5碳酸盐台地沉积体系
- 格式:ppt
- 大小:33.95 MB
- 文档页数:73


碳酸盐岩层序地层模式一、碳酸盐岩层序地层学概述Vail 等于1987 年提出层序地层学( sequence stratigraphy ) 的概念,在1988 年, Van Wagoner 又提出了新的定义: 研究以侵蚀面或无沉积作用面或可相互对比整合面为界的年代地层学格架中有成因相关意义的岩相的科学。
层序是最基本的单元,一个层序又可分为若干个体系域,体系域内部是个或多个准层序或准层序组。
准层序即是在成因上具有联系的、相对整一的一套岩层或岩层组。
确定层序所必需的许多信息来源于地震资料(当然也可以从露头和井孔中获得)。
除了对早期的硅质碎屑岩讨论以外,对碳酸盐岩层序地层学的讨论也是有用的,因为这样可以唤起对沉积倾向性的理解。
所有的沉积体系都记录了相同的基本过程的影响,但它们的记录又各有所侧重,地质学家应该了解沉积体系在记录海平面变化、气候或其它环境因素中的倾向性,这样便能更加全面地对层序地层学进行学习和研究。
碳酸盐体系不同于硅质碎屑体系. 碳酸盐沉积物是在盆内形成的,因此除了区域构造沉降和海平面变化外,海洋气候和水文条件也控制着碳酸盐的沉积过程。
J. S. Sang在对世界不同碳酸盐台地研究后认为:短期的全球海平面变化( 相对海平面变化)是控制碳酸盐生产率、碳酸盐台地或碳酸盐滩发育及其相分布的主要因素。
1、海相碳酸盐岩——生物礁层序地层学研究目前,涉及我国新生代、中生代和古生代海相碳酸盐岩—生物礁油气层序地层学最为突出的几个成功研究实例是四川普光、新疆塔里木和南中国海油气区,获得了油气突破性发现。
首先研究涉及层序单元划分与基本特征、主要层序界面识别、层序单元划分等内容, 完成了上第三系生物礁——碳酸盐岩层序地层格架、典型生物礁储层层序格架、层序界面或单元划分、层序地层格架下的沉积体系域特征与沉积体系控制因素等研究。
2、碳酸盐岩沉积及层序发育的主要控制因素大地构造作用决定碳酸盐沉积作用背景, 大地构造背景下的盆地结构是影响碳酸盐岩层序几何形态的一个关键因素。

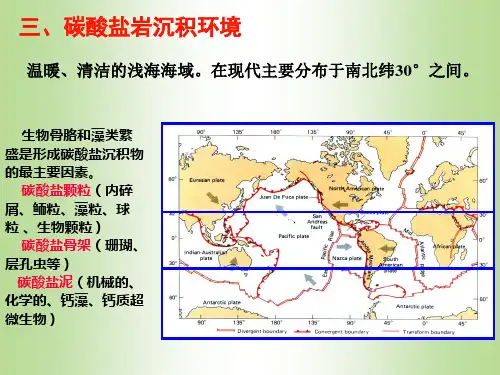

碳酸盐沉积环境碳酸盐沉积环境海洋碳酸盐沉积环境现代碳酸盐岩的分布特征分布地带:碳酸盐沉积主要分布于低纬度(南北纬30o左右)的清澈、温暖、滨浅海地带条件:浅水、暖水、清水、阳光充分、没有大量细碎屑沉积物的注入。
生物:钙藻大量繁殖,珊瑚礁发育。
沉积物:主要是两类沉积物(1)颗粒碳酸盐(贝壳砂、鲕粒砂、葡萄状团块、球粒);(2)造礁生物粘结岩。
少量灰泥在南北纬40o之间的深海盆地底部,有大量浮游生物碳酸盐沉积。
浅海碳酸盐的发育与藻类有密切关系在水深15m中所产生的CaCO3比深陆缘海每单位面积的CaCO3多几倍。
主要与浅水绿藻及蓝绿藻特别丰富有关。
由于藻类的光合作用,从海水中吸收大量CO2,从而促使海水中的CaCO3过饱和而沉淀出文石质灰泥,而且钙藻的外壳也是文石质灰泥(成为颗粒的主要供给者)。
藻类繁盛提供了大量碳酸盐沉积物。
浅海碳酸盐的发育与生物有密切关系藻类的生活需要温暖、浅水、清洁透光环境。
海水浑浊妨碍光合作用,阻止钙藻生长,堵塞底栖生物的摄食器官,影响其繁衍(妨碍了大量碳酸盐颗粒的产生)。
海水太深,阳光和氧气不足,对藻类和底栖无脊椎动物生长都不利。
海水太深,水压大,溶解CO2多,CaCO3不饱和,因此深水不会有大量碳酸盐的产生。
深水碳酸盐沉积物主要靠海水表层浮游生物(颗石藻、有孔虫、翼足类等)和浅水陆棚区漂运来的灰泥或粉屑。
浅海碳酸盐颗粒的复杂成因内(源)碎屑:盆地内准同生改造的碳酸盐颗粒。
内(盆内):直接来源与准同生改造;成分:碳酸盐。
在海岸高能带,由于波浪、潮汐、海流等作用,使碳酸盐沉积物发生簸选,将细粒碳酸盐带走,而留下各种砂砾级碳酸盐颗粒,形成各种砂砾屑滩、介壳滩、沿岸砂坝、砂咀、滨外砂堤、砂洲、潮汐三角洲、潮汐砂坝等(西沙群岛)。
细粒碳酸盐(灰泥、粉屑)沉积在:(1)较深水盆地区:陆棚边缘、障壁砂坝前缘的较深水区(滩前、滩间)。
(2)较低能的浅水区:障壁后的泻湖及潮坪区。
碳酸盐与生物和生物礁碳酸盐沉积物主要是生物成因的。

沉积相知识汇总1.朵状三角洲:是指三角洲前缘的指状砂体受到海水的冲刷、改造,再分配而形成席状砂层,使三角洲前缘变得较为圆滑而近似于半圆形的三角洲。
2.鸟嘴状三角洲;是指当海洋的波浪作用大于河流的作用时而形成的平面形态呈鸟嘴状的三角洲。
3.建设性三角洲;是指在以河流作用为主,泥砂在河口堆积的速度远大于波浪所能改造的速度的条件下形成的三角洲。
其特点是增长速度快,沉积厚、面积大,砂/泥比值低。
4.破坏性三角洲;是指在海洋作用增强而超过河流作用时,河口区形成的泥砂堆积被海洋水动力改造,破坏时形成的三角洲。
其特点是形成时间短,分布面积小。
5.港湾型三角洲;是指河流流入三角港或其他形状的港湾所堆积的泥砂被潮汐作用破坏和改造,外形受港湾控制的三角洲。
6.扇三角洲;是指从邻近高地直接推进到海、湖等稳定水体中的冲积扇。
7.深水三角洲;是指曲流河注入水体深度一百米左右的蓄水盆地所形成的三角洲沉积体。
8.浅水三角洲;是指曲流河注入水体,深度一般几米~几十米的蓄水盆地所形成的三角洲沉积体。
9.三角洲平原;是指从河流大量分叉位置至海平面以上的广大河口地区,是三角洲沉积的陆上部分。
10.三角洲前缘;是指位于三角洲平原向海方向一侧,处于海平面以下的河流与海水剧烈交锋的河口地区。
11.分流间湾;是指分流河道间相对低洼的与海(湖)相连的海湾地区。
12.远砂坝;是指位于河口坝前方较远部位,沉积物较河口坝细的主要为粉砂,并有少量粘土和细砂组成的沉积体。
13.三角洲复合体;由于河流与海洋作用的消长以及河口的往返迁移,三角洲的成长、废弃多次重复出现,形成多个单一的三角洲沉积体交错叠置,称为三角洲复合体。
14.辫状河三角洲;是指由冲积扇前的辫状河注入蓄水盆地所形成的沉积体。
15.曲流河三角洲;是指曲流河注入蓄水盆地形成的沉积体。
16.浅水湖泊三角洲;是指河流注入广阔的滨浅湖区形成的沉积体。
17.澙湖;是指滨浅海地带由于障壁地形的遮挡而形成的与广海呈隔绝或半隔绝的水域。
碳酸盐台地沉积模式分类及主控因素1 前言碳酸盐岩地模式是理解碳酸盐岩相的分布以及(在一定程度上)原生孔隙率的分布及其成岩史有关的保存情况的重要辅助工具。
随着油气勘探的不断深入, 海相碳酸盐岩地层分布区逐渐成为重要的油气勘探场所。
中国的塔里木盆地、四川盆地和鄂尔多斯盆地都有重大的发现, 特别是塔里木盆地、四川盆地最近都有重要的突破, 因此, 碳酸盐岩台地的研究也进入一个新的高潮, 各种观点、方法层出不穷。
通常用来描述不同台地的许多术语,对于地质学家来说往往具有不同的含意。
本文根据前人对碳酸盐台地的研究总结归纳其类型划分及其相模式。
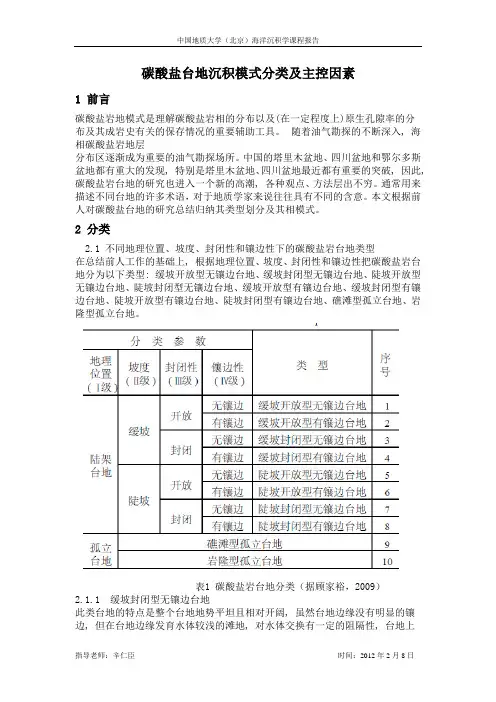
2 分类2.1 不同地理位置、坡度、封闭性和镶边性下的碳酸盐岩台地类型在总结前人工作的基础上, 根据地理位置、坡度、封闭性和镶边性把碳酸盐岩台地分为以下类型: 缓坡开放型无镶边台地、缓坡封闭型无镶边台地、陡坡开放型无镶边台地、陡坡封闭型无镶边台地、缓坡开放型有镶边台地、缓坡封闭型有镶边台地、陡坡开放型有镶边台地、陡坡封闭型有镶边台地、礁滩型孤立台地、岩隆型孤立台地。
表1 碳酸盐岩台地分类(据顾家裕,2009)2.1.1 缓坡封闭型无镶边台地此类台地的特点是整个台地地势平坦且相对开阔, 虽然台地边缘没有明显的镶边, 但在台地边缘发育水体较浅的滩地, 对水体交换有一定的阻隔性, 台地上主要以潮坪沉积和澙湖沉积为主。
在潮上带发育膏泥坪、膏盐坪和含膏白云岩等, 而在潮间带主要沉积以泥晶为主的云岩、藻云岩和少量的藻屑云岩, 下部见砂屑坪沉积等; 澙湖中以泥晶白云岩和纹层白云岩等为主; 台地边缘滩中见凝块白云岩和少量的鲕粒白云岩。
2.1.2 陡坡开放型无镶边台地这是一类比较简单的台地。
台地岸坡坡度较陡, 滨岸水动力强度较高, 滨岸沉积主要是粒级较粗的生物碎屑灰岩、生物灰岩或一定量的鲕粒灰岩, 在一些地区岸边还发育一定高度的风成沙丘;向外则是一个相对有一定坡度的开阔台地, 沉积物主要是泥晶灰岩和泥质灰岩, 沉积物由岸至海逐渐变细, 直到台地边缘出现更大坡度的斜坡。

第十三章碳酸盐台地沉积环境第一节沉积环境类型和特征碳酸盐台地:原指地形平坦的浅水碳酸盐沉积环境。
现指所有碳酸盐沉积环境(不考虑地形是否平坦)。
包括潮坪、生物礁、浅滩、局限台地、开阔台地等一、潮坪指潮汐周期性影响的岸边平坦沉积环境1、潮上带1)位置:平均高潮线以上,低能环境2)岩性:灰泥和泥晶白云岩,层薄色浅;潮湿环境多藻席,干旱环境多石膏3)构造:泥裂、鸟眼,水平纹层和叠层石2、潮间带1)位置:平均高、低潮线之间,较低能环境2)岩性:灰泥和颗粒灰岩,层薄色浅;柱状和波状叠层石3)构造:交错层理、生物扰动、水平纹理及泥裂3、潮下带1)位置:平均低潮线之下,低能或高能环境2)低能潮下带(小潮差):暗色颗粒质灰泥岩,强烈生物扰动,层理不发育,多正常海相化石3)高能潮下带(中大潮差):厚层颗粒灰岩,分选、磨园好,多波痕和交错层理二、生物礁和浅滩1、生物礁2、浅滩是指沉积水体浅、能量高、沉积颗粒灰岩为主的沉积环境。
包括:裙滩、堤滩、点滩和台缘滩2、浅滩1)裙滩(岸滩)(1)位置:位于地形坡度较大的无泻湖的海岸地区,波浪作用较强。
(2)岩性:中厚层亮晶颗粒灰岩,颗粒分选磨园好,发育交错层理。
2)堤滩(障壁滩)(1)位置:位于有泻湖的海岸地区,外侧为开阔海,呈堤状障壁平行岸线延伸(2)岩性:中厚层亮晶颗粒灰岩,颗粒分选磨园好,发育向上规模变小的交错层理。
3)点滩(1)位置:零星分布于台地内部的、与局部水下隆起相关的浅滩(2)岩性:中厚层亮晶颗粒灰岩,颗粒分选磨园好,发育交错层理、波痕。
4)台缘滩(1)位置:分布于台地边缘、浅水高能环境(2)岩性:厚层块状亮晶颗粒灰岩,颗粒分选磨园好,发育交错层理、平行层理和波痕。
三、局限台地1、位置:指海水循环受限、盐度不正常的低能浅海,与广海之间有障壁2、岩性:暗色厚层灰泥灰岩,缺少层理构造,较强生物扰动四、开阔台地1、位置:指海水循环好、盐度正常、发育礁滩的浅海2、岩性:中厚层颗粒灰岩、灰泥灰岩,多正常海相化石,生物扰动多五、台地边缘1、位置:指浅水台地与深水斜坡相邻的沉积区,能量较强。


延长探区天然气气藏特征以及成藏的控制因素摘要:伴随着社会生产生活水平的逐步提升,天然气已逐渐成为人们日常生活中必不可少的一部分,天然气是一种较为安全的燃气,它与其他燃气存在一定的区别,天然气气藏具有其独有的特征,为了对天然气形成及储藏等因素有更为深入的了解,本文以某地区为例对延长探区天然气气藏特征及成藏控制因素进行了分析与研究。
关键词:延长探区天然气气藏特征成藏控制因素分析天然气是一种清洁型能源,符合我国环保理念,因此在人们生活中得到了广泛应用,它在很大意义上提供了我国国民生活质量。
基于天然气强大的功能性及优势,现今人们对天然气的需求量与日俱增,在这一形势下建立天然气气藏,为人们储藏并提供充足的天然气刻不容缓。
一、延长探区实际概况本文在延长探区天然气气藏特征及成藏控制因素分析中之所以选取该地区为例,是因为该地区具有得天独厚的天然气气藏条件,符合天然气气藏对地质的基本要求。
该地区分属盆地,盆地周围是广为辽阔的区域,具有多处适合天然气气藏聚集的区域。
二、延长探区天然气气藏特征该地区地域辽阔,具备丰富的天然气气藏地理优势,为了对延长探区天然气气藏特征,本文选取该地区三处适合建设天然气气藏的区域,以下是对这三处地质特征的具体分析:1.目的层沉积相特征该地区目的层沉积相经历了古地理格局及近岸碎屑湖盆、陆表海的演变历史,根据平面沉积相带及纵向沉积序列的分布状况可将目的层划分为五大体系,分别是海岸沉积体系、碳酸盐台地沉积体系、辫状河三角洲沉积体系、曲流河三角洲沉积体系及湖泊沉积体系。
1.1海岸沉积体系。
海岸沉积体系在整个目的层沉积相中比较发育,该体系的区域分布主要包括泻湖一障壁岛及潮坪两个亚相,泻湖一障壁岛沉积与潮坪沉积微相是由障壁海岸沉积延安南泥湾一带发育而成。
1.2碳酸盐台地沉积体系。
碳酸盐台地沉积体系是由台地亚相沉积发育形成,在沉积发育的过程中可见到的碳酸盐台地沉积微相类型主要有生物灰岩微相、生物碎屑灰岩微相、泥晶灰岩微相。

中国科学 D 辑 地球科学 2005, 35 (7): 627~637 627桂林中、晚泥盆世微生物碳酸盐沉积、礁 和丘及层序地层、古环境和古气候的意义*沈建伟**(中国科学院南海海洋研究所和广州地球化学研究所边缘海重点实验室, 广州510301)毛家仁(贵州工业大学资源与环境学院, 贵阳 550003)摘要 钙质碳酸盐的微生物沉淀作用从太古代开始一直在碳酸盐台地的发育中起到了至关重要的作用. 微生物碳酸盐在重大的环境变化中(包括台地环境的海侵事件和生物绝灭后的复苏阶段)是碳酸盐沉积体的重要贡献者; 在层序地层不同体系域中有不同的微生物形成的碳酸盐沉积.桂林晚泥盆世法门期碳酸盐台地中的生物礁和丘完全由微生物和胶结物形成, 代表了弗拉-法门期生物事件后骨骼后生生物完全缺失的礁生态系中微生物为主的、独特的生物礁类型; 它们与特定的古环境和古气候有关, 可以用作研究环境变化和生物事件的指标. 关键词 微生物碳酸盐沉积 层序地层 古环境 古气候 晚泥盆世 桂林2004-04-28收稿, 2004-10-15收修改稿 *中国科学院“百人计划”项目资助** E-mail: jwshen@碳酸盐台地和礁环境中的钙质微生物(Calcimicrobes)及其沉积物——微生物石(Microbialite)是最近提出的一个关于微生物群落(Microbial com-munity)成因的凝块、纹层和非纹层构造的术语.微生物石是由底栖(原核或真核)微生物群落加积并捕获和粘结碎屑物质的沉积物[1]. 在地质时期和现代生物礁及碳酸盐沉积体中都有踪迹, 但它们与古气候和古环境的关系近年来才被认识和讨论[2~4]. 实际上它们在生命的起源、进化和绝灭, (古)气候和(古)环境的变迁中都起到了很重要的作用. 微生物群落由于自身的代谢多样性可以适应和殖生所有的生境. 在地质时期的碳酸盐和生物礁沉积中, 钙质碳酸盐的微生物沉淀作用非常重要[5]. 最近20多年的研究表明, 微生物石在现代生物礁环境中扩散非常迅速, 研究已发现微生物群落与珊瑚礁的毁坏有联系, 它们的繁盛和广布会导致珊瑚礁生长的衰减, 因此, 微生物石628中国科学D辑地球科学第35卷可以用做环境变化的指标. 研究还证实微生物石对季节的变化很敏感, 它们在湿热的季节里生长和发育得更快, 在一些现代礁系统中海面温度的升高会刺激微生物石的生长, 在这一点上微生物石可以用作气候变化的指标1). 微生物石还在礁系统的生物地球化学循环中起作用, 它们直接生长在珊瑚体之上而导致光和矿物质可得到量的减少, 引发珊瑚丛体的黑斑病[6].弗拉-法门期生物绝灭事件对许多海洋生物组合, 特别是造礁生物影响重大, 礁生态系中的骨骼后生生物在全球范围内受到影响, 表现为造礁层孔虫和珊瑚在弗拉-法门界线附近消失贻尽. 晚泥盆世法门期一般被认为是生物礁发育的贫困期, 没有真正的骨骼后生生物骨架. 然而, 微生物类和藻类作为这一时期碳酸盐沉积的主要贡献者已在世界泥盆系的主要发育区得到了证实, 如中国华南[5,7], 加拿大[4,8,9]和澳大利亚[10~14]. 由微生物类和藻类进行的造礁活动持续至法门期. 弗拉期层孔虫-珊瑚-钙质微生物的礁体被法门期几乎全部由钙质微生物和它们的沉积物形成的礁体所替代. 生物礁的发育在华南的桂林和西澳大利亚的坎宁和波拿巴特盆地持续穿过骨骼后生生物灭绝的界线[5,11,12,14]. 现在普遍认为微生物碳酸盐和生物促生的胶结物是地质历史中生物礁骨架的重要贡献者, 它们在形成礁骨架方面的作用等于或甚至超过骨骼后生生物[2]. 本文对桂林晚泥盆世法门期的微生物礁和丘(图1)的生物组成、岩性特征和礁穴类型进行描述, 对微生物碳酸盐沉积在桂林上泥盆统碳酸盐台地层序地层格架中的产出形式进行论证, 并对它们与古环境、古气候、生物事件和海平面变化的关系进行讨论.1 桂林泥盆系生物礁组合中泥盆世晚期(吉维期)桂林及其临近地区从碎屑沉积的陆架发展成碳酸盐台地, 其是广西中部和南部广阔碳酸盐陆架(象州台地型)的一部分. 切割沉积基底的主断裂是控制沉积盆地分异的主要因素.图1 研究区的地理位置和桂林晚泥盆世法门期微生物礁和丘的出露点1) Science News, 2002. Microbial linked to coral reef decay. 第7期沈建伟等: 桂林中、晚泥盆世微生物碳酸盐沉积、礁和丘及层序地层、古环境和古气候的意义629华南泥盆系特有的台凹相沉积是这些主断裂活动的结果, 这些台凹相沉积往往分离广阔的碳酸盐陆架并形成很多次级的台地. 在桂林地区, 灵川和绕山断裂把桂林的碳酸盐陆架分割成相对小的桂林、岩山和阳朔亚台地(subplatforms). 碳酸盐台地为造礁生物(钙质骨骼生物、钙质微生物和藻类)提供了适宜的生境. 泥盆系造礁生物层孔虫和珊瑚属级分异的高峰发生在艾菲尔期至吉维期; 而钙质微生物和藻类长时期与层孔虫和珊瑚共同造礁, 并在法门期独立成为主要的造礁生物, 钙质微生物构成了桂林法门期独特的生物礁. 在桂林泥盆系礁组合中, 微生物碳酸盐沉积从吉维和弗拉期礁前斜坡的叠层石和核形石变化为法门期台地边缘和斜坡的微生物礁和丘和台地内部的微生物席状沉积[7].桂林吉维期钙质微生物礁格架主要由微生物沉积结构Spongiostroma(图2(a)), 微生物Rothpletzella (图3(a))和同沉积胶结物形成. Girvanella和疑源微生物碳酸盐主要产出在礁后相和局限台地相, 或者和Rothpletzella一起在礁前斜坡相形成叠层石. 弗拉期的钙质微生物类和藻类的造礁作用比以前重要, 但是, 它们和吉维期的钙质微生物类和藻类一样主要分布在礁前斜坡相. 然而, 在弗拉最晚期, 微生物在台地边缘形成生物礁和丘, 其中可见复体珊瑚. 微生物(Renalcis, Keega-like microbes, Rivularia, Ortonella, Rothpletzella)、藻类(Parachaetetes, Solenopora)、核形石-叠层石和核形石-凝块叠层石是弗拉期微生物碳酸盐沉积的主要贡献者[7].2 桂林法门期的钙质微生物礁桂林晚泥盆世法门期微生物礁骨架由Epiphyton, Renalcis和其他微生物构成. 它们与鲕粒滩、腕足类介屑滩和头足类滩一起形成法门期碳酸盐台地边缘相, 这明显与吉维期和弗拉期的台缘相珊瑚-层孔虫礁和层孔虫-珊瑚-微生物礁不同.2.1 Renalcis-Epiphyton礁和丘法门期Renalcis-Epiphyton礁出露在桂林城西约9~10 km寨江村旁的一座孤山(图1). 礁发育在法门期上斜坡沉积上. 出露的礁体有点呈楔状的丘形, 直图2 微生物沉积结构和骨架(a) 微生物沉积结构Spongiostroma; (b) 丛状Epiphyton形成的骨架(光片, 比例尺每格代表1 mm)630中国科学 D 辑 地球科学第35卷图3 微生物粘结结构(a) 微生物Rothpletzella 形成叠层生长形式包粘黑色的灰泥沉积物; (b) 微生物Epiphyton 呈丛状向上叠生生长,白色部分是同沉积胶结物径约50 m, 最大厚度在礁体的西缘, 达35 m. 礁体由各种类型、不显层理的微生物格架组成. 微生物类主要有: Epiphyton , Renalcis 和Izhella , 其他类型包括Palaeomicrocodium, Rothpletzella, Wetheredella 和肾形的micropromblematica [5]. Epiphyton 是最主要的造礁微生物, 呈分枝丛状(图2(b)), 但也有肾形的丛状. Epiphyton 常见的生长方式是丛体叠生在丛体之上呈 “V”型向上展开生长(图3(b)), 表现出重要的造架、障积和粘结的作用. Renalcis 和Izhella 的数量和造礁作用相对较小, 但通常和Epiphyton 和Palaeomicroco - dium 生长在一起. Rothpletzella 和Wetheredella 主要形成微生物粘结岩. 附礁生物主要是有孔虫、腕足类和海百合茎, 它们多呈碎屑产出. 礁体中也发现有零星的海绵[5].寨江微生物礁的微相包括: (1) 丛状的Epiphyton 骨架岩; (2) 肾形状的Epiphyton 骨架岩; (3) Renalcis- Epiphyton 骨架岩; (4) Renalcis 骨架岩; (5) Microprob-lematica 生物粘结岩; (6) 疑源微生物-Rothpletzella 粘结岩; (7) 球粒和生物屑泥粒岩/颗粒岩; (8) 团块灰岩; (9) 含角砾的泥岩和页岩[5].三种类型的微生物骨架岩在寨江礁的垂直序列上连续产出; 呈肾形状生长的Epiphyton 骨架岩在下, 其上是Renalcis -Palaeomicrocodium 骨架岩, 最上面是发育很好的Epiphyton 骨架岩, 这反映了生物礁发育过程中的环境变化. 礁体沉积中高含量的碳酸盐颗粒和生物屑指示一种浅水的沉积环境. 法门早期的全球海平面下降使得桂林台地暴露, 斜坡沉积随后被潮汐流改造, 伴随一些潮沟的形成. 当海平面再上升并淹没台地时, 微生物迅速占领空置的生境, 开始繁盛, 形成微生物骨架沉积和生物礁[5].2.2 Renalcis 礁和丘在桂林西南约10~11 km 的庙门附近的一座小山出露有法门期的Renalcis 礁(图1), 并在紧邻礁体的礁前斜坡发育有很好的水下岩脉(墙). 礁体厚24 m, 在山体一侧的出露宽度达100 m. 根据区域岩石地层对比, 礁体属法门早、中期的融县组上段. Renalcis 礁可以识别出礁前岩屑相, 礁核相和礁后相. 礁前岩屑相由礁体被海浪打碎的角砾组成, 包括礁骨架岩、球粒泥晶灰岩和球粒微生物泥晶灰岩. 礁体生长在一个由亮晶、砂砾级微生物类和藻类形成的颗粒滩上.礁核相由微生物类(Renalcis, Izhella, Garwoodia, Ri - valluia, Wetheredella, Paraepiphyton, 和Rothpletzella ) 和胶结物形成[15]. 呈生长位置的微生物互相粘结, 形成一些大小不等的微生物粘结块(图4)(一般高30~40 m, 直径5~10 cm). 这些微生物粘结块横向上互相支撑形成18 m 高的刚性骨架. 微生物Renalcis 呈葡萄团粒状或分叉的丛状向上生长(图5(a)). 有些微生物很难鉴定, 暂叫做疑源微生物. 成因与微生物菌和根系有关的礁孔洞常见, 代表礁体露出水面的证据. Renalcis 和Izhella 作为隐生生物首选这些礁穴的壁上生长. 多世代、放射状的纤状方解石胶结物和溶蚀孔洞常见于礁核相. 礁核相的顶部是疑源微生物灰岩、微生物/藻球粒灰岩和钙球灰岩[15], 指示一种短暂的半局限台地环境.第7期沈建伟等: 桂林中、晚泥盆世微生物碳酸盐沉积、礁和丘及层序地层、古环境和古气候的意义631图4 呈生长位置的微生物互相粘结, 形成一些大小不等的微生物粘结块,白色部分是微生物Renalcis的粘结块, 深色部分为碳酸盐泥晶图5 微生物粘结结构和粘结灰岩(a) 微生物Renalcis呈葡萄状团粒形式生长, Renalcis可呈向上分叉的丛体生长; (b) 阳朔岩塘的微生物Renalcis粘结灰岩, 白色呈云朵状的部分和平底上隆成丘形部分是Renalcis, 深色部分是碳酸盐泥晶沉积庙门微生物礁中的水下岩脉分布在180 m宽的礁前相中, 在一个高30 m的陡崖处, 可以清晰的观察和研究水下岩脉. 大多数的水下岩脉沿碳酸盐台地边缘分布, 几乎平行于礁前相. 包壳生物、Renalcis, Izhella和一些Paraepiphyton沿岩脉的壁生长. 它们一般构成起伏的、纹层状的、2~20 cm宽、近乎平行岩墙壁的生长带. 在岩脉中也可以观察到由Renalcis,胶结物和叠层状疑源微生物纹层形成的微生物胶结岩, 及Girvanella和Rothpletzella包壳生物屑和微生物纹层中的窗孔结构. 斑马纹灰岩(0.02~2 m宽)出露于微生物礁体的水下岩脉中, 其内部沉积物由含介形虫和微生物碎屑以及具有泥晶套的钙球组成. 斜三角体的亮晶常见于水下岩脉的斑马纹灰岩和其他充填物中. 放射状的纤状方解石一般是岩脉中的早期胶结物, 它们形成不同宽度的脉体, 在斑马纹灰岩中呈交互的条带. 庙门微生物礁发育的水下岩脉可能与微生物礁的水下生长是同生的.它们形成在一定水深下礁前沉积的断裂和充填阶段, 承受了构造活动的影响.礁后相以具有发育很好的水平层理和窗孔结构632中国科学D辑地球科学第35卷的浅灰色白云质灰岩和钙球灰岩为特征, 只含腹足类和介形虫.礁孔洞在庙门Renalcis礁中非常发育, 包括五种类型: (1) 格架孔洞; (2) 微生物菌和根系孔洞; (3) 裂隙孔洞; (4) 溶蚀孔洞; (5) 开放连通孔洞[15].在阳朔西北靠近白沙的岩塘村(图1)也出露有法门期的Renalcis丘.微生物丘由Renalcis, Renalcis生物胶结岩、亮晶颗粒岩和泥粒岩以及鲕粒灰岩形成, 其中可见次生角砾. 微生物丘发育不连续, 由三个不连续生长、4~15厚的丘和含Renalcis的灰岩叠置形成丘和浅滩沉积相间的沉积序列, 总的地层厚度超过100 m.下部的丘5 m厚, 直径约10 m. 中部的丘最大, 8 m厚, 直径约40 m, 上部的丘厚度4 m左右, 直径达50 m.微生物丘主要由Renalcis和胶结物形成(图5(b)). Renalcis的形状多样, 有房室状、气囊状和团状.Paraepiphyton常见, 主要生长在丘的核部, 密集叠生, 形成树枝状和丛状的骨架.Paraepiphyton常与微生物鲕粒、内碎屑、团块和团粒产出在一起. 在丘的翼部发育斑马状平底晶洞构造.2.3 叠层石礁桂林晚泥盆世法门期微生物形成的叠层石礁主要出露在沙塘和庙门(图1), 一般产出在融县组上段下部的球粒和鲕粒灰岩段. 在古地理位置上, 它们沿台地边缘分布. 沙塘叠层石礁是其中最大的一个, 达76 m厚, 250 m长, 位于桂林西北约5 km[16] (图1). 根据牙形刺生物地层和岩石地层对比, 礁的年代归于法门早期. 礁生长在一个生物碎屑和微生物/藻滩上, 滩是由砂、砾级的、方解石亮晶胶结的红藻、管孔藻, 微生物、腹足类、双壳类和腕足类的生物屑构成, 生物屑占岩石量的70%以上, 多世代放射状纤状方解石胶结物在滩相岩石中发育很好. 水下岩脉发育, 但不常见. 叠层石礁的核部由灰色含柱状和分枝状的叠层石灰岩形成(图6), 厚度一般超过39 m. 两种比较特征的叠层石构成礁灰岩, 一种是细圆柱状的, 4.5~20 cm高, 直径1~2.5 cm, 大的直径可达3.5~5 cm. 叠层石的横断面呈圆形、不规则或椭圆形. 叠层石的外壁呈锯齿状, 叠层石之间充填有微生物Garwoodia, 管孔藻、拟刺毛藻、海百合茎、有孔虫、腕足类、介形类、球粒和核形石的碎屑. 另一种是树枝状的叠层石, 它们的特征与柱状的相似, 不同点在于前者分开成两个平行的变小的叠层石. 当细小而分叉的叠层石近于平行或垂直主叠层石时, 叠层石就呈分枝状或蘑菇状. 在礁灰岩的风化面上, 可观察到灰白色的柱状和蘑菇状的叠层石, 含量占岩石的50%~60%. 叠层石高度一般30~65 mm, 有些达到20图6 桂林沙塘微生物形成的叠层石礁和丘, 主要由柱状和分叉的叠层石构成 (Shen 和 Webb, 2004)第7期沈建伟等: 桂林中、晚泥盆世微生物碳酸盐沉积、礁和丘及层序地层、古环境和古气候的意义633cm. 叠层石彼此连接形成刚性骨架. 在形成叠层石的富微生物层和富沉积物层之间的孔洞可见示底结构, 指示一种浅水沉积环境[16].沙塘叠层石礁的上覆岩层是浅灰色块状的生物屑颗粒灰岩和透镜状的鲕粒颗粒灰岩互层, 之上是灰白色厚层状的球粒灰岩、疑源微生物灰岩和亮晶砂、砾级的内碎屑灰岩. 水平纹层和窗孔结构常见于上述灰岩中. 礁孔洞类型包括: (1) 微生物粘结岩中的孔洞; (2)拟刺毛藻骨架中的孔洞; 和(3) 叠层石纹层中的孔洞[16].3 微生物碳酸盐沉积和层序地层桂林泥盆系碳酸盐台地及礁组合沉积的中、晚泥盆世地层中5个三级层序界线的时代可初步确定为(1) 下varcus亚带和中varcus亚带之间; (2) hermanni- cristatus和disparilis带之间; (3) 下rhenana亚带和上rhenana亚带之间; (4) expansa带内部; (5) praesulcata 带和sulcata带之间[7](图7). 比较明显的大的海泛面有两次: (1)ensensis带和varcus带之间; (2) linguiformis 带和下triangulasis亚带之间[7](图7). 第一个层序由艾非尔晚期海侵体系域的浅水陆架碎屑沉积和吉维早期高水位体系域的碳酸盐浅滩沉积组成(图7). 这种在碎屑陆架上广布的浅滩环境中由大量的珊瑚、层孔虫和其他后生生物主导, 微生物碳酸盐沉积不发育. 第二个层序由吉维中期海侵体系域中至厚层状的台地相碳酸盐沉积和吉维晚期高水位体系域图7 桂林中、晚泥盆世层序地层、海平面变化和微生物礁和丘634中国科学D辑地球科学第35卷厚层状、向上变浅的碳酸盐沉积组成(图7). 海侵体系域代表的是浅滩向碳酸盐台地的转变, 同时基底断裂开始活动, 浅滩被断裂切割, 断块的差异活动为后来台地和台沟的发育创造了条件. 这一时期的微生物沉积不发育. 高水位体系域代表的是碳酸盐台地发育时期, 这时台地沉积相分异明显, 台地、台地边缘和台缘斜坡相发育完好. 微生物(Rothpletzella)沉积在台缘斜坡环境形成叠层石生物层[17], 在台地内部环境形成纹层状灰岩. 在高水位体系域沉积顶部靠近层序界面, 微生物(Rothpletzella)形成很小的叠层石, 一般高1~2 cm, 密集排列, 形成似纹层的灰岩. 第三个层序由弗拉早期海侵体系域的中层状粒泥灰岩、微生物纹层灰岩和弗拉晚期高水位体系域富含双孔层孔虫、球状生长的层孔虫、珊瑚和腕足类的灰岩构成(图7). 在海侵体系域中, 微生物碳酸盐沉积不仅在台内环境形成纹层灰岩和小的核形石, 而且在台地边缘环境形成纹层灰岩(如: 凉水井剖面[17])和叠层石生物层[7]. 在高水位体系域中, 微生物碳酸盐沉积主要发育在台地边缘和台缘斜坡环境, 形成核形石(如: 凉水井剖面[17])以及戴帽核形石和凝块状核形石[7]. 第四个层序由弗拉最晚期海侵体系域的向上变薄的灰岩和法门早、中期高水位体系域的窗孔灰岩和微生物纹层灰岩构成(图7). 在海侵体系域中, 微生物在台缘环境形成核形石, 并在海侵体系域的最晚期开始形成微生物礁和丘. 在高水位体系域中, 微生物碳酸盐沉积广泛发育, 除了在台内环境形成窗孔灰岩和微生物纹层灰岩以外, 在台缘和近台缘的斜坡环境形成各种微生物礁、丘和叠层石礁[5,15,16]. 第五个层序由法门最晚期海侵体系域的微晶生物碎屑灰岩和高水位体系域向上变浅的微晶灰岩、薄层状砂屑灰岩、少量窗孔灰岩和含珊瑚和核形石的灰岩构成(图7). 微生物沉积在海侵体系域不发育, 在高水位体系域主要形成核形石.4 讨论桂林晚泥盆世(法门期)灰岩记录了微生物礁组合的发育和衰亡, 它们一般产出在法门早、中期.因为微生物和微生物石反映古地理和沉积特征, 研究它们可洞察正常海相台地边缘和礁前斜坡环境中生长的微生物礁、泥盆系生物礁的发育及生物事件, 因此, 它们可用来研究环境变化、生物事件、层序地层和海平面变化.在层序地层不同体系域中有不同的微生物形成的碳酸盐沉积(表1): 高水位体系域的台地海退期, 在正常海洋沉积环境中, 微生物群落转移到有压力的环境或隐生的礁生境. 在隐生的礁缘环境中形成无骨骼的微生物石和由微生物产生的胶结物. 如桂林弗拉期, 微生物一般殖生在层孔虫和珊瑚骨架孔洞的隐生环境中. 在海侵体系域的礁后环境中, 微生物碳酸盐形成小的核形石, 在潮间环境形成叠层石. Renalcis在高水位礁缘相起粘结和胶结的作用. 在高水位体系域, 微生物碳酸盐沉积往往形成大的戴帽核形石和凝块核形石. 在高水位的底部, 直接生长在碎屑沉积之上的叠层石一般代表初期到最大的台地淹没事件(如凉水井剖面[17]). 在低水位体系域, 微生物碳酸盐在台内和潮缘向上变浅的旋回中形成叠层席(如杨堤剖面[7])和微生物纹层(如桂林炼油厂背后剖面[7]), 这些微生物沉积往往与层序界面有关. 台地的海泛代表的是生物造礁作用的暂时停止, 而在法门最底部的台地海泛与弗拉期造礁生物的绝灭有表1 微生物碳酸盐沉积在桂林中、晚泥盆世层序地层和台地不同环境中的产出海侵体系域(TST) 高水位体系域(HST) 台内台缘台缘斜坡台内台缘台缘斜坡纹层状灰岩层序Ⅲ层序Ⅲ层序Ⅱ层序Ⅳ核形石层序Ⅲ层序Ⅳ层序Ⅲ层序Ⅲ层序Ⅴ凝块核形石层序Ⅲ叠层石层序Ⅲ层序Ⅳ层序Ⅱ礁和丘层序Ⅳ层序Ⅳ层序Ⅳ第7期沈建伟等: 桂林中、晚泥盆世微生物碳酸盐沉积、礁和丘及层序地层、古环境和古气候的意义635联系. 桂林法门期叠层石礁形成在法门期高水位体系域的底部, 成礁时间早于Renalcis-Epiphyton礁和Renalcis礁, 因此, 其与层序界面有关(图7). 这可以与巴哈马现代潮间带的凝块叠层石和西澳鲨鱼湾的潮间带叠层石进行对比, 前者指示一种海平面上升和营养物质增加的环境[18]; 后者是海平面相对低时形成的[19].一般认为晚泥盆世生物礁的衰落经历了一段很长的时间(从吉维晚期到法门晚期)[20], 钙质微生物、无骨骼的微生物石和生物胶结岩随时间逐步增加并最终成为主要造礁组分. 珊瑚和层孔虫礁可指示一种寡养分的(oligotrophic)生态系, 这时微生物碳酸盐沉积转移到隐生和有压力的海洋生境; 微生物群落和形成的微生物石指示一种中养分(mesotrophic)的生态条件. 一般认为微生物很可能在营养物质供应发生变化时获得繁盛. 叠层石、核形石和大的微生物凝块叠层石一般直接占领后生生物绝灭后的生境[4]; 但它们作为一种灾难劫后生物还有争议[16]. 当生物绝灭事件后, 后生生物大量减少, 相对富养成分(eutrophic)增加时, 微生物群落将获得大发展, 如现代弗罗里达礁岛的微生物碳酸盐生产率的增加是由营养增加和养分富集引起[21]; 这也被用来解释生物绝灭事件[22]. 标志中、晚泥盆世阶的界线和生物礁类型的主要生物事件在桂林地区表现很明显. 吉维期末的生物危机造成艾菲尔和吉维期的主要造礁生物(块状层孔虫和皱纹珊瑚)大部分绝灭, 吉维期的珊瑚-层孔虫礁群落被弗拉期的层孔虫-珊瑚-钙质微生物群落代替. 弗拉期生态表型为球状、分枝状和不规则状的层孔虫类型取代了吉维期的板状、块状和圆丘状类型的层孔虫, 环境由吉维期寡养成分变为弗拉期的中养成分, 钙质微生物类和藻类从吉维期的礁前斜坡环境迁移到弗拉期的台地边缘环境, 成为重要的造礁生物. 晚泥盆世弗拉-法门界限和法门-杜内界限附近的生物事件终结了后生生物骨骼造架进入法门期和杜内期[23]. 受这些生物事件的影响, 晚泥盆世礁骨架的组分从后生生物骨骼变为钙质微生物、无骨骼的微生物石和生物胶结岩, 环境又由弗拉期的中养成分变为法门期的富养成分. 因此, 桂林晚泥盆世法门期完全由微生物形成的生物礁指示一种后生生物绝灭后从寡养分到富养分的环境变化.微生物礁和碳酸盐泥丘从元古代到今天都有发育, 全球古地理重建表明其分布广泛, 从热带到温带和极地都有产出. 对已报道和研究的地质时期微生物礁和碳酸盐泥丘的统计表明, 冰期阶段是微生物礁和碳酸盐泥丘最为发育的时期. 这一时期凉或冷的海水侵占了海底. 这种冷海水在深水区更为普遍, 随着上升流和海平面上升, 它们淹没过陆架1), 使碳酸盐沉积物形成特殊的沉积构造. 不管是地质时期还是现代的微生物礁和碳酸盐泥丘都包含有发育很好的平底晶洞和斑马构造, 这些构造代表早期的孔洞, 并由早期海底胶结物固定. 如果这些孔洞的形状多样时, 它们就有可能是先前天然气水合物沉积(充满天然气的冰)的残余物. 具有重要意义的是, 现今的海洋天然气水合物的产出往往与凉或冷的海水和丰富的碳酸盐亮晶和泥晶胶结物有关, 泥丘的早期海底胶结作用形成稳定的沉积基底, 光合营养的、非自养的和厌养的微生物殖生其上.晚泥盆世法门早、中期特别丰富的微生物类和藻类的碳酸盐沉积可以用作解释法门期冰室和海平面下降的一个原因. 最近的研究[24,25]证实海洋中单细胞生物(微生物类和藻类)都会分泌一种化学物质——二甲硫化物(dimethyl shlfide (DMS))来保护自己以避免高盐度的伤害. 当微生物类和藻类死亡腐烂后, 二甲硫化物就会进入海水中, 并最终随海水一起蒸发而进入大气层. 二甲硫化物进入大气层后就会氧化分解出硫. 这些硫的颗粒最易吸聚水汽而成为水滴的核心. 随着水滴在大气中的不断增加, 大量的水滴云就会形成. 这些水滴云反射阳光而使地球表面的温度下降, 冰室气候形成. 当气温下降时, 微生物、藻类的生长和繁盛就会减弱, 气候又重新回暖. 微生物类、藻类繁盛的同时, 会消耗掉海水中的大量氧气, 其他生物会大量减少.1) Krause F F. Paleozoic carbonate mud-mound evolution and global paleogeography. Abstract, 2002 Denver annual meeting of the Geological Society of American。
从海岸到盆地沉积表面坡度极缓(小于1°)大陆架缓坡浅水环境内形成的一套有成因联系的碳酸盐沉积相的组合。
20世纪六七十年代在对现代碳酸盐海岸大陆架的研究中建立了碳酸盐台地沉积模式(Wilson等,1975),即台地边缘有生物礁形成的障壁岛,或有碳酸盐浅滩镶边,台地内有广阔且平坦的浅水碳酸盐、低能潟湖和碳酸盐潮滩,通常所指的有碳酸盐沉积的浅海环境。
镶边的礁或浅滩向海盆方向有一陡坡通向深水盆地,从台地上崩落的浅水碳酸盐在陡坡外可发生再沉积。
鉴于很难用碳酸盐台地模式概括古代陆表海碳酸盐沉积的特征,Ahr (1973)又提出碳酸盐缓坡的术语,用来描述从滨海波浪作用的浅水逐渐过渡到低能的深水环境,沉积物表面坡度极缓(一般小于1°),不存在明显坡折的碳酸岩沉积体系; 而将上述有生物礁或浅滩镶边、前缘有陡坡的碳酸盐台地称为碳酸盐陆架或镶边陆架沉积体系。
至于碳酸盐台地这一术语,现已演变为统指所有浅水碳酸盐沉积,包括缓坡、镶边陆架和孤立的岩隆,特别是某些一时尚难确定其沉积环境的碳酸盐沉积的非正式术语。
碳酸盐缓坡的沉积特征对比古生代和中生代世界缓坡碳酸盐沉积体系与生物礁和浅滩镶边的碳酸盐陆架沉积体系,两者的区别是,①缓坡碳酸盐沉积物表面坡度极小; 而镶边陆架碳酸盐沉积物表面则存在明显坡折。
②缓坡不存在生物礁。
③缓坡近岸内坡相碳酸盐产率比镶边陆架台地顶部少。
④海平面略有下降时,缓坡相带向海盆方向迁移,不会同时完全暴露,而镶边陆架的台地表面会全部暴露。
⑤当海泛缓慢发生时,缓坡不会象镶边陆架的台地那样突然被海水淹没,而是被缓慢逐渐淹没,有利于潜水面的不断上升,使泥炭沼泽环境得以长期持续,形成厚煤层甚至巨厚煤层;而易被海水突然淹没的镶边陆架上则很难形成厚煤层。
碳酸盐缓坡的沉积相碳酸盐缓坡上的沉积作用随水深、水温和波浪、潮汐作用强度的变化而异,主要的沉积相有内坡相、中坡相、外坡相和盆地相等(下图)。
碳酸盐缓坡的主要沉积相A—内坡相潟湖潮滩沉积; B—内坡相潟湖沉积; C—内坡相高能浅滩或障壁岛沉积;D—中坡相; E—外坡相; F—盆地相(据Burchette和Wright,1992)内坡相上临滨(海滩或潟湖滨线)和正常浪基面之间的缓坡沉积作用带。