,在饲养过程中,畜禽肌肉和脂肪的生长发育及代谢对肉品质的形成也起着至关重要作用㊂畜禽肌肉的生长发育及代谢是一个及其复杂的过程,由多种基因和信号通路在不同水平上参与调控,各调控因子与信号通路分工协作组成精细复杂的调控网络,有序调控肌肉的生长发育㊁肌纤维类型的转化㊁肌纤维的能量代谢等生物学过程㊂而脂肪组织是畜禽维持生命活动必不可少的组织,通常储存在皮下㊁内脏㊁肌肉等部位㊂与肉品质最相关的脂肪为肌内脂肪和肌间脂肪㊂其中肌内脂肪的含量与肉品质最为密切,是肉品领域的研究热点,肌内脂肪的含量会影响肉的系水力㊁风味㊁多汁性等品质㊂过氧化物酶体增殖物激活受体γ辅激活因子1α(PGC-1α)是肌肉和脂肪生长代谢过程中必需的转录共调节因子,它参与调控肌细胞线粒体生物合成㊁肌纤维类型的转化㊁肌细胞能量代谢等生物学过程㊂PGC-1α在脂肪的分化㊁沉积㊁合成㊁代谢等方面也发挥重要的调节作用㊂此外,PGC-1α还参与机体的适应性产热㊁肝脏的糖异生㊁血管生成㊁调控细胞中活性氧簇水平㊁调控机体的生物钟基因等生理过程㊂PGC-1α功能广泛,参与众多生理调节过程㊂本文将对PGC-1α分子结构特征,PGC-1α调控肌纤维能量代谢㊁肌纤维糖代谢㊁肌纤维类型转化㊁脂肪分化㊁肌内脂肪沉积㊁脂肪代谢及其与宰后肉品质的可能关系进行了系统阐述,并对相关可能的研究热点进行了展望㊂以期为更深入地探究PGC-1α信号通路及其靶基因调控畜禽肌肉脂肪生长代谢和提高肉品质提供参考㊂1㊀PGC-1α概述PGC-1α是由Spiegelman团队1998年最先在小鼠棕色脂肪组织中发现的一种转录共调节因子[1]㊂PGC-1α属于PGC-1家族,该家族共有3个成员,另外两个分别为过氧化物酶体增殖物激活受体γ(PPAR-γ)辅激活因子-1β(PGC-1β)和PGC-1相关辅活化因子(PRC),其家族成员蛋白长度存在着一定的差异,但存在着相应的保守序列㊂PGC-1家族的N端结构域均含有转录激活域,C端结构域均包含富含丝氨酸/精氨酸的RS域和RNA结合区域(RMM)[2]㊂PGC-1α与PGC-lβ同源性较高,而与PRC的同源性则相对较低㊂人的PGC-1α基因位于染色体4p15 1区域,全长为681kb,由13个外显子和12个内含子组成,其mRNA含有6908bp,编码一个包含798个氨基酸,分子量91kDa的蛋白质[3],其他常见畜禽的PGC-1α基因与蛋白质基本信息见表1(引自NCBI)㊂PGC-1α的蛋白结构域,其N端有一个富含酸性氨基酸的转录激活区(activationdomain,AD),该区内有一个LXXLL结构域(X:任意氨基酸;L:亮氨酸),此结构域是PGC-1α与配体依赖型核受体结合的基础㊂负调控元件和转录因子结合位点位于PGC-1α的中间区域,当转录因子与PGC-1α结合时,负调控元件就会暴露出来[4]㊂C末端是一个RNA结合基本序列RRM和富含丝氨酸/精氨酸的RS区域,这个区域可以与RNA聚合酶Ⅱ的C末端相互作用,处理新转录的RNA㊂PGC-1α上还有与细胞呼吸因子(NRF)㊁肌细胞特异性增强子2C(myocyteenhancerfactor2C,MEF2C)及PPARγ结合的位点[3]㊂因此,PGC-1α是作为转录因子的激活因子来调控其他基因的表达㊂表1㊀人与常见畜禽PGC-1α基因和蛋白质基本信息物种所处染色体基因长度/kbmRNA长度/bp内含子数外显子数蛋白肽链长度(氨基酸残基数量)蛋白质分子量/kDa人46816908121380392猪86966738121379690狗36415841131480391牛67156324121379690羊67186680121378789鸡43486615121380892鸭43619716121380892鸽子43644913121367077㊀㊀PGC-1α分子本身的促转录激活活性较低,只有被相应的受体募集后,其活性才显著增强㊂PGC-1α与核受体结合后,会导致PGC-1α构象发生改变,并与下游因子作用,发挥转录激活作用㊂PGC-1α不仅对PPARγ具有组织特异性的辅激活作用,而且也是类维生素AX受体(RXR)㊁肌细胞增强因子2c(myocyteenhancerfactor2C,MEF2C)㊁甲状腺激素受体(thyroidhormonereceptor,TR)㊁糖皮质激素受体(glucocorticoidreceptor,GR)㊁雌醇受体α(es⁃trogenreceptor,ERα)和PPARs等核受体(nuclearreceptor,NR)的辅激活因子[2,5-7]㊂PGC-1α的表达具有组织特异性,通常在线粒体含量丰富和氧化代谢活跃的器官或组织中高表达,如骨骼肌㊁心脏㊁棕色脂肪组织㊁肝脏㊁肾脏和大脑组织等,而在肺㊁小肠㊁结肠和胸腺中只有很少量的表达,在胎盘㊁脾和外周白细胞中未见表达[8]㊂前已述及,PGC-1α在肌肉脂肪的生长发育及代谢中发挥着重要调控作用,下面将针对其活性调控㊁肌肉脂肪生长代谢及其与肉品质和一些生理功能的相关作用进行论述㊂2㊀PGC-1α活性调控相关信号因子PGC-1α含有磷酸化㊁乙酰化㊁糖基化㊁甲基化㊁泛素化等翻译后修饰的位点,这些翻译后修饰对于其发挥作用时的精细化调控具有重要意义[9]㊂其中当前研究较多的为乙酰化和磷酸化修饰㊂沉默信息调节因2相关酶1(sirtuin1,SIRT1)和AMP依赖的蛋白激酶(adenosine5-monophosphate-activatedproteinkinase,AMPK)是调控PGC-1α去乙酰化和磷酸化的关键酶,此两种酶对于机体肌肉脂肪生长发育和能量代谢的精准调控和稳态维持具有重要的意义㊂SIRT1可以将乙酰化后的PGC-1α去乙酰化,从而提高PGC-1α的活性[10-11]㊂此外SIRT1是体内代谢的感受器,当机体处于能禁食或者饥饿等状态下,SIRT1会加速PGC-1α的去乙酰化,导致其活性上升,可增加线粒体的合成㊂而一些乙酰转移酶例如组蛋白乙酰化酶氨合成通用控制蛋白5(histoneacetyl⁃transferaseGCN5,GCN5)和核受体共激活因子-3(steroidreceptorcoactivator3,SRC-3)可以使PGC-1α发生乙酰化,从而抑制其活性[12-15]㊂此外,SIRT1的去乙酰化作用还是PGC-1α调控生物钟基因表达的重要事件㊂SIRT1与乙酰化酶协调作用,精细化调节PGC-1α发挥作用㊂AMPK是体内能量感受器,当机体能量处于缺乏状态时,AMPK可使PGC-1α磷酸化位点磷酸化,从而提高PGC-1α活性,激活与能量代谢相的通路,引起线粒体增生㊁脂肪酸氧化等生物学过程增加[14]㊂3㊀PGC-1α与肌肉生长代谢及肉品质3 1㊀PGC-1α与肌肉线粒体合成及肉品质线粒体是为骨骼肌生长发育提供能量的细胞器,它对骨骼肌发挥正常生理功能具有重要的意义,PGC-1α是调控线粒体生物合成和氧化磷酸化过程中的关键调节因子[15-16]㊂研究发现,PGC-1α可参与调控肌纤维中线粒体的生成,并且还能够调节线粒体的融合及分裂,在某些组织,如白色脂肪㊁肌肉㊁神经㊁心脏中超表达PGC-1α,都会促进线粒体的生成[15-17]㊂PGC-1α促进线粒体生成主要通过与转录因子结合发挥作用,常见的为核呼吸因子-1(nuclearrespiratoryfactor-1,NRF-1)和核呼吸因-2(nuclearrespiratoryfactor-2,NRF-2)㊂研究发现,PGC-1α与核呼吸因子结合后会刺激线粒体转录因子A(mitochondrialtranscriptionfactorA,mtTFA)的合成㊂这些因子直接影响线粒体生成,在线粒体内引起线粒体DNA的双向转录,实现了线粒体的增殖[18-19]㊂畜禽宰杀放血后,肌肉中的线粒体发生肿胀,最终结构破坏而破裂,但肉品质形成过程中,线粒体的生理代谢状态与肉嫩度㊁肉色㊁持水力等品质有着密切关系㊂研究表明,宰后初期肌肉线粒体耗氧率与肉品嫩度密切相关,高嫩度牛肉拥有更高的线粒体耗氧率[20]㊂宰后肌肉中线粒体影响肉色稳定性主要通过两种途径,一是线粒体与氧合肌红蛋白竞争氧气,使其转变为脱氧肌红蛋白状态,此情况过度发生可导致肉色变暗;另一方面,线粒体具有高铁肌红蛋白还原酶活性,可以将氧化的高铁肌红蛋白转化为还原态脱氧肌红蛋白,为鲜红色氧合肌红蛋白的生成提供还原态肌红蛋白[21-22]㊂肌肉持水力是肉品一个重要的品质,最近研究表明,牛肉宰后成熟过程中,线粒体脂肪成分的变化与肌肉持水力的变化密切相关[23]㊂PGC-1α已被证明其与畜禽生长和肉品质密切相关,且已被列为能够候选基因[24],然而未见PGC-1α调控肌肉中线粒体与宰后肉品质的相关研究,PGC-1α对肌肉中线粒体的调控及宰后肉品质的变化形成需要开展深入研究㊂3 2㊀PGC-1α与肌肉糖类代谢葡萄糖是肌肉组织主要的能源物质,糖类氧化供能为肌肉的各类生理活动提供能量㊂PGC-1α在体内糖代谢的过程中发挥重要调节作用,主要表现在以下几个方面:首先PGC-1α是糖异生过程的关键调节因子㊂在禁食情况下,PGC-1α会在肝细胞中大量表达,与其他相关调节因子配合在转录水平上激活糖异生关键酶组,如葡萄糖-6-磷酸酶㊁磷酸烯醇式丙酮酸羧激酶等,最终导致肝糖输出增加[25-26]㊂其次,葡萄糖进入肌肉细胞需要葡萄糖转运载体4(glucosetransporters4,GluT4)的转运,PGC-1α可与肌细胞增强子因子2(myocyteenhancerfactor2,MEF2)共同作用,刺激GluT4的表达,从而增加肌细胞内葡萄糖的水平㊂此外,PGC-1α在某些情况还可抑制肌细胞葡萄糖的氧化,其与雌激素相关受体(estrogen-relatedreceptorα,ERRα)结合后,刺激丙酮酸脱氢酶4表达,从而抑制葡萄糖氧化和增加葡萄糖吸收来补充肌糖原贮备,为下一次的肌肉运动做准备㊂肌肉中的糖原是宰后生成乳酸的原料,动物胴体在宰后冷藏排酸过程中,糖原转化为乳酸导致肌肉pH值下降,这是宰后肌肉排酸的原理㊂而宰后pH的下降幅度和速度影响肉品质形成,宰后肌肉pH值过高或过低都会形成异质肉㊂而PGC-1α对于肌肉糖代谢具有调控作用,宰前肌肉中PGC-1α的表达水平和活性对于宰后肌肉糖原水平㊁pH值变化及肉品质形成是否具有影响,未见相关报道,需要开展相应研究㊂3 3㊀PGC-1α与骨骼肌肌纤维类型转换及肉品质不同肌纤维类型对于肌肉发挥生理功能具有重要的作用,比较常见的例子是,动物不同部位的肌肉的肌纤维组成存在着明显差异,且肉品质也存着差别㊂肌肉纤维类型受遗传㊁运动㊁营养㊁和环境等多种因素的影响㊂PGC-1α是调控肌纤维类型转变的主要因子,PGC-1α基因高表达,可以提高与氧化型肌纤维有关的基因表达,提高细胞色素C和肌红蛋白的含量提高有氧呼吸能力与线粒体的数量,增强抗疲劳的能力等,主要为使酵解型肌纤维向氧化型肌纤维转化[27-28]㊂超表达PGC-1α的转基因小鼠,其骨骼肌中Ⅱ型肌纤维表现出Ⅰ型肌纤维的蛋白特性,其中TNN1蛋白㊁肌红蛋白和肌钙蛋白Ⅰ明显增加,Ⅱ型肌纤维逐步转化为Ⅰ型肌纤维[29]㊂人和动物的骨骼肌类型变化研究表明,PGC-1α的表达量与快肌纤维的含量成负相关,与慢肌纤维的含量成正相关[30-31]㊂相关研究已证实,寒冷可以刺激诱使鸡的胸肌部分从ⅡB型转化为ⅡA型,而PGC-1α的上调表达在其中发挥了关键的作用[32]㊂PGC-1α通过调节肌纤维类型影响畜禽肉品质已经被证实,但是其发挥作用的详细分子机制还不清晰,需要开展相应的深入研究㊂3 4㊀PGC-1α与肌肉中活性氧含量及肉品质PGC-1α可促进肌肉等组织中线粒体的合成,还能刺激线粒体呼吸链电子转运活性,从理论上讲,PGC-1α将导致细胞内活性氧(reactiveoxygenspecies,ROS)水平提高,但是实际上并非如此,在肌肉和棕色脂肪中,运动与寒冷环境的暴露均和ROS负面影响没有关联,这主要是PGC-1α可以增强很多抗氧化酶的表达[33-34]㊂即PGC-1α有两种能力,刺激线粒体电子转运的同时抑制ROS水平㊂这样,肌肉组织,棕色脂肪通过提升线粒体代谢应对外部环境变化的过程中,不会对自身造成氧化损伤㊂而ROS与宰后肉品的形成密切相关,动物在宰杀后,ROS主要来源于线粒体和脂肪的氧化,产生的ROS往往会对某些肉品质,肉色㊁嫩度㊁系水力等产生负面影响[23,35]㊂ROS与宰后肉品质形成一直是肉品科学领域研究的热点,PGC-1α已被证实是影响肉品质的候选基因之一,但是其调控宰后肌肉中ROS的作用机制及如何影响肉品质未见相关报道㊂4㊀PGC-1α与脂肪生长代谢及肉品质4 1㊀PGC-1α与脂肪细胞分化动物脂肪组织中大约1/3是脂肪细胞,其余的2/3是成纤维细胞㊁微血管㊁神经组织和处于不同分化阶段的前脂肪细胞㊂由前脂肪细胞分化为脂肪细胞的过程是一个涉及多个信号通路的复杂调控过程,该过程大致可为4个阶段,分别为生长抑制阶段㊁克隆扩增㊁早期分化和终末分化[36]㊂PPARs在动物脂肪发育分化的早期分化阶段开始发挥调控作用,它们与相应的因子协调作用,共同调节脂肪的增殖分化㊂PPARγ是PPARs家族成员,它是脂肪细胞分化的及其的重要因子,其通常可作为前体脂肪分化处于早期分化的标志基因,是脂肪细胞增殖分化过程中起决定性作用的基因㊂研究证实,PPARγ缺失的胚胎干细胞能够分化为多种细胞,但唯独不能分化为脂肪细胞㊂此外,PPARγ基因敲除的小鼠,在胚胎期10d左右就会死亡,且未在胚胎内检测到脂肪细胞,而正常小鼠在胚胎期10d即可检测到脂肪细胞的存在[36]㊂这说明PPARγ在脂肪分化形成过程中起关键作用,PPARγ发挥脂肪分化调控作用时,需要先与RXRα形成异源二聚体,然后与所调节基因启动子上游的过氧化物酶体增殖物反应元件(PPRE)结合才发挥转录调控作用,而PGC-1α作为PPARγ配体,能促进PPARγ与相应调控因子的结合[37]㊂很多哺乳动物体内存在着白色脂肪组织㊁米色脂肪组织和棕色脂肪组织三种,白色脂肪主要作用为贮存能量,米色脂肪具有贮存能量和非战栗产热的功能,棕色脂肪主要进行非战栗产热㊂在细胞结构和功能上,白色脂肪细胞拥有一个大脂滴用于存贮能量,而棕色脂肪细胞拥有多脂滴㊁多线粒体的结构㊂PGC-1α能够促进白色脂肪向棕色脂肪转化,它能够刺激白色脂肪中线粒体的大量生成,还能增加解偶联蛋白1(UCP1)等分子的生成,这些改变可使白色脂肪逐渐转化为棕色脂肪组织[38]㊂4 2㊀PGC-1α与脂肪氧化供能脂肪是畜禽体内重要的储能物质,在冷暴露㊁禁食㊁运动等情况下,可为机体提供能量,其中脂肪酸β氧化产能是其最为主要的供能方式㊂脂肪是也骨骼肌获取能量的重要物质㊂研究表明,过表达PGC-1α可增加骨骼肌线粒体的生物合成,也可使脂肪酸氧化相关酶含量上升或者活性增强,从而增加脂肪酸氧化供能[39-40]㊂在小鼠骨骼肌和猪前脂肪细胞过表达PGC-1α,可促进脂肪酸氧化过程中相关基因肉碱棕榈酰转移酶1β(CPT1β)㊁肝型脂肪酸结合蛋白(FABP1)㊁过氧化物酶酰基辅酶A氧化酶1(ACOX1)㊁中链酰基辅酶A脱氢酶(MCAD)㊁脂肪酸转位酶(CD36)等的表达,其中CPT1β是脂肪酸氧化过程中的限速酶[38-41]㊂CD36㊁FABP1是脂肪酸转运的重要蛋白,可将脂肪酸逐步转运至肌肉等组织,便于氧化供能㊂而ACOX1㊁MCAD是参与脂肪酸氧化过程中的关键酶㊂过表达PGC-1α还可促进氧化磷酸化相关基因ATPSynthase㊁CytC㊁COXⅢ等的表达[27]㊂而在PGC-1a敲除后的小鼠表现为心脏功能不全,肌肉耐力下降,轻度心动过缓,心肌脂肪酸氧化能力下降,能量产生减少[42-44]㊂以上研究说明PGC-1α在肌肉的脂肪酸氧化供能方面起重要的调节作用㊂4 3㊀PGC-1α与肌内脂肪沉积及肉品质肌内脂肪的沉积是一个涉及多种信号通路和代谢因子的复杂过程,PPARs家族成员㊁肌内脂肪转运相关因子等发挥了重要的作用㊂PGC-1α是PPARs家族某些因子的配体,其在肌肉脂肪代谢过程中发挥了重要作用㊂PGC-1α不仅能够增加肌肉脂肪的分解代谢(前已述及),而且还可增加肌细胞中脂肪的合成代谢㊂通过肌细胞培养实验和转基因小鼠试验证实,PGC-1α不仅能增加脂肪的分解代谢,还可以增加肌细胞内脂肪酸和磷脂等脂肪的合成代谢[45-46],且PGC-1α转基因小鼠的脂肪酸转运蛋白等脂质代谢相关蛋白也增加了[46]㊂PGC-1α对于肌内脂肪的双向调控作用,对于动物维持生命活动具有重要的意义,不仅能够保障机体对于能量的需求,还对机体后续的生命活动具有重要的意义㊂其发挥脂肪调控作用,还要取决于动物机体所处的状态㊂畜禽上的相关研究已经证实,PGC-1α与脂肪沉积及肉品质存在一定关联㊂在猪上的研究表明,PGC-1α参与猪脂肪沉积的基因,PGC-1α基因多态性与失水率㊁剪切力等肉品指标显著相关[47-49]㊂因此,PGC-1α已被列为猪脂肪沉积及肉品质的候选基因,且在藏猪上的研究表明PGC-1α与肌内脂肪沉积密切相关[36]㊂在鸡上的研究也证实,PGC-1α多态性与鸡腹部脂肪的沉积显著相关[50-51]㊂然而,在牛上的研究表明,肌内脂肪含量及嫩度等品质与PGC-1α存在一定的相关性,但是未达到显著水平[52]㊂以上研究表明由于遗传背景的差异,不同畜禽PGC-1α在调控肌肉脂质代谢方面可能存在着差异㊂但是当前研究大多停留在分析推测层面,并未对其作用的机理及信号通路作用方式进行深入研究,因此需要对PGC-1α调控肌肉代谢,尤其是调控脂肪代谢开展深入的研究,为优质肉品的生产提供研究基础㊂4 4㊀PGC-1α与机体的适应性产热适应性产热是机体应对外界刺激以产热的形式消耗能量的生理过程,对于动物在特定环境下,维持正常体温和生命活动是必须的,主要发生在骨骼肌和棕色脂肪组织㊂其中小型动物,如小鼠,大鼠等主要依靠棕色脂肪组织进行适应性产热,而畜禽则以肌肉适应性产热为主㊂棕色脂肪的分化形成需要PPARγ发挥作用,但其发挥作用需要PGC-1α的辅助,PGC-1α结合并激活PPARγ后才能刺激棕色脂肪细胞分化过程中基因的转录[15,53-54]㊂PGC-1α还可通过另外两个方面来加快适应性产热,首先是促进适应性产热原料的摄取,促进棕色脂肪和肌肉对产热原料,如葡萄糖和脂肪的摄取;促进适应性产热过程中关键因子的合成及表达,主要是为了适应性产热过程的顺利进行,如促进线粒体的生物合成,促进呼吸链相关基因的表达,促进氧化磷酸化相关基因的表达等[55-56]㊂当前未见PGC-1α调控畜禽适应性产热与肉品质的相关研究,但宰后迅速科学降低屠体的温度,防止肉品质因为过热而出现变质是当前肉品科学领域的一个重要的研究方向㊂5㊀PGC-1α与生物钟相互反馈调控畜禽骨骼肌代谢㊀㊀生物钟是生物机体生命活动的内在节律性㊂体温㊁血压㊁睡眠㊁内分泌㊁肝脏代谢㊁行为等重要生命活动均受到生物钟相关基因的调控[57-59],研究表明生物钟还可参与调控细胞周期[60]㊂其中昼夜节律及光照是调节生物钟基因表达的最常见的外部环境因素,这些因素的变化会影响畜禽的生长发育和动物性产品的质量㊂生物钟相关调控规律已在畜禽生产领域得到了应用,其可用于改善动物的生长,提高动物性产品的质量㊂Tao等[61]的研究表明,生物钟基因在蛋鸭卵巢的表达水平与产蛋量密切相关㊂光刺激可通过影响生物钟基因的表达,提高肉仔鸡生长期体重和胸肌产量,改善饲料转化率[62]㊂生物钟基因与奶山羊乳腺代谢密切相关,饲喂不同饲料可改变调生物钟基因表达,调控奶山羊的泌乳[63]㊂畜禽骨骼肌中存在着生物钟基因,骨骼肌的生命活动受到生物钟基因的调控,PGC-1α是连接生物钟和能量代谢的关键调控因子[64]㊂研究表明,PGC-1α在骨骼肌中的表达呈现明显的昼夜节律性,且PGC-1α敲除小鼠在能量代谢方面出现异常的生理节律㊂PGC-1α与生物钟基因形成反馈调节回路,首先PGC-1α是生物时钟基因的上游调节因子,PGC-1α能够诱导生物时钟关键基因的表达,如脑和肌肉芳香烃受体核转运样蛋白1基因(Bmal1)㊁时钟基因(Clock)和反向成红细胞增多症基因(Rev-erba)等㊂此外,PGC-1α还可以和视黄酸受体相关的孤儿受体(RORα/γ)协同作用,使染色质的局部结构活化,从而激活Bmal1的转录[65]㊂此外,SIRT1对PGC-1α的去乙酰化是导致Bmal1激活的关键事件[66]㊂其次,Clock1a:Bmal1b复合体又能参与调控PGC-1α的表达㊂在畜禽骨骼肌中生物钟基因与PGC-1α共同调节骨骼肌的糖脂和能量代谢等生命活动,对于畜禽骨骼肌的生长发育具有重要的意义㊂当前缺乏PGC-1α与生物钟基因联合作用调控畜禽肉品质的相关入研究,这可能会成为肉品领域新的研究方向㊂6 小结与展望综上所述,PGC-1α作为一种多效转录调控因子,除参与调控肌肉脂肪生长发育及能量代谢外,还参与骨骼肌脂肪的沉积㊁肌纤维类型转化等生理活动,不仅能够在转录水平上调控骨骼肌能量代谢,而且还与生物钟基因相互作用反馈调节肌肉脂肪的生长发育㊂近年来随着我国人民水平的提高和饮食结构的改善,对于肉品质提出了更高的要求,例如肉品嫩度㊁多汁性和大理石花纹等,这些品质与肌纤维类型和肌内脂肪含量密切相关㊂如何生产肌纤维类型比例合适㊁肌内脂肪适中的肉品,是当前动物营养领域和肉品科学领域的研究热点㊂这与骨骼肌和脂肪生长代谢显著相关,且PGC-1α在其中发挥了重要作用㊂尽管针对PGC-1α调节骨骼肌生长发育㊁肌纤维类型转换㊁脂肪沉积㊁能量代谢的分子机制,已进行了大量的系统研究,也取得了一些重大进展,但还存在许多问题,诸如PGC-1α如何精细调节肌内脂肪沉积,PGC-1α调控肌纤维转换和能量代谢的详细信号通路,以及PGC-1α与脂肪因子瘦素㊁脂联素㊁抵抗素等的相互激活转录机制,特别是如何通过有效地干预PGC-1α调控肌肉脂肪沉积及靶向控制PGC-1α介导肌纤维类型转换等㊂今后需对这些问题进行深入探索,以期通过PGC-1α调控畜禽肌肉的生长发育㊁脂肪代谢㊁能量代谢等生理过程来提高肉品质㊂参考文献:[1]㊀MITRAR,NOGEEDP,ZECHNERJF,etal.Thetranscriptionalcoactivators,PGC-1αandβ,cooperatetomaintaincardiacmito⁃chondrialfunctionduringtheearlystagesofinsulinresistance[J].JMolCellCardiol,2012,52(3):701-710.[2]㊀JANNIGPR,DUMESICPA,SPIEGELMANBM,etal.Regula⁃tionandbiologyofPGC-1α[J].Cell,2022,185(8):1444.[3]㊀ESTERBAUERH,OBERKOFLERH,KREMPLERF,etal.Humanperoxisomeproliferatoractivatedreceptorγcoactivator1(PPARGC1)gene:cDNAsequence,genomicorganization,chro⁃mosomallocalizationandtissueexpression[J].Genomics,1999,62(1):98-102.[4]㊀PUIGSERVERP,RHEEJ,LINJ,etal.Cytokinestimulationofenergyexpenditurethroughp38MAPkinaseactivationofPPARγco⁃activator-1[J].MolCell2001,8:971-982.[5]㊀TCHEREPANOVAI,PUIGSERVERP,NORRISJD,etal.Modu⁃lationofestrogenreceptor-αtranscriptionalactivitybythecoactivatorPGC-1[J].BiolChem,2000,275(21):16302-16308.㊀[6]㊀BHALLAS,OZALPC,FANGS,etal.Ligand-activatedpregnaneXreceptorinterfereswithhnf-4signalingbytargetingacommonco⁃activatorPGC-1α:functionalimplicationsinhepaticcholesterolandglucosemetabolism[J].BiolChem,2004,279(43):45139-45147.㊀[7]㊀RHEEJ,INOUEY,YOONJC,etal.RegulationofhepaticfastingresponsebyPPARγcoactivator-1α(PGC-1α):requirementforhepatocytenuclearfactor4αingluconeogenesis[J].ProcNatlAcadSciUSA,2003,100(7):4012-4017.[8]㊀马燕.藏羚羊和藏系绵羊PGC-1α基因编码区的克隆与分析[D].西宁:青海大学,2012.[9]㊀张林.超表达猪源PGC-1α促进小鼠和猪肌纤维类型转变的研究[D].武汉:华中农业大学,2014.[10]RODGERSJT,LERINC,HAASW,etal.Nutrientcontrolofglu⁃cosehomeostasisthroughacomplexofPGC-1αandSIRT1[J].Nature,2005,434(7029):113-118.[11]WANGW,WUD,DINGJ,etal.Modifiedrougandecoctionatten⁃uateshepatocyteapoptosisthroughamelioratingmitochondrialdys⁃functionbyupregulatedSIRT1/PGC-1αsignalingpathway[J].PoultSci,2023,102(10):1-19.[12]LERINC,RODGERSJT,KALUMEDE,etal.GCN5acetylrans⁃ferasecomplexcontrolsglucosemetabolismthroughtranscriptionalrepressionofPGC-1α[J].CellMetab,2006,3(6):429-438.[13]YEF,WUL,LIH,etal.SIRT1/PGC-1αisinvolvedinarsenic-inducedmalereproductivedamagethroughmitochondrialdysfunction,whichisblockedbytheantioxidativeeffectofzinc[J].EnvironPollut,2023,320:121084-121086.[14]NETOIVS,PINTOAP,MUNOZVR,etal.Pleiotropicandmulti-systemicactionsofphysicalexerciseonPGC-1αsignalingduringtheagingprocess[J].AgeingResRev,2023,87:101935-101954.㊀[15]PUIGSERVERP,WUZ,PARKCW,etal.Acold-inducibleco⁃activatorofnuclearreceptorslinkedtoadaptivethermogenesis[J].Cell,1998,92(6):829-39.[16]LIL,LUZ,WANGY,etal.Genisteinalleviateschronicheatstress-inducedlipidmetabolismdisorderandmitochondrialenergeticdys⁃functionbyactivatingtheGPR30-AMPK-PGC-1αsignalingpath⁃waysintheliversofbroilerchickens[J].PoultSci,2023,103(1):1-12.[17]GARNIERA,FORTIND,ZOLLJ,etal.Coordinatedchangesin。

HAS2和PTGS2在牦牛卵巢中的定位表达及FGF10对其表达影响HAS2和PTGS2在牦牛卵巢中的定位表达及FGF10对其表达影响引言:牦牛是高原域内的重要畜牧动物,其生殖系统的正常发育对其育种和繁殖具有重要意义。
卵巢是牦牛生殖系统的重要组成部分,卵巢中包含多种关键基因,调控着卵巢的正常发育和功能。
本文旨在研究卵巢中HAS2和PTGS2两个基因的定位表达,并探究FGF10对其表达的影响。
一、HAS2和PTGS2的定位表达在牦牛卵巢中的意义1. HAS2基因因为发育和繁殖过程需求大量的透明质酸(Hyaluronic acid,HA),而HAS2编码的酶是合成透明质酸的关键酶。
透明质酸在卵巢中的分布和水平对卵泡的形成和发育起到重要作用。
早期研究表明,在牦牛卵巢中HAS2的表达主要发生在颗粒层细胞和卵泡膜细胞中,这提示HAS2可能在卵泡发育中发挥着重要作用。
2. PTGS2基因PTGS2基因编码环氧合酶2(prostaglandin-endoperoxide synthase 2),也被称为炎症素合酶2。
研究表明,PTGS2可以调节卵巢中前列腺素的合成,并且参与调控卵巢发育和卵巢周期。
在牦牛的卵巢中,PTGS2的表达主要发生在卵巢静止期的卵泡颗粒细胞和卵黄膜细胞中。
这一发现揭示了PTGS2在牦牛卵巢静止期卵泡发育维持中的重要作用。
二、FGF10对HAS2和PTGS2表达的影响1. FGF10基因FGF10是成纤维细胞生长因子(Fibroblast Growth Factor)家族的一个成员。
研究表明,FGF10在哺乳动物的生殖系统中起着重要调控作用。
其在卵巢中的表达可以促进卵泡发育和卵巢激素的合成。
早期研究还表明,FGF10可以影响HAS2和PTGS2的表达,并调节透明质酸和前列腺素在卵巢中的水平。
2. FGF10对HAS2和PTGS2表达的影响研究结果显示,FGF10处理后,牦牛卵巢颗粒细胞中HAS2和PTGS2的表达水平有显著上调的趋势。


mtr-miR-162靶向GNG11对奶牛乳腺上皮细胞增殖和乳蛋白合成相关基因的调控作用mtr-miR-162靶向GNG11对奶牛乳腺上皮细胞增殖和乳蛋白合成相关基因的调控作用乳腺是哺乳动物体内主要负责乳汁分泌的器官,其中的乳腺上皮细胞是产生乳腺分泌物的细胞类型。
乳腺上皮细胞的增殖和乳蛋白的合成对于母婴健康至关重要。
在乳房组织的生长和发育过程中,许多基因和非编码RNA参与了乳腺细胞的调控。
在这些调控中,转录因子GNG11和非编码RNA mtr-miR-162展现了重要的调控作用。
mtr-miR-162是一种小分子RNA,它是由基因组中的miRNA预序列转录产生的。
mtr-miR-162已经在多种物种中被广泛研究并发现在调控细胞增殖和分化中起到关键作用。
研究发现,mtr-miR-162与GNG11基因存在靶向关系。
GNG11是G蛋白素家族的成员之一,该家族在多种信号转导通路中发挥着重要作用。
而mtr-miR-162能够通过靶向GNG11基因影响乳腺细胞的增殖和乳蛋白合成。
研究表明,mtr-miR-162的表达水平在乳腺细胞增殖和乳蛋白合成中起到关键作用。
当mtr-miR-162的表达水平增加时,GNG11基因的表达受到抑制,从而降低了乳腺细胞的增殖能力。
与此同时,mtr-miR-162的高表达水平还可抑制乳蛋白合成相关基因的表达,从而影响乳腺细胞合成乳蛋白的能力。
进一步研究发现,mtr-miR-162通过靶向GNG11基因下游调控了多个信号转导通路,包括PI3K/AKT、MAPK和Wnt等。
这些信号转导通路在细胞增殖和分化中起到关键作用。
通过对乳腺上皮细胞中这些通路的调控,mtr-miR-162能够影响细胞的增殖和乳蛋白合成。
此外,研究还发现mtr-miR-162的表达受到多种调控因子的调节。
例如,在乳腺生长期间,雌激素和雌激素受体能够增加mtr-miR-162的表达水平。
这种调控可以通过影响mtr-miR-162与GNG11基因之间的靶向关系,进而调节乳腺上皮细胞的增殖和乳蛋白合成。

淫黄葛合剂对APP/PS1 HAMP −/−小鼠认知功能和铁死亡氨基酸代谢通路的影响*刘珊, 贺小平, 赵炎, 钟健民, 张烨华, 刘怡铭, 李佳璇, 董贤慧△(河北中医药大学 河北省心脑血管病中医药防治研究重点实验室,河北 石家庄 050091)[摘要] 目的:探究淫羊藿苷-黄芪甲苷-葛根素(淫黄葛)合剂(YHG )对铁调素(hepcidin , HAMP )基因敲除的APPswe/PS1dE9(APP/PS1 HAMP −/−)小鼠认知功能及铁死亡氨基酸代谢通路的影响。
方法:实验分为阴性对照(C57BL/6小鼠)组、APP/PS1组、APP/PS1 HAMP −/−组、APP/PS1+YHG 组、APP/PS1 HAMP −/−+YHG 组、APP/PS1+地拉罗司(DFX )组和APP/PS1 HAMP −/−+DFX 组,每组6只。
YHG (淫羊藿苷、黄芪甲苷和葛根素的剂量分别为120、80和80 mg/kg )和DFX (100 mg/kg )灌胃给药,C57组、APP/PS1组和APP/PS1 HAMP −/−组小鼠给予等体积蒸馏水灌胃,每日一次,用药2个月。
采用组织铁试剂盒检测小鼠脑组织铁离子含量;透射电镜检测小鼠海马神经元线粒体形态学变化;Morris 水迷宫检测小鼠学习记忆能力;免疫荧光法检测各组小鼠脑组织神经元核抗原(NeuN )的含量;生化试剂盒检测小鼠脑组织谷胱甘肽(GSH )表达情况;Western blot 检测小鼠脑组织谷氨酸半胱氨酸连接酶催化亚基(GCLC )和谷氨酰胺酶2(GLS2)的表达情况。
结果:与C57BL/6小鼠相比,APP/PS1小鼠脑铁含量显著升高(P <0.01),线粒体形态破坏严重,学习记忆能力显著下降(P <0.05),脑组织神经元损伤严重(P <0.01),GSH 、GCLC 和GLS2表达水平均显著降低(P <0.01);与APP/PS1小鼠相比,APP/PS1 HAMP −/−小鼠脑铁含量显著升高(P <0.01),线粒体形态破坏严重,学习记忆能力显著下降(P <0.05),脑组织神经元损伤严重(P <0.01),GSH 、GCLC 和GLS2表达水平显著降低(P <0.05);使用YHG 和DFX 后各组小鼠脑铁含量显著降低(P <0.01),线粒体形态破坏减轻,学习记忆能力显著升高(P <0.05),脑组织神经元损伤减轻(P <0.01),GSH 、GCLC 和GLS2表达水平显著升高(P <0.05)。

哺乳动物生殖调控机制的分子基础哺乳动物的繁衍繁殖是保证物种存在和延续的关键因素。
如何控制哺乳动物的繁殖能力,维持繁殖的正常周期和适应外界环境的变化等,一直是发育生物学研究的重点之一。
调控机制在维持哺乳动物繁殖系统正常功能中发挥着重要的作用,随着生物学技术和方法的不断进步,哺乳动物生殖调控机制的分子基础逐渐被揭示。
一、性激素及其受体的作用内分泌系统是哺乳动物繁殖调控的重要系统之一,其中性激素是最为重要的调节因子之一。
性激素受体及其下游分子在繁殖调控机制中发挥着重要的调节作用。
雌激素对女性繁殖周期相关结构和功能的调节至关重要,例如雌性成熟、产卵和妊娠等生成器官中的雌激素受体Alpha(ESR1)在这个过程中发挥作用。
雄性是由睾酮激素调节的,睾酮等睾酮激素家族成员也是哺乳动物繁殖周期调节的关键分子。
雌激素和睾酮激素的作用通过两种受体进行介导,分别是雌激素受体和睾酮受体。
这两种激素对其对应受体的结合激活,调节下游信号通路,从而发挥不同的生物学作用。
二、下丘脑-垂体-性腺轴的作用下丘脑-垂体-性腺轴(HPG轴)是哺乳动物繁殖周期调控的另一个重要机制。
HPG轴通过下丘脑神经元释放促性腺激素释放激素(GnRH),刺激垂体前叶合成和分泌促性腺激素(FSH)和促黄体生成激素(LH),这些激素在性腺中引发性激素的生成和释放。
性激素作为激素信号的重要组成部分,对不同类型的细胞系表现出不同的生物学效应,从而调节整个生殖系统的功能。
三、neurokinin B系统的作用neurokinin B (NKB)受体被认为是控制GnRH释放的神经递质。
NKB被称为调节HPG轴的第三种调节作用物,同时也与性激素的调节机制有可重叠的作用。
在哺乳动物繁殖周期调节机制中,NKB通过GnRH神经元及其下游信号传递分子活性转化,影响性腺的功能。
在干细胞和上皮细胞中,NKB通常通过激活Kisspeptin 受体另一种神经递质,从而影响下丘脑神经元的表达和功能。
广东药科大学学报Journal of Guangdong Pharmaceutical University Mar.2024,40(2)基于STING/NF-κB信号通路探讨茯苓酸对去卵巢小鼠骨质疏松的作用黄成硕1,陈驹2,吴晓静3,郑锦畅1,梁振1,郭伟雄1,陈继铭1(1.广东医科大学附属医院骨科中心,广东湛江524001;2.广东医科大学附属医院药学部,广东湛江524001;3.广东医科大学附属医院培训部,广东湛江524001)摘要:目的评价茯苓酸通过调控STING/NF-κB信号通路对去卵巢小鼠骨质疏松的影响。
方法取40只SPF级雌,10µg/kg)和茯苓酸组(FL,5mg/kg),性C57BL/6小鼠随机分为假手术组(Sham)、模型组(OVX)、17β-雌二醇组(E2通过切除双侧卵巢建立小鼠骨质疏松模型。
8周实验结束后分别检测血清碱性磷酸酶(ALP)和抗酒石酸酸性磷酸酶(StrACP)水平,观察股骨远端形态学和胫骨(皮质骨)骨形成动态变化,测定右侧股骨的生物力学和STING/NF-κB通路关键蛋白表达水平。
结果与OVX组比较,E组和FL组小鼠血清StrACP和ALP水平明显降低(P<0.01),2股骨远端骨小梁面积百分数明显增加(P<0.01),皮质骨双荧光间距明显减少(P<0.01),股骨最大载荷和断裂载荷明显升高(P<0.01),股骨STING和p-NF-κB p65/NF-κB p65蛋白表达水平明显降低(P<0.01)。
结论茯苓酸通过调控STING/NF-κB信号通路抑制骨吸收和促进骨形成,从而有效减少去卵巢小鼠骨丢失。
关键词:茯苓酸;小鼠;卵巢摘除;骨质疏松;17β-雌二醇中图分类号:R285.5文献标识码:A文章编号:2096-3653(2024)02-0119-05DOI:10.16809/ki.2096-3653.2023122001Effect of pachymoic acid on osteoporosis in ovariectomized mice based on STING/NF-κB signaling pathwayHUANG Chengshuo1,CHEN Ju2,WU Xiaojing3,ZHENG Jinchang1,LIANG Zhen1,GUO Weixiong1,CHEN Jiming1*(1.Orthopaedic Center,Affiliated Hospital of Guangdong Medical University,Zhanjiang524001,China;2.Department of Pharmacy,Affiliated Hospital of Guangdong Medical University,Zhanjiang524001,China;3.Training department,Affiliated Hospital of Guangdong Medical University,Zhanjiang524001,China)*Corresponding author Email:**************Abstract:Objective To evaluate the effect of pachymoic acid on osteoporosis in ovariectomized mice by regulatingSTINGNF-κB signaling pathway.Methods Forty SPF female C57BL/6mice were divided into sham operation,10µg/kg)and pachymoic acid group(FL,5mg/kg),and group(Sham),model group(OVX),17β-estradiol group(E2the osteoporosis model of mice was established by bilateral ovariectomy.After8weeks,the levels of serum alkalinephosphatase(ALP)and tartrate-resistant acid phosphatase(StrACP)were detected,the morphology of distal femurand the dynamic changes of tibia(cortical bone)bone formation were observed,and the biomechanics of the rightfemur and the expression levels of key proteins in STING/NF-κB pathway were determined.Results Compared withgroup and FL group were significantly lowered(P<0.01),the OVX group,the levels of serum StrACP and ALP in E2area percentage of trabecular bone in distal femur was increased(P<0.01),and the double-fluorescence distancebetween cortical bones was decreased(P<0.01).The maximum load and fracture load of femur were significantlyelevated(P<0.01),and the expression levels of STING and p-NF-κB p65/NF-κB p65protein in femur weresignificantly increased(P<0.01).Conclusion Pachymoic acid can inhibit bone resorption and promote boneformation by regulating STING/NF-κB signaling pathway,thereby effectively reducing bone loss in ovariectomized mice.Key words:pachymoic acid;mice;ovariectomization;osteoporosis;17β-estradiol收稿日期:2023-12-20基金项目:广东省医学科学技术研究基金项目(B2021148);2021年度广东省科技专项资金(“大专项+任务清单”)竞争性分配项目(2021A05234);湛江市非资助课题(2020B01275)作者简介:黄成硕(1986-),男,副主任医师,主要从事骨科疾病诊治,Email:*****************通信作者:陈继铭(1980-),男,主任医师,主要从事骨科疾病诊治,Email:**************。
猪GSTM 2基因的多态性及与生产性状相关性分析收稿日期:2010-04-26基金项目:“十一五”国家科技支撑计划重点项目(2008BADB2B02)作者简介:王嘉博(1985-),男,硕士,研究方向为动物遗传育种与繁殖的分子生物学研究。
E-mail:wangjiaboyifeng@ *通讯作者:刘娣,教授,博士生导师,研究方向为猪遗传繁育及家畜生物技术。
E-mail :liudi1963@王嘉博1,2,祝继原2,俄广鑫2,杨少成2,刘娣1,2*(1.东北农业大学动物科学技术学院,哈尔滨150030;2.黑龙江省农业科学院,哈尔滨150086)摘要:母猪产仔性状是评价猪场效益的重要指标,也是猪育种工作的重点。
谷胱甘肽硫转移酶mu 亚基(Glutathione S-transferase Mu2,GSTM2)基因是与母猪生殖系统免疫和生长发育有关的基因。
研究采用PCR-SSCP 的方法分析了GSTM 2基因的单核苷酸多态性,并分析了每个多态位点对母猪繁殖性状的影响。
结果表明,G2、G5、G7位点对母猪总产仔数和活产仔数影响显著,这三个位点都是AB 基因型个体的总产仔数平均数最高,进一步验证了杂合基因型高于纯合基因型性状的理论。
这三个位点对断奶窝平均重影响不显著。
G2位点对窝平均初生重影响显著。
实现了对GSTM2基因多态性的分析,并与母猪产仔性状和初生窝重等生产性状进行了相关性的分析。
得到了基因与性状之间的第一手材料。
关键词:猪;GSTM 2基因;SSCP ;生产性状中图分类号:S828文献标志码:A文章编号:1005-9369(2011)06-0023-05Polymorphism of porcine GSTM 2gene and its correlation analysis with production traits/WANG Jiabo 1,2,ZHU Jiyuan 2,E Guangxin 2,YANG Shaocheng 2,LIU Di 1,2(1.第42卷第6期东北农业大学学报42(6):23~272011年6月Journal of Northeast Agricultural UniversityJune 2011[2]余秋波,王应雄.细胞粘附分子与胚胎着床[J].中国优生与遗传杂志,2005(1):6-8.[3]Suzuki N,Zara J,Sato T,et al.A novel cytoplasmic protein,bystin,interacts with trophinin,tastin and cytokeratin,and may be involved in trophinin mediated cell adhesion between trophoblast and endometrial epithelial cells[J].Proc Natl Acad Sci USA,1998,95:5027-5032.[4]Aoki R,Fukuda M N.Recent molecular approaches to elucidate the mechanism of embryo implantation:Trophinin,bystin,and tastin as molecules involved in the initial attachment of blastocysts to the uterus in humans[J].Semin Reprod Med,2000,18:265-271.[5]Kimber S J,Spanswick C.Blastocyst implantation:the adhesion cascade[J].Semin Cell Dev Biol,2000,11(2):77-92.[6]Suzuki N,Nakayama J,Shih I M,et al.Expression of trophinin,tastin,and bystin by trophoblast and endometrial cells in human placenta[J].Biol Reprod,1999,60:621-627.[7]Nadano D,Sugihara K,Paria B C,et al.Significant differencesbetween mouse and human trophinins are revealed by their expression patterns and targeted disruption of mouse trophinin gene[J].Biol Reprod,2002,66:313-321.[8]Fukuda M N,Miyoshi M,Nadano D.The role of bystin in embryo implantation and in ribosomal biogenesis[J].Cell Mol Life Sci,2008,65(1):92-99.[9]Aoki R,Suzuki N,Paria B C,et al.The BYSL gene product,bystin,is essential for survival of mouse embryos[J].FEBS Lett,2006,580:6062-6068.[10]Adachi K,Soeta-Saneyoshi C,Sagara H,et al.Crucial role of BYSL in mammalian preimplantation development as an integral factor for 40S ribosome biogenesis[J].Mol Cell Biol,2007,27:2202-2214.[11]肖丽娟,杨增明.Trophinin,tastin 和bystin 在哺乳动物胚胎着床及早期胎盘形成中的作用[J].生殖医学杂志,2002,11(2):120-123.[12]许振英.中国地方猪种种质特性[M].杭州:浙江科学技术出版社,1989:16-38.东北农业大学学报第42卷谷胱甘肽硫转移酶(Glutathione S-transferases ;简称GSTs)是生物体内一种重要的一相解毒酶,GST mu 2基因属于GSTs 的mu 亚型,mu 亚型在消除自由基、过氧化物、亲电子基团和中间数具有酶功能,并参与细胞保护和调节细胞生长,GSTM2具有前列腺素E 合成酶(PGES)的活性,可以参与前列腺素E2(PGE2)的合成[1]。
哺乳动物生殖遗传过程中的rna调控及染色
质重塑学术报告
近年来,对于哺乳动物生殖遗传过程中RNA调控及染色质重塑的研究日渐深入。
生命的起源和特性是由基因和其表达调控来决定的。
RNA在哺乳动物生殖中起着相当重要的调控作用。
它们能够调节基因的表达,并参与到生殖细胞形成、发育、成熟和精子与卵子结合等过程中。
在哺乳动物生殖过程中,RNA调控更为重要的是在染色质重塑中所起到的作用。
染色质重塑是指在DNA中添加或去除成对的化学链,以此来调节基因的表达状态。
RNA可以通过各种途径调控染色质重塑,例如:通过酶促反应移除或添加DNA上的甲基化。
RNA调控还通过影响蛋白质的组装来影响染色质重塑,从而调控胚胎发育和与疾病相关的基因。
值得注意的是,RNA调控在新兴的基因编辑技术中也起到了关键的作用,其技术被应用于疾病治疗和农业领域中。
由于RNA分子在哺乳动物生殖中的关键作用,它们是非常有前景的生物工程领域资料。
这表明生物学家研究RNA调控显著的作用在于推动基于RNA技术的治疗及农业生产。
总之,RNA调控在哺乳动物生殖遗传过程中及其他生物领域中扮演着非常重要的作用,其研究的深入将会有利于生物技术的进步以及对哺乳动物产生的保护和进一步发展。
二、预期目标1、本项目的总体目标通过系统而创新性地研究肾脏发育过程中三个重要环节(后肾间充质和输尿管芽的相互作用、帽状间充质细胞分化为上皮细胞以及肾单位的形成)所涉及的关键分子的表达变化、功能作用及其主要分子信号通路,阐明哺乳动物后肾发育过程中细胞定向分化的分子调控机制。
建立和完善肾脏输尿管发育异常和遗传性肾病患者及家系成员的临床信息及生物样本资源库,开展相关疾病的遗传分子诊断。
提出并验证急性肾损伤修复再生的过程是肾脏发育某些过程的重现的学术观点,成功应用肾脏发育关键因子促进损伤肾脏的修复再生。
建立健全我国肾脏发育生物学研究的科研基地和技术平台,在肾脏发育生物学研究的国际前沿,取得一批具有自主知识产权的研究成果,提升我国在肾脏发育生物学研究领域的国际学术地位。
建立、培养、锻炼一支从事肾脏发育生物学基础和应用研究的高素质、高水平的学术团队,培养一批热爱肾脏发育生物学研究并具有一定专业技术水平的博士后、博士和硕士研究生等后备队伍。
2、五年预期目标(1)阐明Robo2基因变异导致肾输尿管发育异常、Cox2基因敲除导致表浅肾单位成熟障碍的分子机制。
(2)明确肾脏发育关键分子及主要信号通路在肾脏发育重要环节细胞定向分化过程中新的作用机制。
(3)构建后肾帽状间充质细胞分化为肾脏上皮细胞重要环节不同部位不同分化命运的细胞文库、基因表达谱和关键调控因子的分子解剖图谱。
(4)发现肾发育异常新的致病基因或基因突变位点;开展肾发育异常及遗传性肾病的遗传分子诊断。
(5)提出并验证急性肾损伤修复再生的过程是肾脏发育某些过程的重现的学术观点,应用肾脏发育关键因子促进损伤肾脏的修复再生。
(6)建立胚肾发育特异性表达GFP的转基因品系以及靶器官特异性Cre/loxP、条件性敲除靶基因的动物模型及其后续研究的技术平台。
(7)建立健全肾发育异常及遗传性肾病患者的临床信息及生物样本资源库。
(8)发表高水平的SCI收录论文40篇以上,申请专利2项以上。
(9)锻炼和造就一支从事肾脏发育生物学研究的高素质、高水平的学术团队,培养一批博士后、博士和硕士研究生。
(10)提升我国肾脏发育生物学领域的国际学术地位。
三、研究方案1、学术思路在肾脏的发育过程中,输尿管芽和后肾间充质的相互作用、间充质细胞分化为上皮细胞、肾单位的形成是肾脏发育的三个重要环节。
细胞定向分化是肾脏发育的核心。
本项目需要解决的关键科学问题是:在肾脏发育的重要环节有哪些关键调控因子影响了细胞的定向分化?这些分子的信号通路和调控机制如何?肾脏发育关键因子对先天性肾脏输尿管发育异常的分子诊断和损伤肾脏的修复再生有什么作用?为了解决这些问题,本项目结合发育与生殖研究国家重大科学研究计划2010年度重要支持方向“针对哺乳动物的一种重要组织器官,如肾脏、肝脏、造血系统等,在分子遗传水平上研究发现控制重要组织器官发育的关键因子和信号通路,揭示组织器官发育的根本机制”的要求,确定本项目的学术思路(见下图):以肾脏发育三个重要环节为切入点,在分子遗传水平上,重点研究哺乳动物后肾发育过程中细胞定向分化的关键因子和信号通路,揭示肾脏发育的根本机制,将胚肾发育关键因子应用于肾脏输尿管发育异常和遗传性肾病的分子诊断,促进损伤肾脏的修复再生。
2、技术路线本项目用于寻找和探索新基因功能和机制,普遍使用的技术途径是:筛选候选基因----验证其表达变化----干预实验----明确其功能和机制----应用研究。
(1)应用转基因鼠和细胞特异条件性靶基因敲除鼠,结合基因组学、比较生物信息学研究技术,筛选肾脏发育关键候选基因。
(2)用分子生物学验证已有较好前期研究基础和新的候选基因在特定细胞、组织的时空表达特征。
(3)利用siRNA干预、质粒转染、转基因、基因敲除等基因修饰以及抗体中和、受(配)体拮抗等方法,进行体内、外的干预实验。
(4)明确关键候选基因和蛋白的功能和分子作用机制。
(5)将基础研究获得的肾脏发育关键因子应用于肾输尿管发育异常及遗传性肾病的分子诊断和损伤肾脏的修复再生。
3、创新点与特色(1)阐明肾脏发育重要环节关键因子在细胞定向分化过程中新的作用及分子调控机制,为培育可供移植的肾脏器官奠定基础。
(2)构建后肾发育重要环节不同部位、不同分化命运的肾脏细胞文库以及能反映后肾发育重要环节关键分子时空表达特征的分子解剖图谱。
创建胚胎肾脏、成熟肾脏、肾发育异常及遗传性肾病模型的肾脏资源库。
(3)建立肾发育异常及遗传性肾病患者的样本资源库及信息网络平台,发现并验证新的致病基因,建立我国肾发育异常及遗传性肾病的分子诊断体系。
(4)提出并验证肾损伤修复再生的过程是肾脏发育某些过程的重现的学术观点,应用新的肾脏发育关键因子的整合促进损伤肾脏的修复再生。
(5)主要特色是:针对遗传性肾脏疾病、肾输尿管发育异常及急性肾损伤多见、危害大的临床现象,将肾脏发育生物学基础研究的成果直接转化为遗传性肾病和肾发育异常的分子诊断及促进急性肾损伤的修复再生,“出口”明确,研究实用。
4. 可行性分析(1)具有坚实的理论基础本项目以肾脏发育过程中后肾间充质和输尿管芽的相互作用、帽状间充质细胞分化为上皮细胞以及肾单位的形成三个重要环节为切入点,重点研究影响细胞定向分化的关键因子和主要信号通路的调控机制,在理论上是有道理的。
因为胚肾发育需要经历这些必不可少的环节,后肾间充质细胞发育成肾单位需要细胞不断地定向分化为肾脏不同的固有细胞,细胞定向分化是肾脏发育的核心。
在肾脏发育的重要环节、围绕肾脏发育的核心问题,分别以Robo2、Cox2、Wnt、Notch、Wt1、VEGF等信号分子作为研究的重点是有依据的。
Robo2和Cox2拥有较好的前期研究基础,Wnt信号通路在后肾间充质上皮转分化(MET)中具有重要的作用,Notch信号调控肾单位的节段形成,Wt1决定足细胞的命运,VEGF是内皮细胞募集血管球形成所必需的,分别在肾脏发育的不同环节具有非常重要的调控作用。
(2)具有较好的前期研究基础本项目组有专门从事转基因动物和基因敲除动物模型研究的课题组成员和单位,主要学术骨干均在国外从事多年的分子细胞生物学基础实验研究,熟练掌握项目研究所需要的方法和技术。
包括利用细胞特异性Cre/loxP转基因鼠,进行特异性靶器官基因敲除、显微胚胎肾分离、体外胚肾组织和细胞培养的关键技术等。
积累了宝贵的研究经验,建立了一套完善的研究方法。
在急性肾小管坏死与再生修复的研究方面,在世界上首先提出骨调素及其受体CD44与肾小管坏死后上皮细胞的再生以及上皮细胞的极性形成有关,得到国际学术界的肯定,论文发表在国际肾脏病杂志(Kidney Int)和美国肾脏病杂志(Am J Nephrol)。
近年来有关急性肾小管坏死后上皮细胞再生方面的研究,连续得到国家自然科学基金的资助。
(3)拥有良好的设备和成熟的技术本项目的课题承担单位均具有很强的科研实力。
本单位开放的国家重点实验室以及各学科自身的实验室,已具备完成本项目所需要的设备和技术条件,是完成本项目的硬件保证。
(4)富有活力和团结拼搏精神的科研人才队伍本项目的研究队伍主要由长期从事肾脏及发育相关研究的学科带头人和中青年学术骨干组成,均工作在科研的第一线,主要学术骨干多有国外留学的经历,在本领域发表过有影响的SCI论文,他们有理论、有实践经验、有热情、有上进心和科研动力,富有活力和团结拼搏精神,与国外有关研究机构和研究人员保持着良好的关系,是完成本项目的人才保障。
人才队伍包括从事发育生物学、遗传学、基因工程学、再生医学、分子生物学、细胞生物学、肾脏病理学、病理生理学、肾脏病学的专职科研人员和研究生队伍。
项目和课题负责人思路清晰、作风严谨,有很强的协调能力。
尤其重要的是本项目的幕后有一批关心、支持、帮助本项目立项和实施的专家队伍,他们在精神上的鼓励、学术上的指导、技术上的点拨,是本项目成功实施的重要保障。
5、课题设计为了解决本项目的关键科学问题,结合发育与生殖研究国家重大科学研究计划2010年度重要支持方向的要求,本项目以肾脏发育过程中的三个重要环节细胞定向分化的关键因子和信号通路及其在分子诊断和修复再生中的应用为切入点,设计了四个课题:(1)肾脏发育过程后肾间充质和输尿管芽相互作用主控基因的调控研究。
(2)胚肾帽状间充质细胞分化为上皮细胞的关键因子和信号通路。
(3)肾血管球形成和足细胞发育的分子调控机制。
(4)肾脏发育关键因子在分子诊断及修复再生中的作用。
肾脏发育是连续的,重要环节的划分和课题的设计是人为的。
本项目以肾脏发育过程中的重要环节为主线设计课题。
由于肾脏发育的各个环节不是孤立的,而是相互联系的,因此课题间的研究内容也是相互联系的。
如第一课题输尿管芽和后肾间充质的相互诱导贯穿肾脏发育的全过程;第二课题帽状间充质细胞定向分化为肾脏上皮细胞和第三课题肾单位的形成均受到第一课题输尿管芽和后肾间充质相互作用的影响。
四个课题涉及的关键调控因子和信号通路也不是孤立的,如Wnt分子及信号通路可能与四个课题的研究内容都有关系。
因此一个环节科学问题(关键因子和信号通路)的解决,对另一环节的研究和认识可以提供借鉴和参考。
另外,一、二、三课题发现的肾脏发育关键因子可以应用于第四课题的肾脏发育异常的分子诊断和肾损伤的修复再生,而第四课题不仅可以应用和验证一、二、三课题的结果,其自身的研究成果对前三个课题也有帮助。
四个课题总的目标是一致的,就是发现并阐明肾脏发育重要环节中细胞定向分化的关键调控因子和主要信号通路,揭示肾脏发育的根本机制,将胚肾发育关键因子应用于肾脏输尿管发育异常和遗传性肾病的分子诊断,促进肾损伤的修复再生。
课题间的有机联系见下图:第一课题肾脏发育过程中后肾间充质和输尿管芽相互作用主控基因的调控研究研究目标1.阐明Robo2条件性敲除,通过后肾间充质和输尿管芽的相互作用导致肾输尿管发育异常的分子机制及其与主控基因GDNF/Ret的内在联系。
2.明确COX2基因敲除导致肾被膜下表浅肾单位障碍的分子机制,探讨COX2在浅表肾单位后间充质和输尿管芽相互作用的意义。
3.构建肾脏输尿管发育异常的动物模型,为深入研究肾脏输尿管发育过程提供良好的技术平台。
研究内容1.利用转基因小鼠和不同时间靶器官特异性Cre/loxP小鼠,进行靶器官细胞特异性敲除Robo2,结合基因组学技术,观察分析不同时期、特定部位(如后肾间充质和输尿管芽)基因的表达谱差异及表型变化,筛选和验证候选关键调控基因,然后进行功能和机制研究(包括与GDNF/Ret的关联),明确Robo2基因变异导致多个输尿管芽和多个肾形成、输尿管发育异常以及肾小球数目减少的机制。
2.应用Cox2绿色荧光报告鼠和条件性Cox2基因敲除鼠,观察Cox2的时空表达特征,研究Cox2对GDNF/Ret等主控基因的影响,明确Cox2对肾脏皮质肾单位(肾小球、肾小管)发育成熟的影响及其分子机制。