第三章遗传的染色体学说
- 格式:doc
- 大小:739.54 KB
- 文档页数:25

目 录第一部分 考研真题精选一、选择题二、填空题三、判断题四、名词解释五、问答题第二部分 章节题库第一章 绪论第二章 孟德尔定律第三章 遗传的染色体学说第四章 孟德尔遗传的拓展第五章 遗传的分子基础第六章 性别决定与伴性遗传第七章 连锁交换与连锁分析第八章 细菌和噬菌体的重组和连锁第九章 数量性状遗传第十、十一章 遗传物质的改变第十二章 重组与修复第十三章 细胞质和遗传第十四章 基因组第十五章 基因表达与基因表达调控第十六章 遗传与个体发育第十七章 遗传和进化第一部分 考研真题精选一、选择题1以下哪种性染色体-常染色体套数,会出现雄性果蝇( )。
[中山大学2019研]A.XX:AAB.XXY:AAC.XXXA:AAAD.X:AA【答案】D【解析】果蝇的性别由X染色体数目与常染色体组数之比决定,与Y无关。
X:A的比值≥1时发育为雌性,≤0.5发育为雄性。
ABC三项错误,X:A的比值等于1,出现雌性果蝇。
D项,X:A的比值小于1,出现雄性果蝇。
2基因型为aaBbCcDd个体自交后代中,出现aaBbccDd的概率是( )。
[湖南农业大学2018研]A.1/4B.1/8C.1/16D.1/32【答案】CaaBbCcDd个体自交,将各基因分开考虑,后代aa的概率为1,【解析】Bb的概率为1/2,cc的概率为1/4,Dd的概率为1/2,因此出现aaBbccDd 的概率为1×1/2×1/4×1/2=1/16。
3对于拟南芥短径突变,己分离到纯合的品系并获得短径与长径的个体数目分别为62与38,则该突变的外显率为( )。
[中山大学2019研]A.0.62B.0.38C.0.613D.0.387【答案】A外显率=62/(62+38)=0.62。
【解析】4细胞减数分裂终变期能产生四体环的是( )。
[沈阳农业大学2011研]A.易位纯合体B.易位杂合体C.四分体D.四合体【答案】B易位杂合体是两条非同源染色体间互换片段,另外两条不发生【解析】互换,从而形成十字形结构的四体环。




《遗传的染色体学说》讲义在生命的奥秘中,遗传现象一直是人们探索和研究的重要领域。
而遗传的染色体学说的提出,为我们理解遗传的本质和规律提供了关键的理论基础。
首先,让我们来了解一下什么是染色体。
染色体是存在于细胞核中的线状结构,由 DNA、蛋白质和少量 RNA 组成。
在细胞分裂时,染色体可以清晰地被观察到,它们呈现出特定的形态和特征。
那么,遗传的染色体学说究竟是什么呢?简单来说,它认为基因位于染色体上,染色体是基因的载体。
这一学说的提出并非一蹴而就,而是经历了众多科学家的研究和探索。
早在19 世纪末,孟德尔通过豌豆杂交实验发现了遗传的基本规律,但当时人们并不清楚基因在细胞中的位置和作用方式。
随着显微镜技术的发展,人们能够更清晰地观察到细胞的结构和染色体的行为。
在这个过程中,萨顿通过对蝗虫生殖细胞的研究,观察到染色体在减数分裂过程中的行为与孟德尔遗传定律中基因的分离和组合规律十分相似。
他由此提出了染色体学说的初步想法,即染色体在遗传中可能起着重要的作用。
摩尔根则通过果蝇杂交实验,为遗传的染色体学说提供了有力的证据。
他发现果蝇的某些性状与特定的染色体相关联,进一步证明了基因位于染色体上。
那么,染色体是如何实现遗传功能的呢?在细胞分裂过程中,染色体进行复制,然后在减数分裂时,同源染色体配对并发生交换,这使得遗传物质能够重新组合,增加了遗传的多样性。
基因在染色体上呈线性排列,不同的基因位于不同的位置。
当染色体传递给子代细胞或个体时,其上的基因也随之传递,从而实现了遗传信息的传递。
遗传的染色体学说对于我们理解生物的遗传现象具有重要意义。
它解释了为什么子代能够继承亲代的特征,以及遗传变异是如何产生的。
例如,在有性生殖过程中,双亲的染色体通过配子的结合传递给子代,使得子代获得了双亲的遗传信息。
同时,由于减数分裂过程中的染色体交换和随机组合,子代又具有了不同于双亲的新的遗传组合,这就是遗传变异的来源之一。
此外,遗传的染色体学说也为遗传学的进一步发展奠定了基础。




第三章遗传的染色体理论及性连锁遗传第一节染色体的形态是细胞核最重要的组成部分。
着丝点:在光学显微镜下,每个染色体都有一个着丝点,细胞分裂过程中,着丝点对染色体向两极移动具有决定性的作用。
若染色体由于某种原因发生断裂,势必成为两段,一段有着丝点:有着丝点的片段可以正常地移向两极,另一端没有着丝点:无着丝点的片段则不能正常地移向两极,常常会丢失在细胞质中着丝点将染色体分为两个臂(arm)。
用碱性染料对染色体进行染色,当两个臂被染色时,着丝点不着色,在光镜下,好象染色体在此区域是中断的。
于是又称着丝点区域为主缢痕(primary constriction)。
次缢痕和随体有些染色体上除了主缢痕区之外,还有一个不着色或着色很淡的区域,通常位于短臂上,称为次缢痕(secondary constriction)。
次缢痕的外侧还有一部分染色体,这一部分可大可小,有时其直径与该染色体相同,有时较小,称为随体(satellite)。
次缢痕的位置和随体的大小,在一个物种内是相对固定的,这些也是识别特定染色体的重要的形态标志。
有的物种则有两对或两对以上的染色体带有次缢痕和随体。
次缢痕与核仁的形成有关。
带有随体的染色体称为随体染色体(satellite chromosome)。
着丝点的位置决定染色体的形态,将染色体分为两个臂。
臂比(arm ratio):染色体的长臂与短臂的长度之比,长臂/短臂表染色体形状与臂比染色体名称臂比中期后期中部着丝粒染色体(metacentric chr.)≈1X字形V字形近中着丝粒染色体(submetacentric chr.)1~1.7似X L字形近端着丝粒染色体(acrocentric chr.) 1.7~3.0似X 棒状端部着丝粒染色体(telocentric chr.) >3.0 倒V 棒状或点第二节有丝分裂和减数分裂有丝分裂中期后期有丝分裂(mitosis)的遗传学意义:从遗传学的观点看,有丝分裂的重要意义就在于其分裂产物都是相同的,它们精确地含有和亲代细胞核完全相同的遗传物质。

第三章遗传的染色体学说1 有丝分裂和减数分裂的区别在哪里?从遗传学的角度来看,这两种分裂各有什么意义?那么,无性生殖会发生分离吗?试加以说明。
解:有丝分裂和减数分裂的区别:(1)有丝分裂是体细胞的分裂方式,而减数分裂一般仅存在于生殖细胞中。
(2)有丝分裂DNA复制一次,细胞分裂一次,染色体数由2n-2n,减数分裂DNA 复制一次,细胞分裂两次,染色体数由2n-n。
(3)有丝分裂在S期进行DNA合成,然后经过G2期进入有丝分裂期。
减数分裂前DNA合成时间较长,合成后立即进入减数分裂,G2期很短或没有。
(4)有丝分裂时每一条染色体独立活动,减数分裂中染色体会发生配对、联会、交叉、交换等。
(5)有丝分裂进行的时间较短,一般为1-2小时,减数分裂进行时间长, 例如人的雄性配子减数分裂需24小时,雌配子甚至可长达数年。
有丝分裂的遗传学意义:通过有丝分裂维持了生物个体的正常生长和发育(组织及细胞间遗传组成的一致性);并且保证了物种的连续性和稳定性(单细胞生物及无性繁殖生物个体间及世代间的遗传组成的一致性)。
减数分裂的遗传学意义:(1)通过减数分裂和受精过程中的染色体数目交替(2n-n-2n),保证了物种世代间染色体数目的稳定性。
(2)在减数分裂过程中,由于同源染色体分开,移向两极是随机的(染色体重组) ,加上同源染色体的交换(染色体片断重组) ,大大增加了配子的种类,从而增加了生物的变异,提高了生物的适应性,为生物的发展进化提供了物质基础。
无性生殖不经过两性生殖细胞的结合,而是由生物体自身的分裂生殖或其体细胞生长发育形成个体过程一般没有和其他个体或结构发生基因交流,自身也不发生减数分裂,因此在正常情况下不会发生分离,但由于外界环境条件的影响通过无性生殖方式产生的个体也有可能会发生变异。
2 水稻正常的孢子体组织,染色体数目是12对,问下列各组织的染色体数目是多少?(1)胚乳;(2)花粉管的管核;(3)胚囊;(4)叶;(5)根端;(6)种子的胚;(7)颖片;解析:(1)胚乳3n=36(2)花粉管的管核n=12(3)胚囊8n=96(4)叶2n=24(5)根端2n=24(6)种子的胚2n=24(7)颖片2n=243 用基因型Aabb的玉米花粉给基因型AaBb的玉米雌花授粉,你预期下一代胚乳的基因型是什么类型,比例如何?解析:基因型Aabb的花粉产生的雄配子Ab,ab基因型AaBb产生的极核为AB,Ab,aB和ab胚乳基因型为AAABBb,AAAbbb,AaaBBb,Aaabbb,AAaBBb,AAabbb,aaaBBb和aaabbb,比例相等。
第二章孟德尔定律1、为什么分离现象比显、隐性现象有更重要的意义答:因为1、分离规律是生物界普遍存在的一种遗传现象,而显性现象的表现是相对的、有条件的;2、只有遗传因子的分离和重组,才能表现出性状的显隐性。
可以说无分离现象的存在,也就无显性现象的发生。
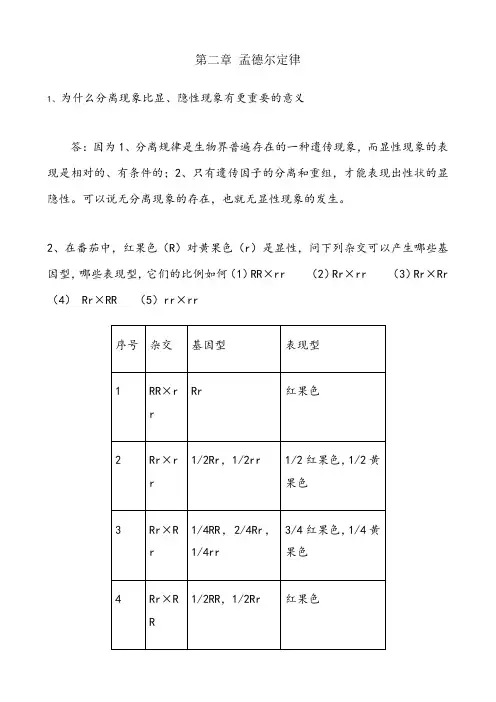
2、在番茄中,红果色(R)对黄果色(r)是显性,问下列杂交可以产生哪些基因型,哪些表现型,它们的比例如何(1)RR×rr(2)Rr×rr(3)Rr×Rr (4)Rr×RR(5)rr×rr3、下面是紫茉莉的几组杂交,基因型和表型已写明。
问它们产生哪些配子杂种后代的基因型和表型怎样(1)Rr ×RR (2)rr ×Rr (3)Rr ×Rr 粉红红色白色粉红粉红粉红4、在南瓜中,果实的白色(W)对黄色(w)是显性,果实盘状(D)对球状(d)是显性,这两对基因是自由组合的。
问下列杂交可以产生哪些基因型,哪些表型,它们的比例如何(1)WWDD×wwdd(2)XwDd×wwdd(3)Wwdd×wwDd (4)Wwdd×WwDd5.在豌豆中,蔓茎(T)对矮茎(t)是显性,绿豆荚(G)对黄豆荚(g)是显性,圆种子(R)对皱种子(r)是显性。
现在有下列两种杂交组合,问它们后代的表型如何(1)TTGgRr×ttGgrr (2)TtGgrr×ttGgrr解:杂交组合TTGgRr × ttGgrr:即蔓茎绿豆荚圆种子3/8,蔓茎绿豆荚皱种子3/8,蔓茎黄豆荚圆种子1/8,蔓茎黄豆荚皱种子1/8。
杂交组合TtGgrr × ttGgrr:即蔓茎绿豆荚皱种子3/8,蔓茎黄豆荚皱种子1/8,矮茎绿豆荚皱种子3/8,矮茎黄豆荚皱种子1/8。
6.在番茄中,缺刻叶和马铃薯叶是一对相对性状,显性基因C控制缺刻叶,基因型cc是马铃薯叶。
第三章遗传的染色体学说名词:染色体真核细胞细胞分裂时期可以被碱性染料着色的一类小体。
是遗传物质的载体。
染色体组二倍体生物的配子中所包含的形态、结构和功能上彼此不同的一组染色体称为染色体组。
同源染色体是指二倍体细胞中来源相同,形态、结构和功能相似,分别来自父母一方的一对染色体。
同源染色体上分别携带着控制相对性状的等位基因。
染色体组型把某生物个体或分类群体的体细胞内有恒定特征的整套染色体按它们的相对恒定特征排列起来的图像。
减数分裂是有性生殖配子形成过程中进行的一种有丝分裂。
染色体在前减数分裂间期复制一次,而细胞连续分裂两次,第一次分裂着丝粒不分开,只是同源染色体分离,导致子细胞中染色体数目减半,因此称为减数分裂。
染色体组型分析将一个细胞的有丝分裂中期染色体用显微照片剪贴的方法,按一定系统排列起来,对其染色体数目、长度、着丝粒及副缢痕的数目、大小、位置等染色体形态特征进行分析研究。
姊妹染色单体由一条染色体经复制形成的仍由着丝粒相连的两个子染色体。
习题一、问答题原核细胞与真核细胞的遗传基础的异同点是什么?原核细胞真核细胞细胞核无膜包围,为类核核有双层膜包围染色体环状DNA分子核中有染色质或染色体。
线粒体和叶绿体中为环状DNA分子连锁群数目一个基因连锁群1个或多个连锁群染色体组成DNA裸露或结合少量蛋白质DNA同组蛋白结合,线粒体和叶绿体中的DNA裸露DNA顺序很少或没有重复顺序有重复顺序细胞分裂无丝分裂有丝分裂性系统基因由供体质受体单向传递减数分裂后形成的含同等基因组的配子融合绘有丝分裂中期典型染色体的形态结构图,并注明各部分各称。
3.简述核小体的结构和染色体的四级结构模型。
染色体的一级结构:核小体染色体二级结构:螺旋体染色体三级结构:超螺旋体染色体四级结构:中期染色体。
4.简述原核生物的细胞分裂过程。
5.什么是常染色质和异染色质?常染色质是细胞分裂间期和早前期染色质丝折叠疏松,高度伸展,染色时不易分辩,而在细胞分裂中期螺旋折叠达到最大程度,染色时着色很浓的染色质。
第三章遗传的染色体学说孟德尔的分析之美,是因为从杂交试验得到的资料,可以用分离定律和自由组合定律来说明;而且还可根据这两个定律,预测另一些杂交实验的结果。
孟德尔的分析都用假设的遗传因子或基因来表示,但没有谈到基因的物质基础或基因在细胞中的位置。
尽管孟德尔定律在很多生物中得到证实,但下一个问题显然是:细胞中什么结构可以和基因相对应?早在1900年以前,生物学家就仔细地记录了细胞分裂时显微镜下可以观察到的一系列变化。
他们注意到细胞内的细胞核(nucleus)和细胞质(cytoplasm)的显明区别,也在分裂时看到叫做染色体(chromosomes)的细胞器的出现,并且详细地描述了细胞分裂和受精过程中染色体的行为。
孟德尔定律在1900年重新发现后,Sutton和Boveri就注意到杂交试验中基因的行为跟配子形成和受精过程中染色体的行为完全平行,因而他们在1903年提出假设,认为孟德尔遗传因子是在染色体上。
我们要懂得这个假设,想先说明细胞的一般结构,进而详细地说明细胞分裂中的染色体行为。
第一节细胞生命的单位是细胞。
细胞主要可分细胞质和细胞核,细胞核位于细胞质之中(图3-1)。
动植物细胞质周围有细胞膜(cell mem-brane),植物细胞的细胞膜外面还有细胞壁(cell wall)。
细胞质细胞质是粘稠的胶体,周围有细胞膜。
细胞膜由内外两分子层构成,其成分是脂质,两层之间嵌有蛋白质。
细胞膜把细胞分为内外两侧,它的功能是维持细胞内物质浓度和电化学梯度,调节代谢物质的出入等。
细胞质中含有:中心粒(centrioles)主要见于动物,由27条很短的微管(microtubules)组成;从横切面看,是9个三联微管(9×3=27)盘绕成的环状结构。
可能与细胞分裂有关,确切作用还不清楚。
线粒体(mitochondria)是双膜构造,外层是外壁,里面是内壁。
内壁向内突出形成很多褶叠,内含有产生能量的酶系,包括三磷酸腺苷(ATP)的合成和还原,碳素化合物的氧化。
它的位置往往是在进行吸能过程(endergonic process)的结构附近,如靠近收缩单位。
核糖体(ribosomes)核糖体常存在于由膜组成的网状结构——内质网(endoplasmic reticulum)上。
内质网主要分颗粒型和非颗粒型两种,核糖体存在于颗粒型内质网上。
它是由大小两个亚单位构成的小颗粒。
蛋白质的合成在核糖体上进行。
溶酶体(lysosomes)一种微小颗粒,呈球形或长圆形,也有双层的膜。
内含蛋白酶、核酸酶和其它水解酶(hydrolase),但大都是一些原酶(proenzymes),处于失活状态。
在高等生物中,当细胞停止执行功能时,这些酶参与细胞的加水降解(hydrolytic de-gradation)。
高尔基体(golgi bodies)见于某些细胞,可能由内质网发展而成。
看来有分泌,贮藏和排泄各种物质的作用。
质体(plastids)仅见于植物,其中最重要的是叶绿体。
叶绿体的表面是双层的膜,内部为无色的基质,基质内有绿色基粒(grana),基粒中含有与光合作用有关的酶和色素,光合作用即在基粒中进行。
已经知道,中心粒、线粒体和质体内含有脱氧核糖核酸(DNA),核糖体内含有丰富的核糖核酸(RNA)。
细菌细胞中有核糖体,但没有线粒体、高尔基体和内质网等。
细胞核在细菌和蓝绿藻(blue-green algae)中,核物质并不和细胞质分开,看不到核的结构,所以它们属于原核类(prokar-yotes)。
大多数现存的种和多细胞生物中,有一核膜(nuclearmembrane)把核质和细胞质分开,可以看到核的结构,所以它们属于真核类(eukaryotes)。
真核类的核膜也是双层的膜,而且跟内质网和细胞膜紧密地联系着。
细胞核中大多数地方充满了核液(nuclear sap)其中有一种很容易被碱性染料染上颜色的物质,称为染色质(chromatin),这是细胞核的主要成分之一。
在细胞分裂时,染色质呈现为形状清晰的染色体。
核中还有一个到几个核仁(nucleolus,复数nucleoli)。
核仁中富有DNA和RNA,是核糖体RNA合成的地方,也是核糖体装配的地方。
染色体因为基因是在染色体上,所以我们把染色体另立一项,详细地谈一谈。
每一种生物的染色体数是恒定的。
多数高等动植物是二倍体(diploid),也就是说,每一身体细胞中有两组同样的染色体(有时与性别直接有关的染色体,即性染色体,可以不成对)。
亲本的每一配子带有一组染色体,叫做单倍体(haploid),用n来表示。
两个配子结合后,具有两组染色体,叫做二倍体,用2n表示。
例如玉米的二倍体染色体数是20(2n=20),即有10对染色体(图3-2)。
人的染色体数是46(2n=46),即有23对染色体(图3-3)。
但多数微生物的营养体是单倍体,例如链孢霉的单倍体染色体数是7。
现在把各种生物的染色体数择要列出(表3-1)。
表3-1各种生物的染色体数通名学名二倍体数动物人类Homo sapiens46金丝猴Rhinopithecus rhinopithecusroxellanae44弥猴Macacamalatta42黄牛Bostaurus60猪Susscrofa40狗Cantis familiaris78猫Felis domesticus8马Equus Calibus4驴Equus asinus62山羊Capara hircus60绵羊Ovis aries54小家鼠Mus musculus 40大家鼠Rattus norvegicus 42水貂Mustela vison 30豚鼠Cavia cobaya 64兔Oryctolagus cuniculus 44家鸽Columba livia domesticus约80鸡Gallus domesticus约78火鸡Meleagris gallopavo约80鸭Anas platyrhyncho约80家蚕Bombyx mori56家蝇Musca domestica12果蝇Drosophila melanogaster8蜜蜂Apis mellifera♀3216蚊Culex pipiens6佛蝗Phlaeobainfumata24♀23淡水水螅Hydra vulgaris attenuata32植物洋葱Alliumcepa16大麦Hordeum uulgare 14水稻Oryza sativa 24小麦Triticum vulgare42玉米Zea mays20金鱼草Antirrhinum majus 16陆地棉Gossypium hirsutum 52中棉Gossypium arboreum26豌豆Pisum sativum 14香豌豆Lathyrus odoratus14蚕豆Vicia faba12菜豆phaseolus vulgaris22向日葵Helianthusannuus34烟草Nicotiana taldcum 48番茄Solanum lycopersicum24松Pinus species 24青菜Brassica chinensis 20甘蓝Brassica oleracea 18月见草Oenothera biennis 14单倍体数链孢霉Neurospora crassa 7青霉菌Penicillium species 4曲霉Aspergillus nidulans8衣藻Chlamydomonas reinhardi 16面包酵母Saccharomyces cerevisiae17染色体在复制以后,含有纵向并列的两个染色单体(chroma-tids),只有在着丝粒(centromere)区域仍联在一起。
着丝粒在染色体上的位置是固定的。
由于着丝粒位置的不同,把染色体分成大致相等或长短不等的两臂(arms)。
着丝粒的位置在染色体中间或中间附近时,染色体两臂的长度差不多,这着丝粒叫做中间着丝粒或亚中间着丝粒。
着丝粒的位置靠近染色体的一端时,根据着丝粒离开端部的远近,这着丝粒叫做近端部着丝粒或端部着丝粒。
着丝粒所在的地方往往表现为一个缢痕,所以着丝粒又称初级缢痕(primary constriction)。
有些染色体上除了初级缢痕以外,还有一个次级缢痕(secon-dary constriction),连上一个叫做随体(satellite)的远端染色体小段。
次级缢痕的位置也是固定的。
在细胞分裂将结束时,核内出现一个到几个核仁,核仁总是出现在次级缢痕的地方,所以次级缢痕也叫做核仁形成区(nucleolar organizer)(图3-4)。
染色体主要由DNA和蛋白质这两类化学物质所组成。
每一染色单体的骨架是一个连续的DNA大分子,许多蛋白质分子结合在这DNA骨架上,成为DNA蛋白质纤丝。
一般认为细胞分裂中期(见本章第二节)时看到的染色单体就是由一条DNA蛋白质纤丝重复折叠而成的(图3-5)。
染色体上的蛋白质有两类:一类是低分子量的碱性蛋白即组蛋白(histones),另一类是酸性蛋白质,即非组蛋白蛋白质(non-histone proteins)。
非组蛋白蛋白质的种类和含量不十分恒定,而组蛋白的种类和含量都很恒定,其含量大致与DNA相等。
所以人们早就猜测,组蛋白在DNA·蛋白质纤丝的形成上起着重要作用。
Kornberg根据生化资料,特别是根据电镜照相,最先在1974年提出绳珠模型(beads on-a-string model),用来说明DNA·蛋白质纤丝的结构(图3-6)。
他认为纤丝的结构单位是核体(nuc-leosome)。
每个核体的核心由8个组蛋白分子,即HA,H3B,H3和H4各两个分子组成,呈调扁圆形。
2间有DNA连丝珠,也就是DNA·蛋白质纤丝(linker),连丝约有50—60个核苷酸对,连丝上另有一个组蛋白分子H1。
核心外周的DNA和相应的连丝DNA约共有200个核苷酸对。
核体与核体以一定间隔相连,成为一串珠子,所以这模型称为绳珠模型。
但以后知道,绳珠模型是制备染丢失的结果。
在活细胞中核体与核体是相互色质样品时,DNA连丝上H1紧靠的,由于连丝上不同H的相互作用,核体键又可卷曲盘旋,呈现螺1线管状(solenoidal structure)(图3-6)结构,这可能是间期核染色质的原有结构。
关于这儿提到的DNA和蛋白质的化学性质和功能,留待第十一章说明。
第二节细胞分裂细胞是靠分裂而增殖的。
象细菌这样的原核类,体细胞和生殖细胞不分,细胞的分裂就是个体的增殖。
而在高等生物中,由两个配子结合而成的合子是单个细胞,单个细胞长成胚,最后长成一个成熟的个体,就是由一个细胞分成两个,两个分为4个,最后成为具有亿万细胞的个体。