真菌漆酶的研究进展
- 格式:doc
- 大小:102.50 KB
- 文档页数:13

万方数据万方数据万方数据真菌漆酶的研究进展及其应用前景作者:周雪婷, 张跃华, 罗志文, 潘亭如, 缪天琳作者单位:佳木斯大学,黑龙江佳木斯,154007刊名:农业与技术英文刊名:Agriculture & Technology年,卷(期):2012,32(9)1.王光辉;季立才中国漆树漆酶的底物专一性 19892.Nina H;Laura-Leena K Crystal structure of a laccase from Melanocarpus albomyces with an intact trinuclear coper site 2002(08)3.雷福厚;蓝虹云漆树漆酶和真菌漆酶的异同研究[期刊论文]-中国生漆 2003(01)4.李慧蓉白腐真菌生物学和生物技术 20055.Harald Claus Laccases:structure.reactions,distrihution 2004(35)6.张丽白腐真菌产漆酶对染料废水降解的研究 20047.张敏;肖亚中;龚为民真菌漆酶的结构与功能[期刊论文]-生物学杂志 2003(20)8.Gimifreda L;Xu F;Bollag J-M Laccases:a useful group of oxido reductive enzymes 1999(03)9.Xu F;Kulys J J;Duke K Redox Chemistry in Laccase-Catalyzed Oxidation of N-Hydroxy Compounds 2000(66)10.堵国成;赵政;陈坚真菌漆酶的酶活测定及其在织物染料生物脱色中的应用[期刊论文]-江南大学学报(自然科学版) 2003(02)11.缪静;姜竹茂漆酶的最新研究进展[期刊论文]-烟台师范学院学报(自然科学版) 2001(17)12.刘尚旭;赖寒木质素降解酶的分子生物学研究进展[期刊论文]-重庆教育学院学报 2001(14)13.何为;詹怀宇;王习文;伍红一种改进的漆酶酶活检测方法[期刊论文]-华南理工大学学报(自然科学版) 2003(31)14.季立才;胡培植漆酶结构,功能及应用 1996(18)15.侯红漫白腐菌Pleurotus ostreatus漆酶及对蒽醌染料和碱木素脱色的研究 200416.Huang Z Y;Huang H P;CaiR X Organic solvent enhanced spectrofluorin etric method for determition of laccase activity 1998(01)17.Badiani M;Felici M;Luna M Laccase assay by means of highperfomance liquid chromatography 1983(02)18.Wood D.A Production,Purification and Properties of Extracelluar laccase of Agaricus bisporus 1980(17)19.林俊芳;刘志明;陈晓阳真菌漆酶的酶活测定方法评价[期刊论文]-生物加工过程 2009(04)20.望天志;李卫莲;万洪文微量热法测定漆酶的活性[期刊论文]-自然杂志 1997(06)21.Kirk T K;Farrell R L Enzymatic "combustion":The microbial degradation of lignin 1987(10)22.张爱萍;秦梦华;徐清华漆酶在制浆造纸中的应用研究进展[期刊论文]-中国造纸学报 2004(02)23.Reid I D Biological pulping in paper manufacture 1991(08)24.Bergbauer M;Eggert C;Kraepelin G Degradation of chlorinated lignin compounds in a bleach plant effluent by the white-rot fungus Trametes Versicolor 1991(35)25.林建城酶在食品工业,轻工业和环境保护上的应用分析[期刊论文]-莆田学院学报 2005(02)26.林鹿;陈嘉翔白腐菌对纸浆CEH漂白废水的脱色、消除毒性和芳香化合物的降解 1996(11)27.E Rodriguez;MA.Pickard;R Vazquez-Duhalt Industial dye decolorization by laccases from ligninolytic fungi1999(38)28.Bollag J M;Myers C Detoxification of aquatic and terrestrial sites through binding of pollutants to humic substances 1992(117-118)29.Majcherczy A Oxidation of ploycyclic aromatic hydrocarbons (PAH) by laccase of Trametes versicolor 1998(22)30.刘涛;曹瑞饪漆酶在环境保护领域中的研究及应用进展[期刊论文]-云南环境科学 2005(03)31.Collins P J;Kotterman M J J;Field J A;Dobson A Oxidation of Anthracene and Benzo[a]pyrene by Laccase from Trametes versicolor[外文期刊] 1996(12)32.Ghindilis A L;Gavrilova V P;Yaropolov AI Lacease-based biosensor for determination of polyphenols:determination of catechols in tea 1992(02)33.Bauer C G;Kuehn A;Gajovic N;Skorobogatko O Holt P J Bruce N C New enzyme sensors for morphine and codeine based on morphine dehydrogenase and laccase 1999(364)本文链接:/Periodical_nyyjs201209002.aspx。

白腐真菌漆酶诱导表达的机制研究本文旨在研究白腐真菌漆酶(PLME)的诱导表达机制。
PLME可以在特定的生物体中极其有效的表达,因此引起了人们的关注。
我们对比分析了非油茄子漆酶(PNME)和西瓜漆酶(PPME)与PLME的相互作用,并讨论了PLME在诱导表达过程中的调控机制。
研究表明,PLME具有明显的诱导表达机制,其诱导表达程度与PLME的含量、时间等因素有关。
PLME可以将来自外源的信号转换成特定的蛋白质,从而实现目标基因的表达。
本文还分析了PLME诱导表达的可能机制,包括信号转导、DNA结合和调控因子等。
此外,本文还探讨了PLME的未来研究方向,以帮助今后的研究工作。
Introduction白腐真菌漆酶(PLME)是一种类似于漆酶的蛋白质,可以在特定的生物体中进行有效表达。
它表达出来的蛋白可用于各种应用,如分泌蛋白、蛋白组学和活性因子等。
目前已有很多研究表明,PLME对油茄子漆酶(PNME)的抗性更强,它可以使特定的信号在体内进行抗性表达。
因此,研究PLME的表达机制变得尤为重要。
Results and Discussion1)PLME的表达机制为了探究PLME的表达机制,我们分析比较了非油茄子漆酶(PNME)和西瓜漆酶(PPME)与PLME的相互作用。
研究表明,PLME可以显著提高PNME和PPME的表达水平(图1)。
这些结果表明,PLME具有强烈的诱导表达效果。
此外,研究表明,PLME的诱导表达程度与PLME的含量和时间有关。
当PLME含量增加时,PNME和PPME的表达水平也显著提高(图2)。
此外,当PLME处于一定时间内时,其诱导表达效果更明显(图3)。
这些结果提示,PLME具有明显的诱导表达机制。
2)PLME的调控机制鉴于PLME具有明显的诱导表达机制,我们进一步研究了PLME的调控机制。
研究表明,PLME可以将来自外源的信号转换成特定的蛋白质,从而实现目标基因的表达。
这种转换过程可分为信号转导、DNA 结合胺基酸的结合等步骤。



漆酶的研究进展及其应用作者:刘岩刘锐苏新国赵冠里杨昭来源:《安徽农学通报》2016年第13期摘要:漆酶是一种多酚氧化酶,由于其在自然界分布广泛,并且在环保、纺织、印染、食品、化学合成等方面都具有广泛的应用前景,近年来得到了广泛的关注和研究。
该文主要综述了国内外漆酶的研究进展及其应用,为细菌漆酶提供新的应用前景和方向。
关键词:漆酶;研究进展;应用中图分类号 Q814 文献标识码 A 文章编号 1007-7731(2016)13-0025-041 引言漆酶(EC 1.10.3.2)又名蓝色多铜氧化酶,可以氧化包括酚类物质、多酚类物质、苯胺、木质素、多环芳香烃甚至无机物等一系列物质,以分子氧气为电子受体,生成反应过程中唯一的副产物水。
因此,其在有毒废水处理、染料脱色、纺织、造纸、酒及饮料、生物传感器、抗癌药物及化妆品合成等方面都具有广泛的应用前景,从而受到了科学界的重视。
当前应用最多的是真菌漆酶,但由于真菌漆酶不耐高温,在碱性条件下迅速失活,存在多种抑制剂,严重限制了其工业化应用。
真菌漆酶一般为含有糖基的糖蛋白,形成了基因工程改造及异源表达上的障碍。
细菌漆酶一般为单体蛋白,且具有耐高温,在碱性条件下稳定,抑制剂少等优点,可以克服真菌漆酶应用的缺点,具有巨大的应用潜力。
2 国内外研究现状及进展漆酶为蓝色多铜氧化酶中最大的一类,具有通过铜粒子将多酚物质氧化,同时将氧气还原成水的催化特性[1]。
早在1883年,Yoshida第一次在日本漆树中发现了漆酶,成为世界上最早的被发现的酶类之一[2]。
植物漆酶由于缺少工业应用价值,而长期被忽视。
在现代工业废水中去除多酚类有毒物质,在纺织印染中去除木质素、色素等生物技术的不断研发中,由于漆酶具有利用氧气作为电子受体,能够氧化多酚类、木质素等多种化学物质,同时生成唯一的副产物水,这些自身具备的优质条件使得漆酶的催化性质在环保、纺织、印染、食品、化学合成方面具有广泛的应用前景,成为最近10年科学界最关注的焦点之一[3]。

真菌漆酶的研究进展及其应用前景摘要:漆酶生产菌株多为白腐真菌,常用的漆酶活性测定方法有分光光度法、abts法、微量热法等,其降解工业“三废”中的有毒有害物质被认为是一种效率较高,成本较低的且最有前途的方法,其对环境保护的研究以逐渐成为国内外研究的热点,本文阐述漆酶的性质、活性中心、结构特点以及其在环境治理方面的应用。
关键词:漆酶;结构;活性中心;环境修复中图分类号:x592文献标识码:a基金项目:黑龙江省教育厅科学技术研究项目资助(项目编号:12521573)为本文通讯作者漆酶最早由yoshi从日本紫胶漆树(rhus vernicifera)漆液中发现。
19世纪末,g.betranel首次将能够使生漆固化的活性物质进行分离,命名为“laccuse”,即漆酶。
漆酶属蓝色多铜氧化酶家族[1,2],与抗坏血酸氧化酶和哺乳动物血浆中铜蛋白同源。
人们将自然界中得到的漆酶分为漆树漆酶和真菌漆酶,其中真菌漆酶极具研究价值。
漆酶在生物制浆、污水处理、防腐剂、杀虫剂等化工产品的降解效果显著,用于环境保护、环境监测等领域,在食品工业等方面也有应用[3],已逐渐成为自然科学的研究热点之一。
漆酶催化氧化不同种类型的底物已达200余种,广泛用于食品、废水处理、造纸等领域。
国内外真菌漆酶研究主要是以担子菌、子囊菌、脉孢霉、柄孢壳菌和曲霉等真菌来研究漆酶的生物学活性,细菌和放线菌的研究较少,现已在细菌生脂固氮螺菌(azospirillum lipoferum)中发现了漆酶的存在。
而高等担子菌中的研究对象包括白腐真菌、杂色云芝、平菇、变色栓菌,其中白腐真菌所产的漆酶为胞外酶,可作为主要的产酶者和研究对象。
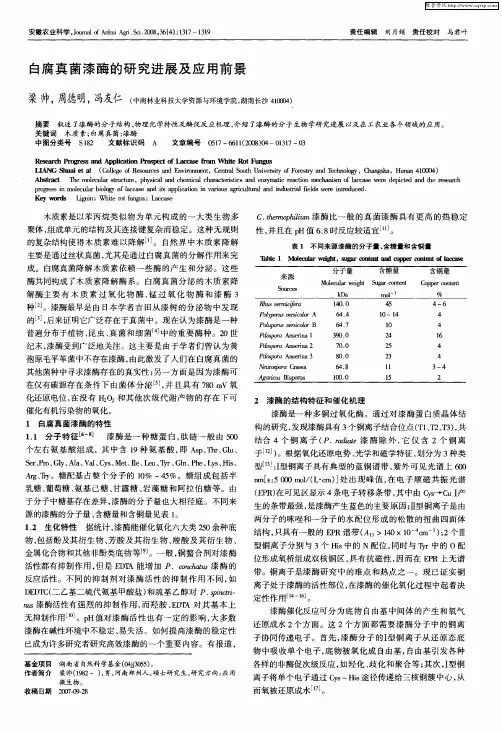
1漆酶的性质1.1理化性质漆酶是一种含铜的多酚氧化酶,不同来源的漆酶铜含量也有所不同,多含有4个铜原子[4]。
漆酶多为1条多肽链组成的单聚体,由500~550个氨基酸分子所组成,相对分子质量主要集中在50~80kd,其碳水化合物约占15%~20%,等电点pi为3~6,反应温度为30~60℃,ph低的环境,漆酶的生物活性较高[5-7]。


漆酶生产工艺的研究与改进一、引言漆酶是一种分子量较小的酶,具有广泛的应用前景。
其中最具代表性的用途就是在工业生产中作为生物催化剂,提高产物的纯度和收率,降低生产成本。
一直以来,漆酶的生产工艺一直是生物技术研究领域的热点之一,众多科学家和研究机构致力于发掘更高效的漆酶生产工艺。
二、漆酶的生产工艺目前,漆酶生产工艺主要采用微生物发酵法。
常见的产生漆酶的微生物包括铜绿假单胞菌和黄杆菌。
不同的菌株对生产条件的需求不同,需要通过不断优化来提高生产效率。
1. 菌株的选择漆酶的生产效率和菌株的选择有很大的关系。
铜绿假单胞菌和黄杆菌是目前最常被用于漆酶生产的微生物,在这些菌株中,铜绿假单胞菌的产量更高。
此外,国内外还有一些新的细菌被挖掘出来,值得进一步研究其漆酶生产的潜力。
2. 发酵条件的控制生产漆酶的过程中,发酵条件的控制至关重要。
pH值、温度、发酵时间等条件都需要严格控制。
此外,发酵过程中的氧气供应也需要控制在适宜范围内。
目前,通常采用气体补给技术和在线监测技术来保证氧气供应的平衡。
3. 收割和提取技术收割和提取技术也是影响漆酶生产效率的重要因素之一。
通常采用离心法或超滤法来收集菌体,然后使用水解酶或超声波等技术来提取漆酶。
这些方法不仅能提高漆酶的产量,还能改善漆酶的纯度和活性。
三、漆酶生产工艺的改进在不断改进的过程中,科学家们提出了很多新的漆酶生产工艺。
常见的改进方法包括:1. 基因工程改造利用基因工程技术,可以通过改造菌株的代谢途径和调节酶基因的表达来提高漆酶的产量。
研究表明,通过基因工程改造,可以将漆酶的产量提高一到两倍。
2. 大规模化生产技术目前,漆酶的生产工艺已经转移到了大规模化生产阶段。
通过使用发酵罐、气体补给器、离心机和超滤器等设备,可以在短时间内生产出大量的漆酶。
此外,使用自动化控制技术也能够提高漆酶生产效率。
3. 生化反应工程生化反应工程是现代生物技术的重要组成部分。
通过研究发酵条件、酶的稳定性和保护因子等因素,可以实现漆酶的高效生产。

白腐真菌及其漆酶的应用研究白腐真菌是一类产生漆酶的真菌,广泛存在于自然界中,能够分解植物纤维素和木质素。
漆酶是一种蛋白质,具有高度催化活性,能够氧化各种有机物质。
因此,白腐真菌及其漆酶的应用研究在多个领域具有重要意义。
首先,在生物质能源转化中,白腐真菌及其漆酶的应用得到了广泛关注。
生物质能源是可再生能源的重要组成部分,其潜在资源非常丰富。
然而,植物纤维素和木质素的高度结晶性和抗酶解性限制了生物质的高效转化。
而白腐真菌及其漆酶能够有效降解植物纤维素和木质素,促进生物质的降解和转化。
因此,基于白腐真菌及其漆酶的生物质能源转化技术可以有效提高生物质的利用效率。
其次,在环境污染治理中,白腐真菌及其漆酶的应用也具有潜力。
许多有机污染物,如农药、染料和有机废物等,对环境和人类健康造成了严重威胁。
传统的污染物处理技术通常昂贵且不具备高效环保的特点。
而白腐真菌及其漆酶通过催化氧化有机物质,能够将有机污染物降解为无毒的物质。
因此,基于白腐真菌及其漆酶的环境污染治理技术有望成为一种高效、经济和环保的处理方法。
此外,白腐真菌及其漆酶的应用还可以推动生物医药领域的发展。
近年来,特别是在抗癌药物的研发方面,白腐真菌及其漆酶被广泛用于合成生物活性化合物。
漆酶作为一种催化剂,具有选择性催化和高效的特点,能够催化各种有机合成反应,合成具有药理活性的化合物。
因此,基于白腐真菌及其漆酶的生物活性化合物合成技术极大地推动了新药研发的进程。
综上所述,白腐真菌及其漆酶在生物质能源转化、环境污染治理和生物医药等领域的应用研究具有重要的意义。
随着对可再生能源、环境保护和药物研发的需求不断增加,相信白腐真菌及其漆酶的应用将继续得到进一步的研究和开发。

产漆酶真菌以及漆酶的研究摘要:漆酶是一种含铜的多酚氧化酶,在细菌、真菌、放线菌、植物中都有报道,但报道最多的为真菌,尤其是白腐真菌。
漆酶能够降解环境中的木质素,在纸浆漂白、废水处理中意义重大。
漆酶通常情况下漆酶含有4个铜原子结合位点,通过电子在这四个铜原子间相互传递达到分解底物的目的。
漆酶基因的异源表达已经吸引了广大学者的目光,至今为止,数十种真菌的漆酶基因已经成功在酵母等异源表达系统中表达,我们可以预见漆酶在人类未来的生活中将发挥重要作用。
关键词:漆酶,白腐真菌,铜原子结合,异源表达现如今,环境污染已严重影响了人类的生存,造纸厂、纺织厂排出的废水则是重要的污染源之一。
木质素是使废水颜色发黑的主要原因[1],倘若在工厂排出废水前先将木质素出去,将会大大降低废水的毒性。
木质素是由聚合的芳香醇构成的一类物质,存在于木质组织中,是世界上仅次于纤维素的第二位最丰富的有机物。
木质素作为纤维素的粘合剂,能增强植物体的机械强度,利于输导组织的水分运输和抵抗不良外界环境的侵袭。
木质素外观上是一种白色或接近无色的物质,相对密度大约在1.35~1.50 之间,非常坚硬,难溶于各种无机或有机溶剂[2]。
木质素属天然芳香族高分子聚合物,是一类在自然界存在量很大但是又较难降解的可再生资源,木质素不含易水解的重复单元,因而不受水解酶的攻击,不仅如此,它还和半纤维素以共价键形式结合,将纤维素分子包埋在其中,形成一种天然屏障,使纤维素酶不易与纤维素分子接触,给纤维素的降解也造成了困难[3]。
因此去木质素成为目前人们面临的一大难题。
1 白腐真菌降解木质素研究表明,在自然界中,木质素的完全降解是真菌、细菌、放线菌及相应微生物群落共同作用的结果。
其中,真菌起主导作用[4],某些真菌能够有效的降解木质素,尤其是白腐真菌(white-rot fungus)可通过一系列的酶促反应实现对木质素的彻底降解。
白腐真菌是一类丝状真菌,分类学上属于真菌门,绝大多数为担子菌纲,少数为子囊菌纲。
收稿日期:2008204230;修回日期:2008211210作者简介:左斌(19862),男,2004级基础医学专业学生。
E 2mail::zbredeye2000@真菌漆酶基因研究进展左斌,王海斌,葛俊综述;贾文祥审校(四川大学华西基础医学与法医学院,成都 610041)摘 要:漆酶是一种含铜的多酚氧化酶,也是木质素生物合成的关键酶之一,目前已发现多种生物能产生漆酶,包括植物、真菌、昆虫、细菌等。
其中,以真菌中的白腐菌研究最多。
近年来,由于漆酶在生物漂白、农作物秸秆利用以及环境垃圾处理方面具有广阔的应用前景,漆酶研究越来越受到国际上的重视。
同时,随着分子生物学相关技术的发展,漆酶研究已深入基因水平,已有多种漆酶基因获得克隆,一些漆酶基因也实现了异源表达。
本文概述了真菌漆酶基因研究的最新进展。
关键词:漆酶;多酚氧化酶;木质素;异源表达中图分类号:R379文献标识码:A文章编号:100525673(2009)0120072205 漆酶(Laccase )是最简单的一种多铜氧化酶(p 2di phenol oxidase EC .1.10.3.2),一般含有4个铜原子,分布于3个高度保守的不同结合位点,每个铜原子在催化机制中都有很重要的作用。
漆酶能催化O 2通过4个电子还原成水,并且伴随着一些酚类底物的氧化。
漆酶最早是由日本的吉田1883年在漆树的分泌物中被发现,随后Laborde (1893)又证实在一些真菌中也含有这种酶。
目前研究发现,漆酶不仅广泛存在于真菌和植物中,而且某些昆虫、细菌也能分泌漆酶〔2〕。
其中对属于白腐菌类的担子菌研究最多。
根据漆酶的来源主要分为漆树漆酶(Rhus Laccase )和真菌漆酶(Fungal laccase )两大类,不同来源的漆酶核苷酸序列差异很大〔3〕,本文综述了真菌漆酶基因克隆与表达的最新进展。
1漆酶基因的克隆1988年Fr oh man 等发明快速扩增c DNA 技术(RACE 技术),随后该方法被广泛用于漆酶基因的克隆〔2,3〕。
真菌漆酶的研究进展宋瑞(安徽大学生命科学学院合肥230039)【摘要】漆酶是一种蓝色多铜氧化酶,和植物中的抗坏血酸氧化酶,哺乳动物的血浆铜蓝蛋白属同族,能够催化多种有机底物和无机底物的氧化[1,2],同时伴随分子氧还原成水。
漆酶广泛分布于真菌、高等植物、少量细菌和昆虫中,尤其在白腐真菌中普遍存在。
漆酶特有的结构性质和作用机理使其具有巨大的应用价值。
本文就真菌漆酶结构,功能的研究进展作一综述,并对其应用作简单介绍。
【关键词】真菌漆酶三维结构功能应用1真菌漆酶结构特征1.1 漆酶的组成漆酶是一种糖蛋白,肽链一般约由500个氨基酸组成[3],糖基含量差异较大,占整个分子质量的10%—80%[4],据相关报道,漆酶的热稳定性可能与其糖基化有关。
糖组成包括半乳糖、葡萄糖、甘露糖、岩藻糖、氨基己糖和阿拉伯糖等。
Mayer[5]认为漆酶并不均一,它由多条5000~7000分子量的糖肽链基本结构单元组成。
由于结构单元之间的缔合度不同,造成了各种漆酶分子量的不同。
另外,分子中的糖基的差异,也会引起漆酶的分子量随来源不同会有很大的差异,从59—390ku不等。
真菌漆酶约含19种氨基酸,绝大部分为单体酶,但也有例外,如双孢蘑菇和长绒毛栓菌漆酶由两个亚基组成[6],而柄孢壳漆酶I由四个亚基组成。
漆酶种类繁多,不同种类的真菌产生的漆酶种类不同,即使同一种真菌在不同环境下也产生不同种漆酶。
1.2漆酶的晶体结构由于漆酶是含糖蛋白质,且糖质量分数较高,一直以来很难获得X-衍射分析所用的单晶体,因此阻碍了关于漆酶结构的研究进展。
1998年第一个漆酶晶体是Ducros V[7]制备的来自灰盖鬼伞(Coprinus cinereusv)T1Cu缺失型漆酶晶体,并分析了其结构。
至今为止,Bacillus subtilis(CoA)[8];Melanocarpus albomyces(MaL)[9];Rigidoporus lignosus(RiL)[10];Pycnoporus cinnabaricus(PcL)[11];Coprinus cinereus(CcL)[12]和Trametes versicolor(TvL)[13]漆酶的三维结构已相继被报道。
漆酶分子整体由3个杯状结构域所组成,分别称作结构域A、B、C,每个结构域主要由β-折叠桶,α-螺旋,loop结构所组成。
三者紧密结合形成球状结构。
这是铜蓝蛋白家族所共有的结构形式[7,9]。
分子当中含有二硫键,漆酶种类不同,二硫键数目也不一样,MaL 漆酶分子由3个二硫键,分别是位于结构域A Cys4~Cys12、结构域A和C界面上Cys114~Cys540、结构域C Cys298~Cys332,而CcL,RiL漆酶中则含有两个二硫键。
在CcL漆酶分子中,由结构域A的Cys85和结构域B的Cys487形成一个二硫键,另一个二硫键存在于结构域A和结构域B(Cys117—Cys204)之间。
一个伸展的loop(氨基酸284—327)连接结构域B和结构域C。
Asn343上有N连接的N—乙酰葡萄胺。
1.3 漆酶的催化中心真菌漆酶分子中一般都含有4个Cu原子,根据磁学和光谱学性质不同可将4个Cu原子分为三类:Ⅰ型Cu2+(T1Cu)和Ⅱ型Cu2+(T2Cu)各一个,都是单电子受体,呈顺磁性;Ⅲ型Cu24+(T3Cu)两个,是耦合离子对,是双电子受体,反磁性[14]。
T1Cu位于结构域C上,T2Cu 和T3Cu位于结构域A和C的界面上。
漆酶绝大多数都只含4个Cu 原子,Karhunen E等[15]研究的射脉菌( Phlebia radiata)漆酶却只含有2个Cu原子(T1Cu、T2Cu各一个),无T3Cu原子,但该漆酶分子中含有1个具有类似T3Cu功能的有机小分子辅基吡咯喹啉(pyrroloquinolinequinone,PQQ)。
Palmieri G等[16]从糙皮侧耳(Pleurotus ostreatus)中纯化的一种新型漆酶POXA1中含有1个Cu原子,2个Zn原子和1个Fe原子,由于缺少T1Cu,在610nm处没有吸收峰,酶蛋白呈白色。
在不同种真菌漆酶分子中,与Cu原子相连的1个Cys 和10个His及其周围的氨基酸配基则是相对保守的,这些氨基酸都分布在含有两对His的N末端区域,或是分布于C末端的Cys和其余结合铜原子的His之间。
Cu离子构成了漆酶的活性中心,在漆酶催化氧化过程中起决定性作用,如果除去Cu离子,漆酶将失去催化功能。
1.3a T1Cu结合位点T1Cu形成单核中心接受来自底物的电子转移,具有典型的蓝铜谱带,紫外可见光谱上600nm处有吸收峰,它是底物反应场所,把来自底物的电子传递三核中心铜原子上。
T1Cu以Cu2+形式存在,在所有的漆酶中都与两个His的N原子和一个Cys的S原子配位结合,形成扭曲的四面体结构。
有些多铜氧化酶还有第四个轴向的氨基酸配基,在不同的多铜氧化酶中配基种类不同,在抗坏血酸氧化酶中为Met,大多数其它真菌漆酶中通常为Phe,而灰盖鬼伞漆酶中为Leu,距离T1Cu 3.51Å,因此不能成为T1Cu的配基。
这种配位的不同被认为可能引起真菌漆酶氧化还原电位升高。
Perrycr等[17]认为此位置的还原电势强弱与其疏水基团大小成正比,如为甲硫氨酸(Met)配基,亮氨酸(Leu)配基,苯丙氨酸(Phe)配基,其还原电势依次升高。
然而Silvia Garavaglia等[10]通过比较CcL(低Eo)和TvL(高Eo)的不同发现,Leu或Phe不同的配位并不是导致氧化还原电位变化的最主要因素,认为T1Cu-NE间的距离则是影响氧化还原电位高低的主导因素。
围绕在T1Cu一侧的是一个疏水性口袋,Leu462两边分别为Phe340和Phe398。
T1Cu存在于Ile454的一侧,且位于一个约12Å宽、6Å深的凹陷中,结构域A和结构域C的三个loop构成了凹陷的表面。
这个区域与还原底物的结合有关。
这些伸展的loop区,决定了底物特异性,而真菌漆酶具有极度广泛的底物专一性,这可能是由于漆酶中缺少这些区域,序列变异性也较大,且结合位点相对较大,再加上loop 构象上存在的变化(表现在室温和100K结构之间的不同)的影响。
1.3b T2/T3Cu结合位点T2Cu与T3Cu形成三核中心接受来自T1Cu单核位点的电子,与高度保守的四个His-X-His花样的8个His配位结合,T2Cu结合2个,每个T3Cu结合3个[8,10]。
两个T3Cu之间结合漆酶的第二个底物氧分子。
T2Cu具有电子顺磁共振效应,不易结合弱的配体,且不稳定,当螯合剂存在时,能够被一些2型耗竭(T2D)衍生物选择性还原去除;T3Cu的两个铜原子间偶联,其EPR性质消失,由于电子在OH和两个T3Cu之间的传递,在330nm处有特征吸收带。
另外,在400~460nm间有一些小吸收带,推测是一些反应中间体,如氧/过氧中间体(PI)[18,19,20]。
T2Cu与2个His和一个水分子或一个氯原子配位,形成T型几何结构[21],这种结构对一价铜比二价铜更有利。
在灰盖鬼伞漆酶晶体结构中,由于去除糖链导致T2Cu缺失,与抗坏血酸氧化酶相比,其构象发生些许变化。
T3Cu对的两个Cu原子相距为5.3Å,而后者只有3.7Å;含氧的配基只结合T3Cu原子对中的一个,而且它在两个Cu原子间是不对称排列的,相距Cu3(a)和Cu3(b)分别为2.17Å和3.26Å;另外与其相似的是T2D漆酶(T2Cu 被螯合,T1Cu和T3Cu保持完整),在T2D漆酶中,与T2Cu配位的His399的咪唑环发生了旋转,因而能与T3Cu原子对中的一个Cu(距离2.5Å)配位;His399存在构象柔性,在1.68Å,100K结构中有两种构象,一种与抗坏血酸氧化酶和血浆铜蓝蛋白中的构象相对应,另一种是与T3Cu(a)配位的新位置。
在100K时,两种构象比例为60:40,而在2.2Å室温结构中比例为80:20。
总之,T3Cu(b)与His109、His66、His453配位,形成一个轻微扭曲的三角平面,距离氧桥3.26Å,距离T3Cu(a)5.1Å。
T3Cu(a)有两种配位状态,一种是与His451、His111、His401的N和氧桥配位,距离分别为1.98、2.01、2.04和2.17Å,犹如一个扭曲的四面体。
另一种是Cu3(a)与旋转进入配位区的His399形成第五个配位,距离2.5Å。
因此,T2D漆酶有两种构象,其平衡受温度变化和结晶条件的影响。
不同来源漆酶的T2/T3Cu结合位点也有所差异,在Rigidoporus lignosus(RiL)中T3Cu的2个铜原子间无OH桥存在,相反T2Cu,T3Cu(a)则分别与1个OH相连,但由于与OH是不对称相连,所以不能认为是OH桥。
然而,Bacillus subtilis(CoA)T3Cu的2个铜原子则是通过1个OH桥配位连接起来,组成双核铜区,具有抗磁性,因而在EPR上无信号产生。
Silvia Garavaglia等[10]认为在Rigidoporus lignosus(RiL)中,T3Cu(a)、T2Cu分别与1个OH相连是导致EPR沉默的原因,具有EPR活性的是与来自3个His的N原子相连的T3Cu(b),而不是T2Cu。
T2和T3中心紧密相连形成一个三核中心,其结合方式可能有以下三种模式:(1)三个同原子族是通过羟基和/或过氧基桥链连接,早期的报道均持这种观点;(2)通过双氧(过氧)连接,两个氧原子到三个铜原子的距离几乎一样,均在2.4~2.6Å之间,同两个T3Cu呈(扭曲)四面体,同T2Cu呈平面四方构型,Hakulinen等的分离态[22]和重组态[23]以及Bento等[24]的CuCl2浸泡态结构支持这种模型,(3)三个铜都同一个µ3-氧连接,另一个氧原子已经转变成水离去[25],这可能是反应过程中的一种状态。
1.3c 活性中心结合方式T1Cu距离三核中心约为1.25nm,通过T1Cu-Cys-His-三核中心联系起来,形成电子传递通道。
Leif J[26]等人研究T. versicolor活性中心时发现其含有2个二硫桥:Cys117与Cys205形成一个二硫桥,把T1Cu 区和T2Cu区联系起来;Cys85与Cys487形成另一个二硫桥,把T1Cu区与T3Cu区联系起来。
从空间结构上看,T1Cu可能距离两个T3Cu 约13Å[27],而T2Cu到两个T3Cu的距离可能略微不等,T3Cu原子对间的距离约为 4.8Å。