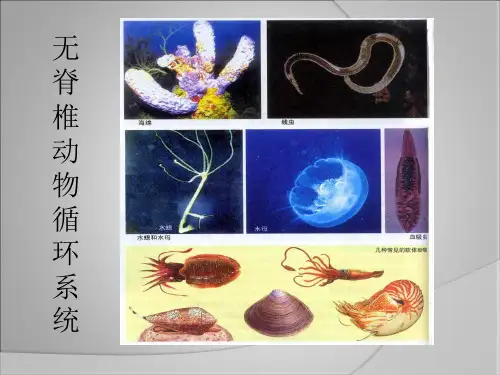
无脊椎动物各系统进化主线 3
- 格式:doc
- 大小:583.50 KB
- 文档页数:12



蚯蚓主要系统的进化趋势
蚯蚓是一种无脊椎动物,属于环节动物门,蚯蚓的进化趋势主要体现在以下几个系统上:
1.消化系统:蚯蚓的消化系统逐渐发展为具有特殊化学功能的肠道。
它们的食物经过胃和肠道的消化,通过分泌酶类和细菌的共同作用,将有机物质分解为肥料。
2.循环系统:蚯蚓的循环系统是开放式的,血液不通过血管流动,而是通过血窦散布到身体各处。
这种系统对于蚯蚓来说是非常有效的,它可以更好地满足身体各部分的氧气和养分需求。
3.呼吸系统:蚯蚓没有专门的呼吸器官,它们通过皮肤进行氧气和二氧化碳的交换。
蚯蚓的皮肤防水层很薄,可以通过体表呼吸来获取氧气。
4.神经系统:蚯蚓的神经系统相对简单,由位于头部的脑和贯穿体节的神经索组成。
蚯蚓具有感觉器官,如触须和感光细胞,它们能感知环境变化,并做出相应的反应。
5.生殖系统:蚯蚓的生殖系统是雌雄异体的,包括生殖腺和生殖道。
蚯蚓是通过交配来繁殖的,雌蚯蚓在交配后会产卵,经过一段时间后孵化出幼虫。
总体来说,蚯蚓的进化趋势是适应生活在土壤中的环境,它们具有高效的消化功
能,能够利用有机物质为自身提供养分。
另外,蚯蚓通过皮肤呼吸和具有开放循环系统的特点,适应了土壤中的氧气获取和呼吸需求。
蚯蚓的神经系统和生殖系统也在适应土壤中的生活方式方面发生了相应的适应性进化。




---------------------------------------------------------------最新资料推荐------------------------------------------------------物发生律或称重演律物发生律或称重演律:德国学者赫克尔提出生物发展史可分为两个相互密切联系的部分,即个体发育和系统发展,也就是个体的发育历史和由同一起源所产生的生物群的发展历史。
个体发育史是系统发展史的简单而迅速的重演。
消化系统的进化主线:原生动物只有胞内消化,可用伪足或胞口摄食,另外还可植食和腐食性;海绵动物仍然是胞内消化;腔肠动物开始有了消化管;胞内和胞外消化;扁形动物为胞外消化,但消化管是不完全的;线形动物出现了完全的消化管,并且有了分化;环节动物以后由于真体腔的出现,消化管更加复杂和分化,同时有了消化腺。
呼吸系统的进化主线:原生动物、海绵动物、腔肠动物都没有呼吸和排泄系统,呼吸作用通过体表完成的;扁形动物和线形动物也无呼吸系统,呼吸也是体表进行的,寄生种类为厌氧呼吸,环节动物的呼吸可通过体表和疣足进行;软体动物的呼吸通过体壁突起的鳃和外套膜进行;节肢动物的呼吸器官包括鳃(虾)、书鳃(鲎)、书肺(蜘蛛)、气管(昆虫)、气管鳃(幼虫) 以及体表;棘皮动物的呼吸是通过管足和皮鳃完成。
1 / 11排泄系统的进化主线:原生动物、海绵动物、腔肠动物的排泄活动也是借体表完成的;原生动物还可通过伸缩泡进行排泄;扁形动物和线形动物的排泄系统为外胚层内陷形成的原肾;扁形动物的排泄系统是焰细胞,线形动物则是原肾管;环节动物的排泄系统是由外胚层和中胚层共同组成的混合型的后肾;软体动物的排泄系统是中胚层的后肾;节肢动物排泄系统有两类,一是体腔管演化而来的肾管,一是马氏管;棘皮动物的排泄是通过管足和皮鳃完成。
循环系统的进化主线:环节动物之前的各门类没有专门的循环系统;原生动物中的细胞质流动起到循环的作用;海绵动物、腔肠动物和扁形动物通过消化循环腔起着循环的作用;线形动物的原体腔也有输送养料的功能;真体腔的出现产生了血管,环节动物开始有了真正的循环系统;除环节动物中的大部分为闭管系统外,其他的高等无脊椎动物的循环系统均为开管式。

一、体制:无对称→球形对称→辐射对称→两侧对称(1)无脊椎动物原生动物:变形虫——无对称放射虫、太阳虫、团藻——球形对称(通过一个中心点,有无数对称轴,可将球体切成相等的对称面)→适应于悬浮在水中草履虫——两侧对称多孔动物、腔肠动物:基本上为辐射对称(通过身体中央轴有许多切面可以把身体分成相等的部分)→适应于固着在水中海葵——两辐对称(海葵由于有口、口道沟的存在,身体只能通过体轴作平行与垂直口道沟的两个对称面)扁形动物、线形动物、环节动物、软体动物、节肢动物、棘皮动物:生活方式从固着、漂浮演化成爬行方式或游泳,身体呈两侧对称→适应于爬行生活,是动物由水生进化到陆生的重要条件之一。
二、胎层:单细胞→单细胞层→二胚层→三胚层(分化盲支:多孔动物门胚胎发育存在逆转)原生动物:单细胞动物没有胚层的概念;即使是团藻也只有一层细胞,;(真正地多细胞动物有胚层的分化)肠腔动物:二胚层扁形动物、线形动物、环节动物、软体动物、节肢动物、棘皮动物:出现三胚层(在动物进化上有着极为重要的意义)三、体腔:无体腔→假体腔→真体腔(是高等无脊椎动物的重要标志之一)原生动物、多孔动物、腔肠动物、扁形动物:无体腔线形动物(假体腔动物):假体腔(初生体腔,即直接跟体壁的肌肉层和消化管道的壁相接触没有中胚层形成的体腔膜包围,也不和外界相通)←胚胎时期的囊胚腔所形成的环节动物、节肢动物、棘皮动物(软体动物真体腔退化):真体腔(体腔的位置处于中胚层之间,外围由中胚层形成的体腔膜所包围)→造成了各种器官的进一步特化四、体节和身体分布:同律分节→异律分节(身体分节是高等无脊椎动物的重要标志之一)原生动物、多孔动物、腔肠动物:不分节扁形动物、线形动物:原始分节(机体各部分结构和机能分化,但身体不分节)环节动物:同律分节节肢动物、软体动物、棘皮动物:异律分节(导致了动物的身体分部)五、体表和骨骼:细胞膜→细胞外有壳→外有纤毛→有角质层→体外有壳→体外含几丁质原生动物:仅细胞膜(部分植物性鞭毛虫有细胞壁,部分有壳肉足虫具外壳、含角质、石灰质等); 扁形动物:有体表纤毛;线形动物、环节动物:体表有角质层;软体动物:有石灰质壳节肢动物、棘皮动物:有几丁质外壳(骨骼是维持体形的支架,无脊椎动物的骨骼一般由外胚层分化而成,故称外骨骼;但棘皮动物的骨骼是起源于中胚层;软体动物头足类的软骨也是起源于中胚层)六、运动器官和附肢原生动物:鞭毛、伪足和纤毛;多孔动物:鞭毛;腔肠动物:有了原始的肌肉细胞;幼虫以纤毛运动;扁形动物:中胚层形成的肌肉使动物体得以蠕动;体表有纤毛用于运动;寄生种类的幼体有纤毛;线形动物:用体壁纵肌作蛇行运动;环节动物:用肌肉、刚毛和疣足运动;软体动物:用肉质的足作爬行运动;节肢动物:用附肢运动棘皮动物;用腕和管足运动。

物发生律或称重演律:德国学者赫克尔提出生物发展史可分为两个相互密切联系的部分,即个体发育和系统发展,也就是个体的发育历史和由同一起源所产生的生物群的发展历史。
个体发育史是系统发展史的简单而迅速的重演。
消化系统的进化主线:原生动物只有胞内消化,可用伪足或胞口摄食,另外还可植食和腐食性;海绵动物仍然是胞内消化;腔肠动物开始有了消化管;胞内和胞外消化;扁形动物为胞外消化,但消化管是不完全的;线形动物出现了完全的消化管,并且有了分化;环节动物以后由于真体腔的出现,消化管更加复杂和分化,同时有了消化腺。
呼吸系统的进化主线:原生动物、海绵动物、腔肠动物都没有呼吸和排泄系统,呼吸作用通过体表完成的;扁形动物和线形动物也无呼吸系统,呼吸也是体表进行的,寄生种类为厌氧呼吸,环节动物的呼吸可通过体表和疣足进行;软体动物的呼吸通过体壁突起的鳃和外套膜进行;节肢动物的呼吸器官包括鳃<虾>、书鳃<鲎>、书肺<蜘蛛>、气管<昆虫>、气管鳃<幼虫> 以及体表;棘皮动物的呼吸是通过管足和皮鳃完成。
排泄系统的进化主线:原生动物、海绵动物、腔肠动物的排泄活动也是借体表完成的;原生动物还可通过伸缩泡进行排泄;扁形动物和线形动物的排泄系统为外胚层内陷形成的原肾;扁形动物的排泄系统是焰细胞,线形动物则是原肾管;环节动物的排泄系统是由外胚层和中胚层共同组成的混合型的后肾;软体动物的排泄系统是中胚层的后肾;节肢动物排泄系统有两类,一是体腔管演化而来的肾管,一是马氏管;棘皮动物的排泄是通过管足和皮鳃完成。
循环系统的进化主线:环节动物之前的各门类没有专门的循环系统;原生动物中的细胞质流动起到循环的作用;海绵动物、腔肠动物和扁形动物通过消化循环腔起着循环的作用;线形动物的原体腔也有输送养料的功能;真体腔的出现产生了血管,环节动物开始有了真正的循环系统;除环节动物中的大部分为闭管系统外,其他的高等无脊椎动物的循环系统均为开管式。
第十二章无脊椎动物各门类之间的系统演化关系一、原生动物起源:孢子虫全部寄生,纤毛动物大多结构比较复杂,肉足动物虽然形态简单,但又行异养生活,可见它们均不是最早出现的,只有鞭毛虫中的虫体,尤其是结构比较简单的无色鞭毛虫,尤其是那些能行腐生性的营养的无色鞭毛虫,它们可能经漫长岁月、发展变化成至今的原生动物。
二、多孔动物起源:多孔动物具大量的领细胞,其结构与原生动物的群体领鞭毛虫(Choanoflagellates)与原绵虫(Proyterospongia)十分的相似,因此多孔动物可能是起源于原生动物,可能是由类似群体领鞭毛虫发展而来的。
由于多孔动物具有特殊的水沟系与胚层逆转现象等,因此把多孔动物视为是多细胞动物后生动物中的侧生动物。
三、扁盘(吞噬动物)起源:它们形状、大小、运动方式与变形虫很相似,但虫体由几千个细胞组成为两层(Gre ll 1982认为它是两胚层),细胞发生四种分化(上皮细胞具鞭毛上皮细胞腺细胞以及埋于间质层的星状纤维细胞),行动物性营养,具体外消化能力(腹面腺细胞分泌消化酶,营养同时被它们吸收),因此扁盘(吞噬动物)可能是起源原生动物,它应该是属于多细胞动物中最原始的真后生动物。
四、腔肠动物起源:从形态结构(实心原肠胚、外长纤毛)推测,腔肠动物可能是由群体鞭毛虫,经个体(细胞)移入,最终发展成为具两个胚层的动物。
另外从腔肠动物开始,再无胚层逆转现象,因此把腔肠动物以后的动物视为是多细胞动物中真后生动物。
五、栉水母动物起源:从栉水母动物体制(两辐对称)、两胚层、消化循环系统(具辐管)说明,可能栉水母动物是与腔肠动物同起源(同祖先),只是栉水母动物比腔肠动物略高等,因为栉水母动物有了中胚层细胞等。
六、扁形动物起源:看法不一。
一派认为扁形动物起源栉水母动物(栉水母动物中的扁栉水母Ctenoplana,能水底爬行,可能经长期丧失游泳能力),另一派认为起源于腔肠动物,由类似浮浪幼虫祖先经水底爬行,口移向腹面逐渐演化而形成的。