生活史进化
- 格式:doc
- 大小:471.50 KB
- 文档页数:36



人类生活史研究的社会学意义人类生活史是指人类在地球上的进化、演变和发展过程,涉及到人类的生命起源、文明崛起、社会结构、文化传承等方面,是一个综合性的学科领域。
人类生活史的研究不仅可以让我们更加了解人类的过去和现在,还对我们未来的发展和进步有重要的指导意义。
从社会学的角度来看,人类生活史的研究有着深远的社会学意义。
一、人类社会演变的历史脉络人类生活史的研究可以帮助我们了解人类社会的演变历程,发现人类社会结构和文化的演变规律。
人类社会从原始社会到现代社会的演变过程,既是一个宏大历程,也是一个长达数千年的历史脉络。
在这个过程中,人类社会从采集社会、农业社会、工业社会到信息社会,一步步实现了社会、文化、经济和技术的发展升级。
对这个演变历程的研究,可以让我们更好地理解人类社会的现状和未来发展趋势。
二、文化传承的重要意义人类社会不同于其他动物社会,最显著的特征之一就是文化传承。
文化是人类社会的产物,是人类社会与其他动物社会最根本的区别。
人类社会的文化传承涉及到种种基本要素,如语言、信仰、价值观念、习俗和传统等。
人类生活史的研究可以帮助我们了解各个时期的文化传承,不同文化间的相互影响和融合。
有了这样的了解,我们才能更好地认识文化的本质和功能,更好地理解不同民族和文化间的差异。
三、社会结构与社会问题人类社会地位结构从采集社会、农业社会到工业社会,产生了巨大的变化。
社会结构的演变对社会问题有诸多影响。
研究人类社会的历史发展,可以帮助我们更好地认识不同文化传承在社会结构演变中的影响。
同时,对不同社会结构和社会问题的研究,可以为我们找到解决方案提供重要的指导,为社会的和谐发展提供保障。
四、人类和自然环境的关系人类生活史的研究不仅关注人类的社会结构和文化建构,还关注人类和自然环境的关系。
随着人类社会的演进,人类对自然环境的影响也越来越大。
对人类与自然环境的关系的研究,能够帮助我们更好地认识人类文明历程中人与自然环境的关系,分析人类环境问题的成因和现实意义,并支持环境主义的理论和实践。

中国社会生活史课程总结3000字中国社会生活史一般被视为中国社金史的一个组成部分而缺乏对其内容体系进行理论探讨.实际上,中国社会生活史与中国社会史既有区别又有联系,作为研究中国社会不同历史时期各种生活现象及其价值观念发展变化的一门学科,其体系主要应包括生活环境与生活方式、社会群体及其行为特征、家庭生活及衣。
住行变证、社会教育与社会交往、生活的态度与价值观念等五个方面的内容,无论是从社会实践还是从史学理论的角度来看,中国社会生活史都应作为一门独立的学科而受到重视.社会生活史是教育部规定的高等学校文科学生的基本必修课之一,讲述的是广大民众包括物质生活、精神生活、人际关系在内的日常生活和活动。
该课是中国历史文化和民众民俗文化的有机组成部分。
过去的历史研究,往往忽略对历史细部的描述,使历史成为枯燥乏味的数字与事件的罗列,即使对历史规律的表述,也因缺少社会生活的实证而削弱了论据的支撑。
通过本课程所展示的社会生活的具体层面来再现历史上民众生活的多彩画面,使学生对历史的认识除了政权更替、制度消长外还有更加丰富、生动的社会生活,使他们对历史的关注除了精英人物之外,还有更广大的民众及其生活。
对社会生活史的重视将必定使历史更加鲜活,更加趋近于历史的本真。
通过学习,不仅能扩大学生的知识面,而且能使他们加深对祖国历史文化的深刻认识及中国国情的了解,从而进一步提高大学生的人文素养。
理想状态的社会生活史:展现真实、生动的生活画卷;所有那个时代、那个地方与人的生活相关的一切,都应当可以成为我们关注研究、叙述的对象。
回到现实,制约社会生活史研究的主要因素:古代生活的有限遗存决定素材现代史家的史学观念约束视野20 世纪域内中国社会生活史发展三阶段20至40年代为第一阶段,学科初建与兴起。
50至70年代为第二阶段,初露头角的社会生活史研究在中国大陆和台湾呈现不同命运。
大陆学界以马克主义的名义,将社会生活研究视为‘庸俗’“烦琐’而遂之于史学门槛之外。





生活史对策概述大气基地贺园园1111700026生活史与生活史对策:生活史(life history)意为生物从其出生到死亡所经历的全部过程,也叫生活周期(life cycle)。
生活史性状包括出生时个体大小;生长形式;成熟年龄;成熟时个体大小;后代的数量、大小、性比;特定年龄和大小的繁殖投入;特定年龄和大小的死亡规律;寿命等。
生活史对策(life history strategy)是指生物在生存斗争中获得的生存对策,也称生态对策(bionomic strategy )或进化对策,例如生殖对策、取食对策、迁移对策、体型大小对策等主要内容:任何生物做出的任何一种生活史对策,都意味着能量的合理分配,并通过这种能量使用的协调,来促进自身的有效生存和繁殖。
每个生物具有生长、维持生存和繁殖三大基本功能,生物必须采取一定的策略配置能够获得的有限资源,其核心主要强调在特定环境中提高生殖、生存和生长能力的组合方式.1)生长对策●生长速度早期演替种:早期迅速生长,具开拓对策(白桦)后期演替种:早期生长缓慢,具保守对策(红松)●生长方式以温带木本植物为例,其顶枝形成有两种主要方式:①有限生长类型:顶枝在冬季完全定型,冬芽形成时就决定了叶子数目。
②无限生长类型:冬芽只含有少量叶原基,在下一个生长季,顶枝尖端在生长季内还能产生新的叶子和节间。
●根冠比率—物质分配2)生殖对策生殖对策实际就包括两个方面的问题:第一是生殖者存活的问题,也即生殖的代价问题,生物生殖必然带来变化的生理压力和个体危险,因此,也就必然会影响到生物的生存;第二是生殖的效率问题,生物选择的对策,都旨在提高生殖的效率,这一点可从植物的生殖行为中证实。
1)体型效应物种个体的大小与其寿命有很强的正相关关系。
2)成体的存活与繁殖成熟个体存活率低,生物繁殖越早,投资于繁殖的能量越多;成熟个体存活率越高,生物的繁殖期越晚(个体较大),分配于繁殖的能量就越少。
3)当前繁殖与未来繁殖如果未来生命期望低,分配给当前繁殖的能量应该高,而如果剩下的预期寿命很长,分配给当前繁殖的能量应该较低。

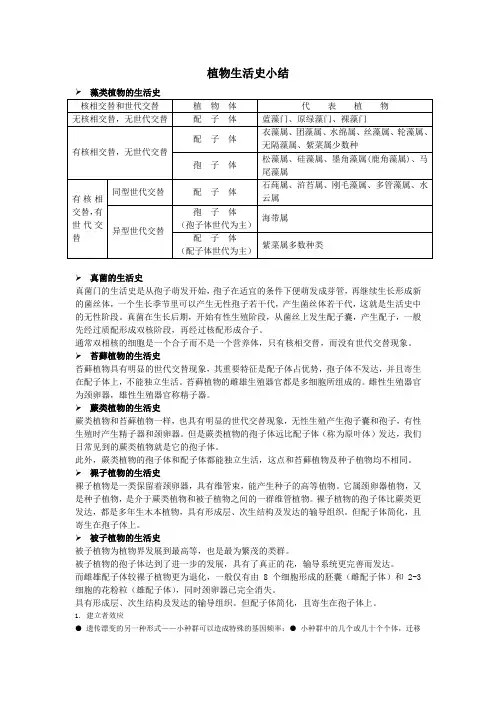
植物生活史小结➢真菌的生活史真菌门的生活史是从孢子萌发开始,孢子在适宜的条件下便萌发成芽管,再继续生长形成新的菌丝体,一个生长季节里可以产生无性孢子若干代,产生菌丝体若干代,这就是生活史中的无性阶段。
真菌在生长后期,开始有性生殖阶段,从菌丝上发生配子囊,产生配子,一般先经过质配形成双核阶段,再经过核配形成合子。
通常双相核的细胞是一个合子而不是一个营养体,只有核相交替,而没有世代交替现象。
➢苔藓植物的生活史苔藓植物具有明显的世代交替现象,其重要特征是配子体占优势,孢子体不发达,并且寄生在配子体上,不能独立生活。
苔藓植物的雌雄生殖器官都是多细胞所组成的。
雌性生殖器官为颈卵器,雄性生殖器官称精子器。
➢蕨类植物的生活史蕨类植物和苔藓植物一样,也具有明显的世代交替现象,无性生殖产生孢子囊和孢子,有性生殖时产生精子器和颈卵器。
但是蕨类植物的孢子体远比配子体(称为原叶体)发达,我们日常见到的蕨类植物就是它的孢子体。
此外,蕨类植物的孢子体和配子体都能独立生活,这点和苔藓植物及种子植物均不相同。
➢裸子植物的生活史裸子植物是一类保留着颈卵器,具有维管束,能产生种子的高等植物。
它属颈卵器植物,又是种子植物,是介于蕨类植物和被子植物之间的一群维管植物。
裸子植物的孢子体比蕨类更发达,都是多年生木本植物,具有形成层、次生结构及发达的输导组织。
但配子体简化,且寄生在孢子体上。
➢被子植物的生活史被子植物为植物界发展到最高等,也是最为繁茂的类群。
被子植物的孢子体达到了进一步的发展,具有了真正的花,输导系统更完善而发达。
而雌雄配子体较裸子植物更为退化,一般仅有由8个细胞形成的胚囊(雌配子体)和2-3细胞的花粉粒(雄配子体),同时颈卵器已完全消失。
具有形成层、次生结构及发达的输导组织。
但配子体简化,且寄生在孢子体上。
1. 建立者效应●遗传漂变的另一种形式——小种群可以造成特殊的基因频率:●小种群中的几个或几十个个体,迁移到它处定居下来,与原种群隔离开来,自行繁殖形成新的种群;有些等位基因没有带出来,导致新种群与原种群的基因频率的差异;●新种群的基因频率取决于建立者(定殖者)——分离出来的几个或几十个个体●意义:通过自然选择,有可能形成新物种。
普通生态学第四章 种群生活史鲑,大麻哈鱼普通生态学•不同种类其生活史类型的变异是巨大的,这些变异是如何进化而来的是生态学的一个关键问题。
•生活史有关繁殖部分是生态学研究中特别活跃的领普通生态学第四章 种群生活史§1 生活史概述§2 繁殖成效§3 性选择§4 繁殖策略普通生态学⏹ 生活史 (life history): 生物从出生到死亡所经历的全部过程。
生态学上,生物的生活史具有多重含义,广义上讲,是指生物的一生中生长发育和繁殖模式。
§1 生活史概述普通生态学生活史的几个方面⏹个体大小 (Body Size)⏹生长和发育 (Growth and development)⏹扩散 (Dispersal)⏹繁殖格局 (Reproductive patterns)⏹性选择 (Sexual selection)实际上, 生活史的有关研究最典型地反映出进化和生态学两门科学是密切联系的。
普通生态学 1.1 个体大小 不同生物个体大小差异非常巨大,从1微米长的细菌到100米高的红杉。
个体的大小是生物生活史的适应特征,最终是选择的产物。
个体大小不同的物种具有各自的优势,是自然选择的结果。
普通生态学个体大小的一般作用规律⏹个体大小与生活周期的长短有很好的相关性;⏹生命周期短的生物通常更快的适应环境的变化;⏹个体大小与物种的分类数量(多样性)、个体数量有联系;⏹生物对食物的需求随个体增大而增加,但是单位需求量却下降。
个体大小与其在生态系统中的作用在生态学研究中经久不衰。
普通生态学 生长:包括生物体细胞数量和生物物质的增加。
发育:伴随着生长过程,生物体的结构和功能从简单到复杂,从幼体形成一个与亲代相似的性成熟的个体。
1.2生长与发育速度普通生态学有关生长发育的几个概念 变态 (metamorphosis): The presence of a major developmental change in shape or form from thejuvenile to the adult.滞育 (diapause): The presence of a resting stage in the life history.衰老 (senescence): The process and timing of aging, degeneration, and death.普通生态学 繁殖:是指有机体生产出与自己相似后代的现象。
生活史对策概述大气基地贺园园1111700026生活史与生活史对策:生活史(life history)意为生物从其出生到死亡所经历的全部过程,也叫生活周期(life cycle)。
生活史性状包括出生时个体大小;生长形式;成熟年龄;成熟时个体大小;后代的数量、大小、性比;特定年龄和大小的繁殖投入;特定年龄和大小的死亡规律;寿命等。
生活史对策(life history strategy)是指生物在生存斗争中获得的生存对策,也称生态对策(bionomic strategy )或进化对策,例如生殖对策、取食对策、迁移对策、体型大小对策等主要内容:任何生物做出的任何一种生活史对策,都意味着能量的合理分配,并通过这种能量使用的协调,来促进自身的有效生存和繁殖。
每个生物具有生长、维持生存和繁殖三大基本功能,生物必须采取一定的策略配置能够获得的有限资源,其核心主要强调在特定环境中提高生殖、生存和生长能力的组合方式.1)生长对策●生长速度早期演替种:早期迅速生长,具开拓对策(白桦)后期演替种:早期生长缓慢,具保守对策(红松)●生长方式以温带木本植物为例,其顶枝形成有两种主要方式:①有限生长类型:顶枝在冬季完全定型,冬芽形成时就决定了叶子数目。
②无限生长类型:冬芽只含有少量叶原基,在下一个生长季,顶枝尖端在生长季内还能产生新的叶子和节间。
●根冠比率—物质分配2)生殖对策生殖对策实际就包括两个方面的问题:第一是生殖者存活的问题,也即生殖的代价问题,生物生殖必然带来变化的生理压力和个体危险,因此,也就必然会影响到生物的生存;第二是生殖的效率问题,生物选择的对策,都旨在提高生殖的效率,这一点可从植物的生殖行为中证实。
1)体型效应物种个体的大小与其寿命有很强的正相关关系。
2)成体的存活与繁殖成熟个体存活率低,生物繁殖越早,投资于繁殖的能量越多;成熟个体存活率越高,生物的繁殖期越晚(个体较大),分配于繁殖的能量就越少。
3)当前繁殖与未来繁殖如果未来生命期望低,分配给当前繁殖的能量应该高,而如果剩下的预期寿命很长,分配给当前繁殖的能量应该较低。
机会生活史的生物生殖力机会生活史的生物生殖力是指生物在特定环境下,通过生殖器官的发育与功能,实施生殖策略,繁衍后代的能力。
生物生殖力在生物进化、种群繁衍和生态系统稳定中起着至关重要的作用。
本文将从生物生殖力的关键因素、影响因素以及提高与改良等方面进行探讨,以期为生物学、生态学和环境科学等领域的研究提供理论参考。
生物生殖力的关键因素包括生殖器官的发育与功能、生殖策略与生殖行为以及环境适应与生态位。
首先,生殖器官的发育与功能是生物生殖力的基础。
各种生物在漫长的进化过程中,形成了各自独特的生殖器官和生殖方式。
其次,生殖策略与生殖行为是生物在生殖过程中采取的策略和行为模式,如一次性生殖、多次性生殖、内部受精、外部受精等,这些策略和行为对生物的生殖力具有重要影响。
最后,环境适应与生态位是生物在自然界中生存和繁衍的基础,生物必须适应不断变化的环境,才能保证种群的繁衍。
生物生殖力受到多种因素的影响,包括遗传与变异、生殖周期与生殖节律以及生殖障碍与生殖疾病。
遗传与变异是生物进化的基础,通过对遗传物质的传递和变异,生物能够在环境中不断适应和进化。
生殖周期与生殖节律是生物生殖过程中的重要调控因素,如季节性生殖、日周期性生殖等,这些周期和节律影响着生物的生殖力和繁殖成功率。
此外,生殖障碍与生殖疾病也会对生物生殖力产生影响,如生殖器官的损伤、生殖激素水平的失衡、生殖道感染等,这些都可能导致生物生殖力的下降。
为了提高生物生殖力,科研人员和兽医工作者采取了多种措施。
首先,应用生殖生物技术,如人工授精、胚胎移植、基因编辑等,以提高繁殖效率和后代品质。
其次,改善生态环境,保护生物栖息地,降低生殖障碍和疾病的发生。
最后,加强生殖健康教育,提高人们对生殖健康的认识和重视,从而为生物的生殖力提高创造有利条件。
总之,生物生殖力在机会生活史中具有重要作用。
了解生物生殖力的关键因素、影响因素以及提高与改良措施,有助于我们更好地保护生物多样性,维持生态系统稳定,并为未来生物科技发展提供方向。
第2章生活史进化张大勇生活史进化对策的研究起始于本世纪40年代末~50年代初,主要是由动物种群统计学(demography)和进化理论相结合而形成的。
在1920~1950年这一时期,生态学家已经开始广泛地运用寿命表方法研究动物种群,因而对于生活史的定量种群统计学后果已经具备了一个有效的分析方法。
这种方法考察的是特定年龄个体的死亡率和生育率。
生态学家已清楚地知道,这些生活史参数无论是在种内还是在种间都有很大的变异。
种群遗传学和数量遗传学的迅猛发展同时也为达尔文关于表型性状适应价值的论述提供了坚实基础。
在第1章内,我们已经提到,早期的种群生态学并不关注种群内部的遗传变异,而种群遗传学也基本上忽略了种群动态过程。
二者之间的有机结合是生态学领域内长期没有得到很好解决的一个难题;而这对于生活史对策研究却是至关重要的。
尽管Fisher(1930)早在30年代就已经提出应把种群统计学性状看作为表型的一部分并探索它们的适应性基础,但人们公认现代生活史进化理论创立于40年代末到50年代初Lack(1947)关于鸟类窝卵数、Medawar(1946,1952)关于衰老、以及Cole(1954)关于单次生殖/多次生殖进化的研究。
其后,生活史进化方面的研究蓬勃兴起,有关文献可说是浩如烟海。
但在本章内我们并不打算对整个领域进行全面地综述,而是选择几个有代表性的核心问题介绍其理论背景和发展趋势。
如果读者想要更全面地了解该领域,可以参阅Roff(1992)以及Stearns (1992)的专著。
侧重于基础理论方面的书籍有Charlesworth(1994)。
在进入本章具体内容之前,我们有必要首先熟悉一下生活史进化研究的基本途径—表型优化理论(参见第3章)。
2.1 进化生物学中的表型优化研究途径近些年,进化生物学家和生态学家已经开始广泛采用工程学和经济学领域内的数学方法来认识生命的多样性问题(Maynard Smith 1978)。
具体涉及到的数学分支包括有控制论、数学规划、对策论等。
在进化生物学中,最优化处理方法(optimization approach)的一个基本前提是:适应性是生命有机体普遍存在的一个性质,并且可以由自然选择过程加以解释。
或者说,适应性作为优化理论的基本出发点,不再考虑如果生物进化不产生适应性的可能。
优化理论只是帮助人们理解适应性产生的选择力量。
这种研究途径假定存在着这样一个优化指标:最优的生活史(或者行为、结构等)应该使得这个指标达到最大值(e.g. Parker & Maynard Smith 1990; Lessells 1991; Roff 1992; Stearns 1992; Charlesworth 1994; Daan & Tinbergen 1997)。
使用这些手段可以帮助人们整理和系统阐述生物行为、结构以及生活史对策的功能解释,并对这些解释进行经验验证(empirical testing)。
2.1.1 表型优化模型所有优化模型都包括一些关于所研究对象的约束条件、优化指标以及遗传方式的假设。
约束条件,又称为对策集(strategy set),是指所有可能的、并受自然选择作用的表型。
显然,什么是“最优的”取决于约束条件是什么,即有哪些可能的表型。
自然界不存在绝对意义上的“最佳表型”。
任何一个有机体(既使是自养生物)都不可能在同一时间内做好每一件事情,因为可用于生长、维持和生殖的能量(或者时间或者其它资源)必然是有限的。
生物必须在这些相互冲突的需求中进行有效的资源分配以寻找一个最佳的分配方案。
例如,利用分生组织和有限碳源,一株植物既可形成花也可以形成叶片,但不能二者同时形成。
对这些分配问题的不同解决方案就形成了各式各样的生活史。
有时,约束条件本身也是自然选择作用的结果;但在分析目前所面临的问题时,人们可以简单地把它们看成不再发生改变的进化背景。
从这个意义上说,所有进化优化模型分析的都是局部(local)适应性问题(Stearns 1992)。
例如,人们分析有性生物的生殖分配时经常假定性别资源分配(sex allocation)是恒定的,不再同时考虑性别资源分配的进化(参见Zhang & Wang 1994)。
生活史理论中关于约束条件的一个重要思想就是“不可兼顾”(trade-off)的概念,或称为“负耦联”。
它通常泛指任何形式上的生活史性状之间的负相关关系。
为了使讨论清晰,Stearns(1992)建议区分三种意义上的负耦联关系,即生理负耦联、微观进化(microevolutionary)负耦联和宏观进化(macroevolutionary)负耦联。
生理负耦联是指直接竞争有机体内有限资源的两个(或两个以上)过程之间的分配权衡。
例如,增加进入种子的能量和物质必然使得植物茎杆、叶片以及根系等营养器官所获得的资源分配量减少,使个体生长、存活都受到影响。
微观进化负耦联是指一个(增加适合度)性状的改变必然与另一个(减少适合度)性状的变化相伴随。
如果微观进化负耦联关系不存在,那么选择将驱使所有与适合度相关的性状都达到由历史和设计制约的极限。
事实上,许多生活史性状远未达到这些极限值,说明微观进化负耦联关系在许多生物身上必定存在。
几乎所有微观进化负耦联的背后都隐藏着生理负耦联,但生理负耦联却有可能不是微观进化负耦联。
例如,考虑这样一种生物,它在产生后代大小与数量方面具有反应规范(或称表型可塑性)。
当食物短缺时,它生产的后代数量少、个头大;而当食物丰富时,它生产的后代数量多、个头小。
在这个例子中,后代个体大小与数量之间的生理负耦联关系是很明确的,但种群进化反应不会出现,因为反应规范没有遗传上的变异。
如果说一个负耦联是微观进化意义上的,人们必须能证明它对选择有反应。
微观进化负耦联是种群水平上对生理负耦联遗传变异选择压力的进化反应。
不管生理负耦联遗传变异今天是否仍然存在,过去它必然曾经存在过,并可能因它已是最优的分配规则而被固定下来。
宏观进化负耦联是指性状之间在独立的系统发育中的相关变化。
考虑一个极端情况。
让我们设想没有遗传变异也没有可塑性的两个性状,它们在物种内已被固定并且也不随环境发生改变。
尽管如此,当人们考察属内不同物种或者科内不同属时,这两个性状却可能表现出负相关关系。
这时我们假定这种模式能够出现的原因只能是由于过去存在的生理和微观进化负耦联在整个谱系内留下了痕迹,虽然我们现在无法在种内观察到它们。
当然,绝大多数宏观进化负耦联都伴随有生理和微观进化负耦联的存在,但它们不见得有相同的强度或正负关系。
进化优化模型通常只考虑生理和微观进化负耦联,宏观进化负耦联关系并不直接反映在优化的约束条件之内。
通过比较宏观进化模式,我们可以鉴别出整个谱系共同具备的条件,得出更一般性的结论。
宏观进化负耦联还可以为我们指明哪些生理和微观进化负耦联需要进一步深入研究,例如相对于谱系平均值的种内负耦联的偏离等。
在给出约束条件后,或者说已建立起性状之间的负耦联关系,下一步就是确定优化指标。
通常来说,优化的指标都是达尔文适合度的某种计测,有时还是间接的度量。
例如,Maynard Smith & Savage (1956)分析动物奔跑速度时以最小能量消耗(或能量消耗保持不变时奔跑速度最大)为优化指标;而在动物觅食理论中则经常假定自然选择会使有机体净能量摄取速率达到最大。
这时模型隐含的假设是适合度与这些间接指标之间正相关。
在生活史进化研究领域,人们最常用的适合度指标有以下两个:不考虑密度制约效应,人们一般采用种群增长速率(或称为马尔萨斯参数)来作为生活史进化的优化指标(如果种群增长是连续的,种群瞬时增长速率用符号r 来表示;如果种群增长是离散的,则用符号λ来表示。
λ和r 的关系为:r e -=λ);如果种群个体数量由于密度制约的作用而保持着稳定,那么适合度也可由有机体一生中期望的后代数(或者称为净生殖速率)来测度,一般用数学符号R 0表示。
具体地,若令l x 表示个体(雌雄异体生物则为雌性个体)从出生到x 年龄的存活概率,m x 为x 龄个体的生育力(雌雄异体生物用生育的女儿数来计算),那么λ(或r )和R 0可分别由下式求出:∑∑∞=∞=--==111x x x x rx x x x m l e m l λ(2.1)∑∞==10x x x m lR (2.2)对于连续增长的种群,以上两式亦可分别表示为:⎰∞-=01dx m l e x x rx(2.1´) ⎰∞=00dx m l R x x(2.2´) 公式(2.1´)通常称为Euler-Lotka 方程。
不考虑密度依赖的作用时,人们一般常用r (或者λ)来做适合度指标;而对于个体数量恒定(由于密度依赖效应)的种群,优化指标既可以采用r 也可以采用R 0(Stearns 1992;Kozlowski 1993;Charlesworth 1994),但这时因为密度恒定显然必须有r =0及R 0=1。
这似乎是一个矛盾:这些适合度指标既要被自然选择最大化又只能为恒定值。
用进化稳定对策(ESS )语言可以较好地解决这个难题(参见第3章)。
在平衡状态下,不能被任何其它基因型侵入的基因型将会在进化竞赛中获胜,或者说最佳基因型的r =0以及R 0=1,而其它入侵基因型的r <0,或R 0<1。
尽管优化指标的选取应看作为模型假设条件的一部分,但选取什么指标并非是任意的,需要做出理论上的合乎逻辑的说明。
选择了某个优化指标往往就附带选择了为使该优化指标成立的限定条件。
对于无性生物如果再忽略竞争引起的频率依赖问题(Kawecki 1993),上面所规定的适合度指标可能是广泛适用的。
对于有性生物来说,即使忽略了频率依赖问题,r 或者R 0可以作为适合度指标仍然要求雌性适合度和雄性适合度都是资源投入的线性函数(Zhang & Wang 1994; Zhang et al. 1996; Zhang & Jiang 1997;参见第4章)。
应用ESS概念框架处理频率依赖条件下的适合度问题是今后生活史理论工作发展的一个方向。
在表型优化理论中,假定的遗传方式对选择优化指标有很大影响。
为了使问题简化,人们经常明确或暗含地假设无性(孤雌)生殖,或者真实遗传(breed true)。
通俗地说就是假定“龙生龙,凤生凤”;后代与亲代之间由于有性生殖过程而必然存在的遗传差异被忽略了。
因为我们面对的有机体绝大多数都是有性生殖的二倍体生物,所以这个假设只能理解为对非常复杂问题的一个初步探索。
对于有性生殖的生物,这个假设可以成立的范围有多大?它要求哪些前提条件以及适用于哪些具体问题?这些疑问总体上说仍然悬而未决。