型
该模型认为膜的骨架是脂肪形成的脂双层结 构,脂双层的内外两侧都是由一层蛋白质包 被,即蛋白质-脂-蛋白质的三层结构,内外两 层的蛋白质层都非常薄。并且,蛋白层是以 非折叠、完全伸展的肽链形式包在脂双层的 内外两侧。1954年对该模型进行了修改:膜 上有一些二维伸展的孔,孔的表面也是由蛋 白质包被的,这样使孔具有极性,可提高水对 膜的通透性。这一模型是第一次用分子术语 描述的结构
膜糖的存在方式
通过共价键同膜脂或膜蛋白相连,即以糖脂或糖蛋 白的形式存在于细胞质膜上。
糖同氨基酸的连接主要有两种形式,即O-连接和N-连接
O-连接:是糖链与肽链中的丝氨酸或苏氨酸残基相连, O-连接糖链较短, 约含4个糖基。
N-连接: 是糖链与肽链中天冬酰胺残基相连,N-连接 的糖链一般有10个以上的糖基。另外,N连接的方式较O 连接普遍。
膜脂的不对称性
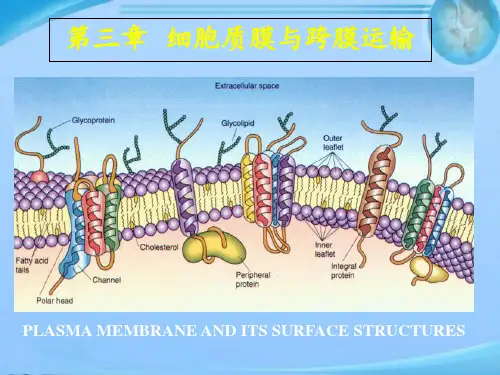
细胞质膜各部分的名称 膜脂与糖脂的不对称性
糖脂仅存在于质膜的ES面,是完成其生理功能的结构基础 非对称性形成原因: 磷脂:ER胞质半膜合成,Flippase选择性转运 糖脂: 催化糖基化反应的酶位于Golgi非胞质半膜,转运不
变
膜糖
存在于原核和真核细胞的质膜上(5%以下),神经细胞糖 脂含量较高;细胞质膜上所有的膜糖都位于质膜的外表面,
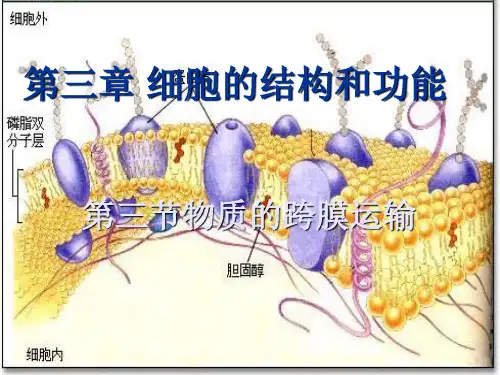
极性的头部、非极性的类固醇环结构和一个非极性的碳氢尾部。胆固醇的分子较 其他膜脂要小, 双亲媒性也较低。胆固醇的亲水头部朝向膜的外侧,疏水的尾部埋 在脂双层的中央。胆固醇分子是扁平和环状的,对磷脂的脂肪酸尾部的运动具有 干扰作用,所以胆固醇对调节膜的流动性、加强膜的稳定性有重要作用。
胆固醇的分子较其他膜脂要小, 双亲媒性也较低。胆固醇的亲水头部朝向 膜的外侧,疏水的尾部埋在脂双层的中央