植物病原真菌对内吸性杀菌剂的抗药性研究进展
- 格式:doc
- 大小:48.00 KB
- 文档页数:11


药用植物内生菌促生与生防作用研究进展作者:徐源清朱越韩佳妮徐桂雯黄轶勉周芳美来源:《农业灾害研究》2024年第02期摘要:我国幅员辽阔且气候多样,药用植物资源丰富的同时,蕴含着多样的内生菌资源。
药用植物内生菌和其宿主植物作为一个共生体,在长期协同进化的过程中,已经形成互惠共存的关系。
在药用植物生长过程中,内生菌既能通过促进植物对氮、磷、钾、铁等无机营养元素的吸收利用和生长激素的合成直接促进药用植物的生长,也能通过生物防治功能间接促进药用植物的生长。
综述了近年来药用植物内生菌促生与生防作用的研究及应用,并对药用植物内生菌的发展趋势进行了展望。
关键词:药用植物;内生菌;促生作用;生物防治中图分类号:S567 文献标志码:B 文章编号:2095–3305(2024)02–00-03植物内生菌是指其生活史的部分或全部阶段生活于健康植物表皮细胞层间隙或组织器官内部的一类微生物,不易受环境条件的影响,可以在宿主植物体内独立地分裂繁殖和传递,在与宿主植物长期协同进化过程中逐渐形成互惠共存的关系。
研究表明,从草本植物、木本植物到藤本植物,这些植物体的根、茎、叶、花、果实等组织器官都普遍存在内生菌,而我国药用植物资源极其丰富,内生菌资源更为多样[1]。
此外,药用植物内生菌的价值极高,在医药、农业等领域可以发挥重要作用。
综述了药用植物内生菌如何发挥促生作用,帮助宿主植物获取生长所需的资源和调节宿主植物生长发育及产生生物防治的效果,为解决某些药用植物繁殖率低、生长困难,致使资源日益枯竭和缓解其他珍稀药用植物生长困境及其他常见农作物供应不足等问题提供理论指导。
1 药用植物内生菌的种类随着对植物内生菌的深入研究,人们对内生菌资源的多样性及价值性认知不断提高,开发和利用不断广泛,这为促植物生长和植物疾病生物防治工作提供了新思路。
近年来,药用植物内生菌也越来越受到国内外研究者的关注。
研究表明,药用植物内生菌种类繁多,主要包括内生细菌、真菌和放线菌,在各种药用植物中分布广泛[2]。

常见杀菌剂作用机理、防治对象及延缓抗药性方法一、杀菌剂类别凡是对病原物有杀死作用或抑制生长作用,但又不妨碍植物正常生长的药剂,统称为杀菌剂。
随着杀菌剂的发展,又区分出杀细菌剂、杀病毒剂、杀真菌剂等亚类。
(一)按使用方式分类1、保护剂保护剂在病原微生物没有接触植物或没浸入植物体之前,用药剂处理植物或周围环境,达到抑制病原孢子萌发或杀死萌发的病原孢子,以保护植物免受其害,另当病原菌落在植物体上接触到药剂而被毒杀,这种作用方式称为保护作用。
具有此种作用的药剂为保护剂。
如波尔多液、代森锌、硫酸铜、绿乳铜、代森锰锌、百菌清等。
2、治疗剂病原微生物已经侵入植物体内,但植物表现病症处于潜伏期。
药物从植物表皮渗入组织内部,经输导、扩散、或产生代谢物来杀死或抑制病原,使病株不再受害,并恢复健康。
具有这种治疗作用的药剂称为治疗剂。
如甲基托布津、多菌灵、春雷霉素等。
3、铲除剂指植物感病后施药能直接杀死已侵入植物的病原物。
具有这种铲除作用的药剂为铲除剂。
如福美砷、五氯酚钠、石硫合剂等。
内吸性杀菌剂内吸性杀菌剂能被植物叶、茎、根、种子吸收进入植物体内,经植物体液输导、扩散、存留或产生代谢物,可防治一些深入到植物体内或种子胚乳内病害,以保护作物不受病原物的侵染或对已感病的植物进行治疗,因此具有治疗和保护作用。
如多菌灵、力克菌、多霉清、霜疫清、噻菌铜、甲霜灵、乙磷铝、甲基托布津、敌克松、粉锈宁、甲霜铜、杀毒矾、拌种双等。
(二)按传导特性分类1、内吸性杀菌剂二、延缓抗药性的方法1、防止病菌抗药性发展。
不能连续用一种杀菌剂,也不能一个生长期连续数次用同一类杀菌剂,对内吸剂应限制使用次数。
2、要根据传病规律,采取相应对策。
农作物病害有空气传播病原、种子苗秧带菌传播土壤或土杂肥带菌传播有害昆虫和螨类传播等几大类。
如蚜虫、飞虱等传播病毒造成的病害,则应在带毒害虫危害作物前治虫。
若带毒昆虫已吸食寄主汁液,再治虫已不能防病。
种传达室病害、如许多禾本科作物的黑穗病等需在种子处理方面下功夫,秧苗带菌则应先处理秧苗后下田。

耋壁堡望:竺:!!!!呈!竺!!!!!竺竺竺竺!!!竺竺!竺墨竺兰墨_・黄瓜白粉病研究进展王友平朱金英郭平银高凤菊摘要论述了黄瓜白粉病病原茵、症状及其发生侵染规律,并对抗性遗传、生理机制、诱导抗性、植物源农药及抗白粉病的其他方法等方面的研究进展进行了综述。
关键词黄瓜白粉病抗病性诱导抗性生物防治ProgressofPowderyMildewinCucumberWANGYouping,ZHUJinying,GUOPingyin,GAOFengjuAbstract:Thefollowingtopicsarereviewedsuchaspathogenicbacteria,symptom,invadingrule,inheritanceofresis-iarlce,physiologicalmechanism,inducedresistance,plant--originpesticide,andcontrolmethodsincucuInber.Keywords:Cucumber;Powderymildew;Resistance;Indueedresistance;Biologicalcontrol白粉病俗称白毛,是一种广泛发生的世界性病害。
在我国,春保护地和夏秋露地黄瓜每年都因白粉病的发生造成大量损失llI,病原菌主要通过气流传播,具有潜育期短、再侵染频繁、流行性强等特点,对黄瓜生产为害严重,导致严重的经济损失[21。
目前,主要防治方法是使用抗病品种及化学农药,但因这2种方法都存在各自的局限性,迫使人们不断地探索新的防治方法。
本文就黄瓜白粉病病原菌、症状、发生侵染规律,抗性遗传、生理机制、诱导抗性、植物源农药及抗白粉病的其他方法等方面进行了综述,以期为白粉病抗性研究提供参考。
1白粉病的病原菌及症状1.1病原菌黄瓜白粉病的病原菌记载较为混乱,《中国真菌志》记载为瓜类单囊壳(Spbaerothecacucurbitae)和葫芦科白粉菌(Erysiphecucurbitacearum)13],与《中国蔬菜病虫原色图谱》闱一致;也有研究认为是E.cicboraceamra和S.fuligenea,并认为后者更为常见阁,屈振淙t61亦研究认为我国吉林省黄瓜上发生的白粉病的优势菌为S.fuligenea。

植物内生菌的杀菌效果研究植物内生菌指的是一类共生于植物体内的微生物,它们与宿主植物形成互惠共生关系,帮助植物吸收养分、促进生长并增强抗病能力。
近年来,研究人员发现一些植物内生菌还具有杀菌效果,能够抑制植物病原菌的生长和繁殖。
本文将对植物内生菌的杀菌效果进行研究。
植物病原菌是导致植物疾病的主要原因,它们会侵入植物体内并繁殖,最终导致植物死亡或减产。
传统的防治方法主要依赖化学农药,但其存在对环境和人体健康的副作用。
因此,寻找一种安全、有效的杀菌方法显得尤为重要。
植物内生菌的杀菌效果引起了研究人员的兴趣。
一方面,植物内生菌可以产生一系列抗菌物质,如抗生素、植物生长调节物质等,对植物病原菌产生抑制作用。
研究人员通过实验发现,一些植物内生菌能够分泌抗菌物质,如溶菌酶、蛋白酶和生物活性物质等,这些物质能抑制病原菌的生长、限制其繁殖,并最终导致其死亡。
此外,抗菌物质还可以激活植物的防御机制,增强植物的抗菌能力,减轻病害的发生。
另一方面,植物内生菌还能调节植物内部环境,限制病原菌的生长。
研究人员发现,一些植物内生菌能够调节植物的根际环境,产生一股抑菌区域,抑制病原菌的入侵。
这种抑菌机制与植物内生菌的共生生活方式密切相关,植物通过合成一些特定的物质,如抗菌物质和植物生长激素,形成对病原菌的抑制作用,从而保护自己免受病害侵害。
在实践中,研究人员对植物内生菌的杀菌效果进行了一系列研究。
以溶菌酶为例,研究人员通过实验发现,溶菌酶能杀灭多种病原菌,如炭疽菌、普通立枯病菌、松材线虫等。
另外,研究人员还发现,植物内生菌分泌的蛋白酶可以降解病原菌的细胞壁,导致其破裂死亡。
这些研究结果表明,植物内生菌具有潜在的杀菌效果,并有望成为一种新型的绿色杀菌剂。
尽管植物内生菌具有较好的杀菌效果,但目前仍存在一些问题。
首先,植物内生菌的杀菌效果受到多种因素的影响,如菌株的选择、培养条件、环境因素等。
因此,如何提高植物内生菌的杀菌效果仍然是一个挑战。

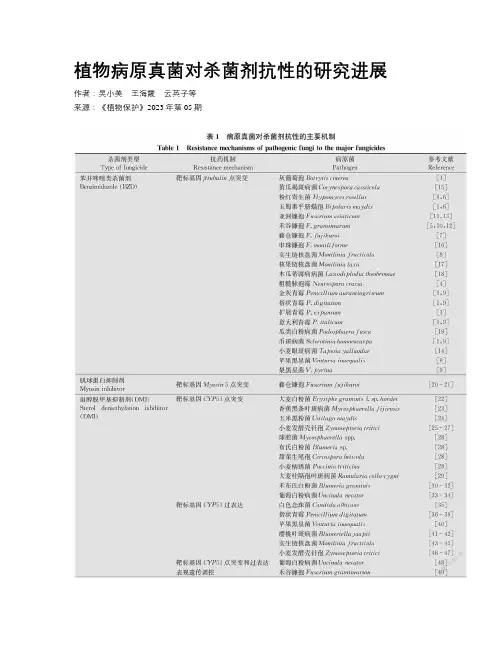
植物病原真菌对杀菌剂抗性的研究进展作者:吴小美王海霞云英子等来源:《植物保护》2023年第05期关键词植物病原真菌;杀菌剂抗性机制;抗性检测;抗性治理对策中图分类号:S 481.4 文献标识码:A DOI: 10.16688/j.zwbh.2023057杀菌剂在植物病害防治中发挥着重要作用,然而随着药剂的长期使用,病原菌抗药性问题也日趋严重,成为制约药剂防治效果和使用寿命的重要因素之一。
20世纪70年代之前所使用的杀菌剂几乎都是保护性杀菌剂,作用位点多,不易引发病原菌产生抗药性,但是随着杀菌剂的发展进入高效、内吸、作用位点较为单一的内吸型杀菌剂时代以来,杀菌剂抗性问题愈发普遍和严重,已成为化学防治所面临的一大挑战。
由于病原菌对杀菌剂的抗性可以随着病原菌的繁殖而稳定遗传给后代,病原菌的抗性群体在药剂的选择压力下会逐步扩展,进而使得病菌群体对杀菌剂的敏感性整体下降。
病原菌的抗药性主要是由病原菌的单个或者多个基因突变造成的,在自然条件下抗性菌株在病菌群体中出现的频率很低,因此不会影响杀菌剂对病害的防治效果。
但是,由于杀菌剂的连续使用,在持续的药剂选择压力下,敏感菌株生长繁殖受到抑制,抗药菌株得以迅速生长和繁殖,在病菌群体中逐渐占优势地位,从而导致杀菌剂的防效下降甚至失效。
抗性菌株的适合度(包括温度适应性、产孢和致病能力等)决定了抗性菌株群体的发展趋势。
如果抗性菌株适合度低,在自然环境中生存力弱,那么一旦停止使用杀菌剂,抗性菌株的种群比例就会下降;但是如果抗性菌株与敏感菌株的适合度相似,在自然界能够保持良好的生长繁殖和致病能力,则容易导致田间杀菌剂抗性问题。
总体来说,病原菌对杀菌剂产生抗性的机制主要包括以下几种情况:1)杀菌剂作用靶点突变导致药剂与靶标的结合能力降低;2)杀菌剂靶标基因的过量表达;3)病菌对杀菌剂外排能力或代谢分解能力增强。
除此之外,近来研究发现,表观遗传在病菌抗药性中也发挥重要作用。
本文对几类常用杀菌剂的抗药性现状及抗性机制进行综述,包括:苯并咪唑类杀菌剂(benzimid-azole,BZD)、肌球蛋白抑制剂(myosin inhibitor)、甾醇脱甲基抑制剂(sterol demethylation inhibitor,DMI)、QoI类抑制剂(quinone outside inhibitor,QoI)、琥珀酸脱氢酶抑制剂(succinate dehydrogen-ase inhibitor,SDHI)以及二甲酰亚胺类杀菌剂(di-carboximide,DC)。

小麦赤霉病菌对多菌灵和不同杀菌剂敏感性的相关分析-农学论文小麦赤霉病菌对多菌灵和不同杀菌剂敏感性的相关分析陈宏州1,肖婷1,许媛1,狄华涛1,马圣洲1,范婷2,杨敬辉1 (1江苏丘陵地区镇江农业科学研究所,江苏句容212400 ;2江苏农林职业技术学院,江苏句容212400)摘要:为探明江苏省小麦赤霉病菌[Gibberella zeae (Schwein.) Petch]对多菌灵的抗药性和该药剂与其他杀菌剂的交互抗性,采用区分剂量法检测了采自江苏省26个县(市)的520株小麦赤霉病菌对多菌灵的抗药性,并采用菌丝生长速率法分别检测了对多菌灵不同敏感性的10个菌株对嘧菌酯、吡唑醚菌酯、肟菌酯、唑胺菌酯、氟环唑、己唑醇、灭菌唑和咯菌腈等杀菌剂的敏感性。
结果表明:江苏省各县(市)菌株对多菌灵的抗性频率差异较大,总抗性频率为50.58% ;通过EC50值相关性分析,小麦赤霉病菌对多菌灵和上述杀菌剂之间不存在交互抗性。
江苏省小麦赤霉病菌对多菌灵的抗性频率较高,迫切需要筛选新的杀菌剂防治小麦赤霉病。
关键词:小麦赤霉病菌;多菌灵;抗药性;交互抗性中图分类号:S435.121文献标志码:A论文编号:cjas基金项目:江苏省农业科技自主创新资金项目“稻麦两熟制高产平衡技术方案” [CX(15)1002] ;镇江市农业科技支撑项目“小麦赤霉病抗性检测与防控技术研究” (NY2013003);“市域主要农作物病原菌抗药性检测与监测” (NY2015019) 。
第一作者简介:陈宏州,男,1984年出生,广西宜州人,助理研究员,硕士,主要从事农作物病害抗药性检测与治理研究。
通信地址:212400江苏省句容市弘景路1号江苏丘陵地区镇江农业科学研究所,Tel: 9,E-mail : [emailprotected]。
通讯作者:杨敬辉,男,1973年出生,云南丽江人,研究员,博士,主要从事植物保护研究。
通信地址:212400江苏省句容市弘景路1号江苏丘陵地区镇江农业科学研究所,Tel: 9,E-mail : [emailprotected]。

QoI类杀菌剂——一类成功的农用杀菌剂的兴衰本文来源:The QoI Fungicides, the Rise and Fall of a Successful Class of Agricultural Fungicides 翻译:薛鹤、张新新1、简介真菌性病害对作物是一个重大威胁。
为了应付这个问题,种植者使用了巨额的化学品,杀真菌剂早已被用于减少作物损失。
植物病害的药剂防治开始于19世纪中后期,当时杀菌剂的主要活性成分为硫,基于此石灰-硫酸铜混合液(波尔多液)被使用。
直到20世纪40年代时,可用杀菌剂阿森纳还是非常少的。
植物病害的防控主要还是依靠这类由用户制备的无机化学制剂。
第二次世界大战对大多数国家的化学产业来说是一个有重大进展的时期。
根据战前化学科学,战后出现新的化学学科和新的化合物。
但是几乎所有成分都是保护剂,也就是说,他们需要在该真菌还未侵染作物时就要存在于作物体内,因为作物一旦感染,他们是没有效果的。
一种抗生素的引进对化学防治药剂发展是一个显著的进步。
1955年,在日本研发出一种专门针对稻瘟病防治的抗生素,灭瘟S.,这是第一个系统性的产品,可以在一个领域被广泛应用。
20世纪60年代,作物化学保护取得相当大的发展。
前期出现了一些前几年产品的整合,并从既定化学领域做进一步的研发。
成功合成化合物如代森锰锌(二硫代氨基甲酸酯)和敌菌丹(邻苯二甲酰亚胺)。
然而,这十年的主要成就是引进了内吸杀菌剂噻菌灵和苯菌灵,其共同特征是甲基苯并咪唑氨基甲酸酯作用(MBC)模式,这标志着国内植物治疗药剂的问世。
此外,百菌清(一个邻苯二甲腈),萎锈灵和氧化萎锈灵(羧酸),乙嘧酚(2-氨基吡啶)和吗啉杀真菌剂的第一批成员,十二环吗啉和环吗啉,此时也都出现了。
然而,20世纪70年代可能是作物保护化学进步最显著的十年。
此时的农用化学品市场增长速度实质约为每年6.3%。
乙磷铝,膦酸盐类杀真菌剂,有在韧皮部移动的独特性,是上世纪70年代的重大发现之一。

农药单剂在使用时往往受防治效果、使用范围和药害等因素制约,而且随着病虫害抗药性的增强,防治效果降低。
目前,农药科研人员在研发新药的同时,主要通过复配来克服原有农药单剂的使用缺陷。
然而,复配农药出现时间不长,尚没有统一的复配原则和标准,不利于复配农药的发展。
本文主要针对近几年国内农药专家的研发经验总结整理而成。
1 农药复配剂的类型农药混合剂按其作用对象主要分为以下几种。
1.1 杀虫剂混合剂为了克服单一杀虫剂存在的不足,将不同类型、不同作用方式的杀虫剂复配。
目前主要有有机磷类与有机磷类、有机磷与拟菊酯类、有机磷与氨基甲酸酯类、有机氮与氨基甲酸酯类以及有机氮与拟菊酯类等复配方式。
这是混剂中品种最多的一类,截止到1998年底,在农业药检所注册登记的杀虫剂为4230个,其中杀虫混剂为1684个,占39.8% [1]。
1.2 杀虫杀菌剂混合剂此类型主要用于拌种或土壤处理,发挥杀虫、杀菌兼治作用。
如10%甲柳·酮乳油、35%马·酮乳油和40%氧乐·酮乳油等杀虫杀菌混剂等[2]。
1.3 杀菌剂混合剂为延缓植物病原菌对内吸杀菌剂的抗性,常将内吸性杀菌剂与保护性杀菌剂复配使用。
内吸性杀菌剂能被植物吸收,起到杀菌效果;保护性杀菌剂残留在植物体表,防止病菌入侵感染。
如15%双·多悬浮剂[3]和40%三唑酮·多可湿性粉剂[4]等。
1.4 除草剂混合剂将持效期长、短不同的除草剂搭配;将内吸传导性除草剂与触杀性除草剂搭配;根据杀草谱互补原理,杀单子叶杂草的除草剂与杀双子叶杂草的除草剂混用。
如5.3%丁·西颗粒剂[5]和48%乙·莠可湿性粉剂[6]等。
1.5 杀螨剂混合剂由于单一杀螨剂往往对螨的发育状况具有较大选择性,有效控制期差异较大,为了达到对各个发育阶段都有较好的防治效果,有效控制期适当,常常将两种杀螨特点不同的药剂加工成杀螨剂混合剂。
如5%阿维·达乳油和22%炔螨·噻螨乳油等。
第 44 卷第 3 期 Vol.44 No.32023 年 9 月Sept. 2023农业科学研究Journal of Agricultural Sciences葡萄霜霉菌对五种主要杀菌剂的抗药性研究进展曹爽,尹晓,马金龙,于泽洋(宁夏大学农学院,宁夏银川750021)摘要:葡萄霜霉病是葡萄种植过程中发生的主要病害之一,在冷凉潮湿、多雨多雾的天气条件下易于发病。
葡萄霜霉病的防治主要以化学防治为主,随着葡萄霜霉菌抗药性问题的出现导致化学防治效果不佳。
结合前人的研究,概述葡萄霜霉病的发生、葡萄霜霉菌对5种主要杀菌剂的抗药性、植物病原菌产生抗药性的机理、葡萄霜霉菌抗药性的检测方法,以期为葡萄霜霉菌的抗药性治理提供理论依据。
关键词:霜霉菌;杀菌剂抗性;作用机理;检测方法中图分类号:S482 文献标志码:A葡萄霜霉病(grape downy mildew)是葡萄的主要病害之一,是由葡萄霜霉菌(Plasmopara viticola)侵染所引起的,对葡萄的产量和品质等均有影响。
葡萄霜霉菌是卵菌的一种。
生产上对葡萄霜霉病的防治措施包括夏季修剪、提高结果部位、合理负载量、科学肥水管理、冬季清园以及采用避雨栽培等措施,但这仅能在一定程度上减轻葡萄霜霉病的危害。
化学药剂防治具有见效快、效果显著、操作方便等特点,目前防治葡萄霜霉病依然过度依赖化学杀菌剂的使用[1-2]。
化学药剂防治效果良好,可有效控制葡萄霜霉病的蔓延,但随着用药频繁和用药浓度加大,葡萄霜霉菌会产生抗药性。
本文从葡萄霜霉病的发生、葡萄霜霉菌对5种主要杀菌剂的抗药性、植物病原菌产生抗药性的机理以及葡萄霜霉菌抗药性的检测方法4个方面进行系统分析和总结,以期为葡萄霜霉菌抗药性的防治提供理论依据。
1 葡萄霜霉病的发生葡萄霜霉病在1834年首次在北美洲发现,后陆续传入欧洲大陆。
1899年在我国首次发现并记录此病,此后在我国各葡萄产区均有发生,其潜育期短,在雨水充沛条件下传播迅速,暴发性强,为害严重时造成巨大经济损失[3]。
龙源期刊网
内吸性杀菌剂
作者:李丽
来源:《农业知识·乡村季风》2016年第06期
产品特点
该产品是一种具有保护和治疗作用的内吸性杀菌剂,对病菌具有极强的杀灭效果,用药后可被根、茎、叶部吸收,并能在短时间内传达到作物的叶片和果实表皮内部,迅速进入病斑及周围,杀死病菌,有效控制病斑的发展,起到突出的治疗效果。
持效期长,对作物安全,不易产生抗药性,可与多种农药混配使用。
适用作物
用于防治小麦、水稻、玉米病害。
注意事项
1.首次使用请在农业技术人员指导下进行。
2.喷雾时力求均匀周到,包括叶片正反面喷透,以药液微有下滴为宜。
3.配药施药时严格遵守农药安全使用准则,应穿戴保护性衣物、手套、口罩等。
施药时不进食、不饮水、不吸烟。
施药后应清洗全身及换洗衣物。
应避免药剂接触皮肤和眼睛,不要直接接触被药剂污染的衣物,不要吸入药剂气体和雾滴。
4.不要因处理废药液而污染水源和水系,注意不要污染食物和饲料。
安徽农学通报2023年03期种子·肥料·农药·农膜作者简介江川安(1981—),男,安徽芜湖人,助理工程师,从事农药研究工作。
收稿日期2022-03-24精甲霜灵、咯菌腈与嘧菌酯复配对水稻烂秧病的室内活性及种衣剂田间药效试验研究江川安金立汤钟祥(安徽丰乐农化有限责任公司,安徽合肥231600)摘要针对水稻烂秧病防控,采用室内生物测定方法开展了精甲霜灵、咯菌腈、嘧菌酯及其不同配比组合对水稻烂秧病的联合毒力以及田间药效试验,明确最佳药剂配比,为药剂开发提供科学指导。
结果表明,精甲霜灵、咯菌腈、嘧菌酯配比为1∶1∶2时,混剂对水稻烂秧病防治增效作用最为明显,增效系数为6.13。
田间药效结果显示,4%精甲霜灵·咯菌腈·嘧菌酯种子处理悬浮剂对水稻晚疫病烂秧病效果较好,制剂量为600mL/100kg 种子,防效可达到80%以上,适宜施药时期为水稻播种前包衣,推荐有效成份使用剂量为18~24g/100kg 种子(制剂量为450~600mL/100kg 种子)。
关键词水稻;烂秧病;精甲霜灵;咯菌腈;嘧菌酯中图分类号S435文献标识码A文章编号1007-7731(2023)03-0095-04水稻烂秧病是水稻生产中发生最普遍、危害性较大的一种病害。
由绵腐菌属引起,在我国各水稻种植区均有发生。
精甲霜灵属于苯基酰胺类,为高效、低毒、低残留、持效期长、施药方式多样化、保护与治疗效果均优异的广谱内吸杀菌剂,可以作茎叶处理、种子处理和土壤处理,对霜霉菌、疫霉菌、腐霉菌等引起的作物病害有显著的防治效果。
咯菌腈杀菌谱非常广,从镰刀菌、雪腐病菌、链格孢菌到立枯病菌、褐腐病菌等,除了卵菌之外的大多数植物病原真菌,都是咯菌腈的靶标。
嘧菌酯是线粒体呼吸抑制剂,即通过抑制细胞色素b 和C1间电子转移而抑制线粒体的呼吸,其杀菌谱广,对晚疫病等真菌病害有良好的活性[1-3]。
为进一步明确精甲霜灵、咯菌腈和嘧菌酯联合对水稻烂秧病作用效果和复配后的田间应用效果,采用室内生物测定方法开展了精甲霜灵与嘧菌酯及其不同配比组合对水稻烂秧病作用效果病原菌的联合毒力以及田间药效试验,为药剂开发提供科学指导。
作者:祝明亮罗义勇李梅云杨金奎张克勤来源:《烟草农业科学》摘要:植物病原真菌的抗药性特别是对内吸性杀菌剂的抗药性是植物病害化学防治中的一个非常严重的问题,它给多种农作物和经济作物的病害防治带来了严重困难,给国家和农民造成了大量经济损失。
研究植物病原真菌抗药菌的生物学特征以及它们的抗药机制对于改进化学农药使用策略,延缓抗药性的产生,推进植物病害的化学防治,具有十分重要的理论和实践意义。
本文综述了近年来国内外植物病原真菌对内吸性杀菌剂抗药性研究在抗性菌和敏感菌的生物学特征、内吸性杀菌剂对植物病原真菌的作用机制、植物病原真菌对内吸性杀菌剂的抗药机制、抗性菌株的分子检测技术以及抗药性利用等方面的成果。
关键词:植物病原真菌;内吸性杀菌剂;抗药性;研究进展植物病原真菌对杀菌剂的抗药性是指本来对药剂敏感的植物病原真菌,由于基因突变或其它原因出现了药物敏感性下降的现象。
尽管早在1954年James等[1]就提出了这一现象,但是这一问题直到20世纪60年代末才引起人们真正重视。
1969年,由于内吸性杀菌剂苯来特(Benomyl)在生产上大量地使用,首先在黄瓜白粉病菌(Sphaerotheca fuliginea)上产生了抗药性,随后又在其它几十种病原真菌上产生了抗药性,并且有越来越严重的趋势。
到目前为止,用于防治植物病原真菌的杀菌剂种类已经很多,与抗药性问题关系密切的有有机硫杀菌剂、内吸性杀菌剂和抗菌素类杀菌剂,特别是内吸性杀菌剂的抗药性问题最为严重。
由于杀菌剂抗药性问题的迅速出现和发展,给植物病害的防治带来了越来越大的困难,给农业造成了重大的经济损失,引起了世界各国政府和农民的普遍关注。
为了有效防止或延缓杀菌剂抗药性的产生,许多国家和地区相继开展了对杀菌剂的抗药性研究。
人们首先对敏感菌和抗药菌的生物学特征进行了大量的对比研究,试图在此研究的基础上发现抗性病原真菌的抗药机制,最终来指导农药的合理使用和病菌的有效防治。
经过大量研究人员的辛苦努力,在植物病原真菌对杀菌剂抗药性问题的研究上,尤其是对内吸性杀菌剂的研究上,目前已取得了较大的进展。
1. 抗性菌和敏感菌的生物学特征对比抗性菌生物学特征即抗性菌的越冬、越夏、生长、繁殖和对环境的适应力等方面的特征。
许多情况下,抗性菌由于遗传变异等原因,在生物学特征上表现出与敏感菌有所不同,如在菌丝生长、产孢、与温度或渗透压的关系、致病力等方面的差异,这些差异影响着抗性菌在自然界与敏感菌的竞争力。
当抗性菌的竞争力强时,比较容易形成抗药性群体;反之,只要降低选择压力,就不易形成抗药性群体。
此外,有些抗性菌对同类或不同类的杀菌剂具有交互抗药性或负交互抗药性。
因此,人们可以根据抗性菌的特点,判断合理的用药策略,延缓或阻止抗药性群体的形成。
1.1 菌丝生长、产孢、孢外酶活力和致病力等方面的差异据研究报道,大多数抗药性病原真菌突变的适合度低于敏感菌。
抗药性菌株表现出菌丝生长速率慢,产孢量和产孢密度低,孢子萌发能力弱且滞后,孢外酶活力低,致病力弱,甚至完全失去致病力等特征[2,3]。
然而并不是所有的抗药性菌株的适合度都低于敏感菌,有些抗药性菌株具有很高的适合度,与野生敏感菌没有差异,甚至比敏感菌还高。
纪明山等研究灰霉病菌(Botrytis cinerea)对多菌灵(Carbendazim)和乙霉威(Dithofencarb)抗药性时,发现抗药菌与敏感菌在生长速率、产孢能力及致病力等方面无显著差异[4]。
徐作珽等报道蔬菜灰霉病菌对速克灵(Procymidone)的抗性突变体表现出较强的适应性,能正常生长繁殖产孢和侵染致病,且生存竞争能力较敏感菌强[5]。
我们在CA平板上测定烟草黑胫病菌对甲霜灵高抗、低抗和敏感菌株生长速率时发现,高抗菌株的生长速率较低抗和敏感菌株更快,而且高抗菌株仍然保持了较高水平的致病力。
1.2 交互抗药性与负交互抗药性交互抗药性是指植物病原真菌对一种杀菌剂产生了抗药性,而对另一种从未使用过但作用机制相似的药剂也产生了抗药性的现象。
与交互抗药性相反,负交互抗药性是指植物病原真菌对某种杀菌剂产生了抗药性,而对另一种从未使用过的药剂敏感性更强的现象。
交互抗药性和负交互抗药性在同类或不同类的杀菌剂之间普遍存在[6,7]。
1.3 多效性多效性是指抗药菌的抗性基因可以同时引起其它的性状,如对低温、高温或渗透压的敏感性。
Thomas等在研究酿酒酵母(Saccharomyces cerevisiae)对苯来特的抗药性时,发现在173个抗性菌株中,有6个菌株对低温敏感,3个菌株对生长温度敏感[8]。
在研究植物病原真菌对二甲酰亚胺类杀菌剂(Dicarboximides fungicide,DCFs )的抗药性时,粗糙脉孢菌(Neurospora crassa)os-1型渗透压突变菌表现出对DCFs的抗药性[9]。
2. 内吸性杀菌剂的作用机制与植物病原真菌的抗药机制内吸性杀菌剂是指能在植物发病后,通过内吸或渗透作用进入植物体内,抑制或杀死病原菌,使病株不再受害、恢复健康的药剂,主要有以下几类:苯并咪唑类(Benzimidazoles),二甲酰亚胺类,有机磷类(Organophosphates),苯酰胺类(Phenylamides),甾醇生物合成抑制剂类(Sterol Biosynthesis Inhibitors,SBIs )和甲氧基丙烯酸酯类(Strobilurins)(表 1)。
表 1 内吸性杀菌剂的种类、杀菌机制及病原菌抗药机制2.1 苯并咪唑类苯并咪唑类杀菌剂是最早也是最成功的内吸性杀菌剂,由于该类杀菌剂大量和长期的使用,病原真菌对它的抗药性问题越来越严重。
在真菌对杀菌剂抗药机制的研究中,真菌对苯并咪唑类杀菌剂抗药机制的研究最系统、最深入。
该类杀菌剂的杀菌机制为:首先结合构成纺缍丝的微管蛋白,使微管的形成受阻,从而破坏纺缍丝的功能,阻碍细胞的正常有丝分裂,使菌株不能正常的生长。
研究表明,不同病原真菌对苯并咪唑类杀菌剂抗性的主要机制是病原真菌的β-微管蛋白与该类杀菌剂的结合力下降,而杀菌剂同微管蛋白的亲和性的改变又与β-微管蛋白的结构变化有关。
分子生物学研究表明,在抗药菌β-微管蛋白的192-202个氨基酸上会发生基因突变,这类突变会导致β-微管蛋白的三维构象发生变化,从而阻止药剂与β-微管蛋白的结合,使病原真菌产生抗药性。
对田间抗药菌研究表明其突变位点主要集中在氨基酸的198 位和200 位[10],其它位点上的突变往往导致竞争力的减弱,因而在田间不易出现。
但玉蜀黍赤霉菌(Gibberella zeae)[11]和马铃薯干腐病菌(G. pulicaris)[12]对苯并咪唑类杀菌剂抗性菌的β-微管蛋白基因并没有发生改变,说明在不同病菌中或者在镰刀菌中对该类药剂发生抗药性突变的位点不同。
2.2 二甲酰亚胺类二甲酰亚胺类杀菌剂是20世纪70年代初推出的一类广谱性杀菌剂。
随着这类杀菌剂的大规模使用,很多植物病原真菌都对其产生了抗药性。
这类杀菌剂对真菌的孢子萌发、菌丝生长均有抑制作用,导致菌体形态改变,菌丝顶端产生了不正常的分枝和肿胀。
另外,在该类杀菌剂的处理下,病原真菌细胞内的甘油,过氧化氢酶和超氧化物歧化酶等内源物的水平会显著提高。
关于DCFs作用于病原真菌的机制,Lyr在1987提出了一种假说――自由基介导的细胞毒假说。
该假说认为DCFs诱导菌体产生还原态的氧自由基,从而攻击细胞膜上的不饱和脂肪酸,引起细胞膜脂质的过氧化反应,最终导致包括改变细胞膜通透性等一系列破坏细胞的连锁反应[13]。
关于这一假说是否成立,还存在着不同的观点。
而真菌对该类杀菌剂的抗药机制,还没有得到确切的结果。
目前,研究者们普遍认为是这些病原真菌细胞内的双组分组氨酸蛋白激酶N端的6个90氨基酸重复区的突变介导着对该类杀菌剂的抗药性(图 1)[14]。
根据抗药菌株同时具有对渗透压敏感的特征[9],研究者们克隆了很多真菌的双组分组氨酸蛋白激酶基因,并且发现:抗药菌的N端6个90氨基酸重复区中存在着碱基的突变或缺失,从而造成氨基酸或开放阅读框的改变,所以该氨基酸重复区的突变可能是抗药性产生的原因[15,16]。
但植物病原真菌对DCFs产生抗药性的机制,还需要更进一步的研究。
图 1 双组分组氨酸蛋白激酶结构示意图2.3 有机磷杀菌剂有机磷杀菌剂是指在化学结构中含有“C-P”键的抗菌化合物,根据它们的抗菌谱可分为两类:防治白粉病的磷酰胺类(Phosporothionate)和防治稻瘟病的硫代磷酸酯类(Phosphorothiolate)。
关于这两类杀菌剂的杀菌机制,研究表明,两者各不相同。
磷酰胺类杀菌剂的作用机制以定菌磷(Pyrazophos)为例。
定菌磷在菌体内能够进一步转化成一种更毒的分子,6-乙氧基羰基-5-甲基吡唑基并[1,2a]嘧啶(pp),从而抑制真菌对氧分子的吸收。
硫代磷酸酯类杀菌剂的作用机制以异稻瘟净(Iprobenfos)为例。
异稻瘟净的分子结构与菌体内磷酯酰乙醇胺(PE)相似,可以替代PE 接受δ-腺苷高半胱氨酸提供的甲基,阻止PE转化为磷酯酰胆碱,破坏菌体生物膜的结构,抑制菌体生长。
关于磷酰胺类杀菌剂的抗药性还很少见,到目前为止只在黄瓜白粉病菌中发现对定菌磷的抗性菌株,而关于硫代磷酸酯类杀菌剂的抗药性,研究认为,室内获得的稻梨孢菌(Piricularia oryzae)高抗突变体的抗药机制是裂解药剂“S-C”键以致使其失去生物活性。
高抗菌一般生存竞争能力较低,所以在自然界很少发生。
田间产生的中抗菌的抗药机制可能与药剂靶点修饰或细胞膜透性改变有关。
2.4 苯酰胺类苯酰胺类杀菌剂是防治霜霉目真菌的专用药剂,具有显著的保护、治疗和铲除作用,广泛应用于马铃薯和番茄晚疫病的防治。
由于该类杀菌剂对病菌作用位点单一,因而很容易导致病菌产生抗药性。
由于抗药性产生而导致药效降低的事例已屡见不鲜。
该类杀菌剂在寄生组织内抑制菌丝生长和吸器形成,但不影响病原菌孢子囊的直接和间接萌发以及游动孢子的萌发[17]。
关于该类杀菌剂的作用机制,研究结果表明,它们是通过阻止尿苷掺入RNA来干扰核酸的合成(特别是γ-RNA 的合成)来实现的。
多种病原真菌试验结果表明并不是全部的RNA合成受阻。
通过研究发现,该类杀菌剂对合成细胞内核苷酸的三种RNA聚合酶有选择性抑制作用,其中对合成γ-RNA的聚合酶A的毒力最大,使核糖体不能正常工作。
关于不同真菌对苯酰胺类杀菌剂的抗性遗传机制,许多实验结果表明是属于单基因变异。
通过对莴苣霜霉病菌(Bremia lactucae)、致病疫霉(Phytophthora infestans)和寄生疫霉(P. parasitica)等的研究发现,对苯酰胺类杀菌剂的抗性是由单个主效基因控制的,其中在莴苣霜霉病菌和致病疫霉中,抗性基因为不完全显性。