绿僵菌简介
- 格式:ppt
- 大小:23.50 KB
- 文档页数:5


绿僵菌菌剂培养及田间生物防治研究杨文彬(山西省医药与生命科学研究院,山西太原 030006)摘 要:绿僵菌作为重要的生物农药制剂,研究及应用非常广泛。
本研究通过试验探索了固体培养绿僵菌L8500-2x菌株的最佳培养基配方及工艺条件,进行了田间试验示范推广,为有效利用绿僵菌进行田间合理的防治提供了科学参考。
关键词:绿僵菌;菌剂培养;生物防治中图分类号:S476.1 文献标识码:A DOI:10.11974/nyyjs.20170533028绿僵菌(Metarhizium anisopliae)是重要的昆虫病原真菌之一[1],将其作为主要生物活性成分的制剂,具有不污染环境、无残留、易生产、致病力强、持效期长、应用效果好、对非靶标生物安全等优点,其寄主范围广,可在8目30科200余种农林害虫上寄生,其应用前景非常广阔[2,3]。
近年来,由于化学农药的大量使用,食品安全和健康问题、环境保护问题倍受重视,生物农药的研发及推广具有深刻意义[4]。
目前,绿僵菌生物农药的生产普遍采用液固双相发酵工艺,其固体载体多为大米和稻壳,国外绿僵菌生产的固体载体多为稻米[5]。
绿僵菌制剂的剂型主要有粉剂、可湿性粉剂等,其助剂的成分和配比是当前研究热点。
本研究为了大量生产绿僵菌菌剂用于田间农作物害虫的防治,先采用液体发酵培养绿僵菌L8500-2x菌剂,再采用固体发酵培养绿僵菌L8500-2x菌粉,进行田间试验示范推广,可为有效利用绿僵菌防治农作物病害提供参考,为适应绿色农业的发展要求,减少高毒、高残留化学农药的使用提供科学方法。
1试验材料与方法1.1 绿僵菌L8500-2x菌株固体发酵培养载体的筛选1.1.1试验材料绿僵菌培养载体:麦麸、硅藻土、磷酸钙、锯末、谷糠、玉米粉;试验害虫是菜青虫2代幼虫。
1.1.2试验方法[6]利用麦麸、硅藻土、磷酸钙、锯末、谷糠、玉米粉等廉价物质作为基质进行绿僵菌L8500菌剂的生产,观察不同供试载体对绿僵菌菌丝生长、产孢量和孢子萌发率的影响,经对比试验,确定固体培养绿僵菌L8500-2x 菌种的最佳培养基配方及工艺条件[7]。
6维度环境与健康网
绿僵菌:广泛使用的生物杀虫剂
编译:菁菁 6维度环境与健康网
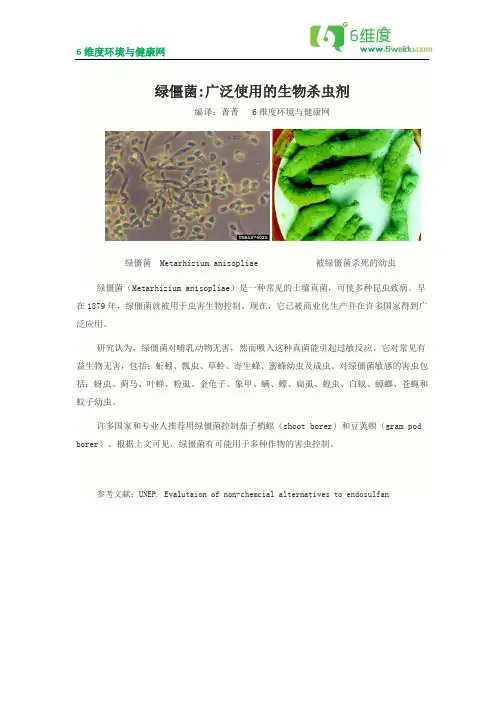
绿僵菌 Metarhizium anisopliae 被绿僵菌杀死的幼虫 绿僵菌(Metarhizium anisopliae)是一种常见的土壤真菌,可使多种昆虫致病。
早在1879年,绿僵菌就被用于虫害生物控制。
现在,它已被商业化生产并在许多国家得到广泛应用。
研究认为,绿僵菌对哺乳动物无害,然而吸入这种真菌能引起过敏反应。
它对常见有益生物无害,包括:蚯蚓、瓢虫、草蛉、寄生蜂、蜜蜂幼虫及成虫。
对绿僵菌敏感的害虫包括:蚜虫、蓟马、叶蝉、粉虱、金龟子、象甲、螨、蠓、扁虱、蝗虫、白蚁、蟑螂、苍蝇和蚊子幼虫。
许多国家和专业人推荐用绿僵菌控制茄子梢螟(shoot borer)和豆荚螟(gram pod borer)。
根据上文可见,绿僵菌有可能用于多种作物的害虫控制。
参考文献:UNEP. Evalutaion of non-chemcial alternatives to endosulfan。


13F聚焦Focus“P 专业视角Perspective微生物杀虫剂的研发及田间防效 微生物杀虫剂的研发面临诸多问题:一是害虫种类多;二是抗药性上升,用量增大,产品市场寿命短;三是农药登记、生产、销售、使用等环节的监管越来越严格;四是防控理念变化要求我们不仅要保护寄生性、捕食性等天敌,还要不伤害其它益虫和非靶标生物。
综上可以看出,广谱、无抗性、安全、持续控害成为新杀虫剂的必备特点。
而“智慧”真菌杀虫剂——绿口/《农化市场十日讯》编辑部整理僵菌CQMa 421就具有杀害虫、护益虫、无抗性的特性。
通过筛选绿僵菌广谱菌株在稻田上进行试验发现,绿僵菌CQMa 421侵染水稻三种主要害虫,即稻纵卷叶螟、稻飞虱和二化螟,但不侵染稻田主要天敌。
目前研发出的产品类别包括可分散油悬浮剂和可湿性粉剂,可以应用于水稻、茶叶、蔬菜、烟草等作物上。
截至目前,农业农村部已登记的绿僵菌CQMa 421可分散油悬浮剂有1个,主要防治水稻稻纵卷叶螟、稻飞虱、二化螟、叶蝉;茶树茶小绿叶蝉;甘蓝黄条跳甲;烟“智慧”真菌杀虫剂——绿僵菌CQMa 421/生物源/14““草和桃树蚜虫等虫害。
其扩作登记包括豇豆上的甜菜夜蛾、苦瓜上的蚜虫、柑橘上的木虱、茄子上的蓟马等10余种害虫。
截至目前,农业农村部已登记的绿僵菌CQMa 421可湿性粉剂有1个,主要针对水稻稻纵卷叶螟、稻飞虱和二化螟。
综合2010-2015年,在全国8个省区进行应用“绿僵菌CQMa 421”油悬浮剂分别防控稻飞虱、稻纵卷叶螟和二化螟的田间试验和大面积应用示范,以及2018年在四川、山东进行的防治小麦蚜虫的田间试验,结果表明,与化学杀虫剂相比,“绿僵菌COMa 421”防治水稻害虫和小麦蚜虫死亡慢、死亡率低,但产量无显著差异。
绿僵菌CQMa 421应用的技术要点 1、保证药剂与害虫的接触,接触机会越多,虫害致死速度越快。
2、使用器械:可以选用常规喷雾器;静电喷雾器用于低容量喷雾、常量喷雾;超低容量喷雾器用于低容量喷雾、超低容量喷雾;无人机喷雾器用于低容量喷雾、超低容量喷雾。

实验名称:绿僵菌抗药性菌株的驯化选育
摘要
绿僵菌(Metarhizium anisopliae)半知菌类、丛梗菌目、丛梗霉科、绿僵菌属,具有广谱性。
绿僵菌对寄主的入侵是寄主与病原菌之间的生理生化作用的综合结果,可通过体壁、气门、消化道等多种途径侵染宿主,其中体壁途径是主要方式。
本文研究嘧霉胺驯化诱导绿僵菌产生抗性菌株,研究绿僵菌对嘧霉胺的抗性风险。
绿僵菌在含嘧霉胺的培养基上经逐代驯化,获得了抗嘧霉胺的菌株R1、R2、R3、R4,经抗药性水平测定,R1、R2、R3、R4的抗性倍数分别为10ug/ml,100ug/ml,500ug/ml,1000ug/ml四株抗药性菌株均为对嘧霉胺的抗性菌株。
本文比较了抗药性菌株和敏感性菌株的生物学性状。
采用菌丝生长速率法测定了抗性菌株与敏感菌株的菌丝生长速率,结果表明经7d培养抗性菌株菌落直径在0.5-1.1cm之间,而敏感菌株为1.3cm,生长速率存在明显差异;在及抗性菌株与敏感菌株的产孢量大约都在3.6×107个/ml,说明室内诱导的高抗突变体不能正常生长,菌株对药剂的抗药性水平与菌株产孢能力没有相关性,而是由菌株本身决定的。
本试验初步研究了抗性菌株的遗传稳定性,抗性菌株的第零代到第六代的的结果表明绿僵菌的抗药性菌株与敏感菌株的不同转代的有差异性,从第零代第三代到第九代逐渐减小,第零代与第三代和第六代下降明显,第三代与第六代下降幅度很小,但始终保持着对嘧霉胺的高等抗性,表明抗性菌株具有良好的遗传稳定性。
关键词:绿僵菌;嘧霉胺;抗性菌株;生物学特性;遗传稳定性。

金龟子绿僵菌(Metarhizium anisopliae) tetraspanin基因的克隆与序列分析贺闽1,夏玉先1*1重庆大学生物工程学院基因工程研究中心,重庆(400044)摘要:Tetraspanin蛋白是一类跨膜四次的膜蛋白,本文测序分析金龟子绿僵菌侵染蝗虫表皮阶段的cDNA文库,获得了金龟子绿僵菌的tetraspanin cDNA序列,以cDNA为探针从金龟子绿僵菌的DNA文库中克隆到了tetraspanin基因,并对tetraspanin蛋白进行了结构与进化分析, Soutern-blot分析表明tetraspanin基因在金龟子绿僵菌基因组中以单拷贝形式存在。
关键词:金龟子绿僵菌;tetraspanin;cDNA;DNA;进化分析。
中图分类号:1. 引言昆虫病原真菌是昆虫主要的自然致死因子,真菌生防剂具有触杀性、流行性、对环境安全和不易产生抗药性等特点,因此利用虫生真菌防治害虫越来越受到重视。
金龟子绿僵菌作为一种重要的昆虫病原真菌,国内外对其致病机理进行了大量研究,鉴定出了一些重要毒力基因,涉及真菌粘附并穿透宿主体壁、利用宿主营养、合成毒素以及逃避宿主免疫反应等[1,2,3,4,5,6]。
附着胞是虫生真菌入侵寄主过程中形成的重要侵染结构,能够分泌体壁降解酶和提供机械压力帮助侵染钉穿透昆虫体壁,附着胞的产生对于建立病原真菌与寄主关系是至关重要的。
因此,阐明附着胞形成分化的分子机制,能够为开发安全高效的真菌杀虫剂、挖掘昆虫病原真菌应用潜力提供科学依据。
Tetraspanin广泛表达于真核生物,是一类含204-344个氨基酸的细胞表面糖蛋白。
哺乳动物中已发现的tetraspanin家族蛋白有32个,果蝇中有37个,秀丽隐杆线虫中有20个,真菌中有4个。
在这众多的家族成员中,来自不同物种的tetraspanin很少存在同源性,但这些家族成员蛋白二级结构具有一定保守性,都由四个疏水跨膜区、胞外区的小环(EC1)和大环(EC2),以及两个短的胞内末端组成[7]。



绿僵菌菌剂培养及田间生物防治研究作者:杨文彬来源:《农业与技术》2017年第10期摘要:绿僵菌作为重要的生物农药制剂,研究及应用非常广泛。
本研究通过试验探索了固体培养绿僵菌L8500-2x菌株的最佳培养基配方及工艺条件,进行了田间试验示范推广,为有效利用绿僵菌进行田间合理的防治提供了科学参考。
关键词:绿僵菌;菌剂培养;生物防治中图分类号:S476.1 文献标识码:A DOI:10.11974/nyyjs.20170533028绿僵菌(Metarhizium anisopliae)是重要的昆虫病原真菌之一[1],将其作为主要生物活性成分的制剂,具有不污染环境、无残留、易生产、致病力强、持效期长、应用效果好、对非靶标生物安全等优点,其寄主范围广,可在8目30科200余种农林害虫上寄生,其应用前景非常广阔[2,3]。
近年来,由于化学农药的大量使用,食品安全和健康问题、环境保护问题倍受重视,生物农药的研发及推广具有深刻意义[4]。
目前,绿僵菌生物农药的生产普遍采用液固双相发酵工艺,其固体载体多为大米和稻壳,国外绿僵菌生产的固体载体多为稻米[5]。
绿僵菌制剂的剂型主要有粉剂、可湿性粉剂等,其助剂的成分和配比是当前研究热点。
本研究为了大量生产绿僵菌菌剂用于田间农作物害虫的防治,先采用液体发酵培养绿僵菌L8500-2x菌剂,再采用固体发酵培养绿僵菌L8500-2x菌粉,进行田间试验示范推广,可为有效利用绿僵菌防治农作物病害提供参考,为适应绿色农业的发展要求,减少高毒、高残留化学农药的使用提供科学方法。
1 试验材料与方法1.1 绿僵菌L8500-2x菌株固体发酵培养载体的筛选1.1.1 试验材料绿僵菌培养载体:麦麸、硅藻土、磷酸钙、锯末、谷糠、玉米粉;试验害虫是菜青虫2代幼虫。
1.1.2 试验方法[6]利用麦麸、硅藻土、磷酸钙、锯末、谷糠、玉米粉等廉价物质作为基质进行绿僵菌L8500菌剂的生产,观察不同供试载体对绿僵菌菌丝生长、产孢量和孢子萌发率的影响,经对比试验,确定固体培养绿僵菌L8500-2x菌种的最佳培养基配方及工艺条件[7]。
![金龟子绿僵菌IPPMHBHC-7及其应用[发明专利]](https://uimg.taocdn.com/a7f3c5eeeff9aef8951e06a9.webp)
专利名称:金龟子绿僵菌IPPMHBHC-7及其应用
专利类型:发明专利
发明人:王广君,郭隆隆,李贝贝,王晔青,涂雄兵,农向群,张泽华
申请号:CN201811184614.4
申请日:20181011
公开号:CN109401981A
公开日:
20190301
专利内容由知识产权出版社提供
摘要:本发明属于生物技术领域,具体涉及一种金龟子绿僵菌IPPMHBHC‑7及其应用。
金龟子绿僵菌(Metarhizium anisopliae)IPPMHBHC‑7,保藏编号为CGMCC No.15999,其对于亚洲小车蝗、毛足棒角蝗具有良好的半数致死时间和僵虫率。
申请人:中国农业科学院植物保护研究所
地址:100093 北京市海淀区圆明园西路2号
国籍:CN
代理机构:北京法信智言知识产权代理事务所(特殊普通合伙)
代理人:刘静荣
更多信息请下载全文后查看。
白僵菌与绿僵菌介绍白僵菌与绿僵菌介绍白僵菌白僵菌是一种半知菌类的虫生真菌,具有营养器官--菌丝和繁殖器官--分生孢子,菌丝有横隔有分枝。
白僵菌的分布范围很广,从海拔几米至2000多米的高山均发现过白僵菌的存在,白僵菌可以侵入6个目15科200多种昆虫、螨类的虫体内大量繁殖,同时不断产生白僵素(大环脂类毒素)和草酸钙结晶,这些物质可引起昆虫中毒,使体液发现机能发生变化,打乱新陈代谢以致死亡。
白僵菌分类:常见白僵菌共有三种:球孢白僵菌、小球孢白僵菌、卵孢白僵菌。
球孢、小球孢、卵孢三种白僵病菌,其不同发育阶段,虽具有共同的形态特征,但也存在一定的差异。
其区别是:球孢和小球孢白僵菌,分生孢子虽都为球形或卵形(约各50%),均生于对称成直角的茸状产孢细胞顶端,分生孢子梗呈直角分支聚成集团。
所不同的是前者孢子大,一般2.5—3.0微米,菌落呈平绒状,在明胶培养基本逐渐从白色变到乳白色,而在马铃薯琼脂培养培养的底部无色或淡黄色。
后者孢子较小,一般为2-2.5微米,菌落白色至乳白色初为疏茸状或棉絮状,后期形成乳粉状的孢子层,不使马铃薯琼脂培养基斜面变色,但在清淅明交培养基上呈粉红色,颜色很不显著,而且10天后便消失。
卵孢白僵菌不同于前两种的主要特征,是其分生孢子大多是亚园形或椭圆形,而且分生孢子开差较大,一般为2.0-6.0×1.5-4微米,生于产孢细胞顶端新延伸的“乙”形丝形器上,菌落表面高低不同,白色,初为茸毛状或棉絮状,后期形成粉状,在明胶培养基的底部呈深红色至桑色,某些菌丝命名马铃薯培养基呈不同程度浅紫色至红色。
目前在农业部登记的白僵菌有两种:一、白僵菌(beauveria)名称又为虫生真菌球孢白僵菌。
是生物源杀虫剂。
原药外观为乳白色粉末,制剂外观为乳黄色粉状物。
二、球孢白僵菌(beauveria bassiana)为植物源杀虫剂。
本产品主要通过触杀作用,可以穿透昆虫体壁,在昆虫体内增殖,进而致死目标害虫。
绿僵菌的研究进展2005年4月Apr.2005热带农业科学CHINESEJOURNALOFTROPICALAGRICULTURE第25卷第2期V o1.25,No.2绿僵菌的研究进展①代鹏②宋妍许天委谢玉平黄俊生③(中国热带农业科学院环境与植物保护研究所海南儋州571737)摘要综述了近年来国内外对绿僵菌属的分类,绿僵菌入侵寄主过程中的相关基因的研究,剂型开发等方面的概况.关键词绿僵菌;分类现状;相关基因;剂型开发分类号$432.44;Q949.32;Q78AdvancesofResearchonMetarhiziumspp. DAIPengSONGYanXUTianweiXIEYupingHUANGJunsheng (EnviromentandPlantProtectionInstitute,CATAS,Danzhou,Hainan571737) AbstractThearticlesummarizestheresearchonclassingstatusofMetarhiziumspp.,correlati on geneinthebreakinginsectandthepreparationofMetarhiziumanisopliaeinrecentyears. KeywordsMetarhiziumspp;classifyingstatus;correlationgene;preparation真菌生防制剂具有触杀性,流行性,对环境安全和不易产生抗药性等特点.70%的昆虫真菌性病害,在害虫控制方面有巨大的潜力.对绿僵菌[Metarhiziumanisopliae(Metarhiziumspp.),Ma]的研究已有100多a的历史,但由于在发酵方法,剂型开发及环境制约等方面存在各种各样的问题,导致其防治效果不稳定,影响了其大规模的应用.直到1988年,金龟子绿僵菌制剂应用于澳洲甘蔗和牧草害虫的防治,其应用才算取得较大进展.应用绿僵菌防治地下害虫及非洲蝗虫也已取得初步成效,绿僵菌的研究和应用成了生物防治领域的一个热点.1绿僵菌属的系统分类现状11绿僵菌属的分类地位自从1879年MeqHHKOB首先大量繁殖绿僵菌防治奥地利丽金龟(,4nisopliaaustriacaHerbst)以来,绿僵菌就成为了害虫微生物防治的主角之一. 绿僵菌属(MetarhiziumSorokin)是Soroki13于1883 年建立的,它隶属于真菌门(Eumycota),子囊菌亚门(Ascomycotina),核菌纲(Pyren—mycetes),球壳菌目(Sphaeriales),麦角菌科(Clavisipitetaceae).自Sorokin建立绿僵菌属以来,先后发表10余个种.由于不同作者采用的标准不统一,故分类状况比较混乱.目前国际上被广泛接受的分类观点是Tulloch建立的,他将绿僵菌属定为2个种,即金龟子绿僵菌和黄绿绿僵菌,金龟子绿僵菌包括小孢变种和大孢变种….利用传统的分类方法很难区分绿僵菌属内各种群间的差异,而野生型菌株的遗传特性则提供了菌株间的遗传差异.现有资料表明,利用可溶性蛋白图谱能区分形态学上相似的种;酯酶同工酶区分绿僵菌突变株也明显表现出多态性,这可能与其遗传变异因子及环境因素有密切的关系.另外,分子标记方法用于绿僵菌的鉴定,分类,进化和其它各方面信息的研究,是当前和今后受重视的领域.①收稿日期:2004—12—30责任编辑/凌青根:// / rdnk@②代鹏(1978~),男,硕士,助理研究员.E-mai1:dapen008~163.tom.③通讯作者.一73—2005年4月热带农业科学第25卷第2期近年来,在DNA水平上研究遗传差异的技术早已被认可,并且已经成为研究虫生真菌群落遗传结构的有效方法.1.2绿僵菌属种间分类方法的应用限制性片段长度多态性(RFLP)和聚合酶链式反应(PCR)技术基础上的DNA随机扩增多态性RAPD方法,为研究绿僵菌的分类鉴定,系统发育,种内分型和地理型分布,及其生态学和流行学,提供了重要的检验手段.rDNA基因在进化中的高保守性和相关间隔区的多变性,使其成为研究分子系统中的最重要和应用最为广泛的工具之一.目前这项技术已广泛应用于绿僵菌的研究r2].Leal等用巢式PCR技术,将对不同地理来源和分离自不同寄主或土壤的40株绿僵菌菌株蛋白酶Prl基因扩增出来,然后用3种限制性内切酶(DdeI,HpaII和RsaI)消化扩增产物.酶切图谱共有15种类型,经聚类分析分为4组.在部分组的菌株间,其限制性酶切片断图谱和地理来源有一定的相关性[3].Fegen及Cobb等人于1993年同时首次将RAPD—PCR方法用于虫生真菌的研究.Fegen的研究发现,利用RAPD—PCR分析DNA片段长度的多态性可以得到相应的DNA遗传和进化等信息.Leal等用6种随机引物,对来自4个国家的20株绿僵菌进行RAPD研究.结果表明,多数菌株有特异RAPD—PCR图谱,一些菌株可表现明显的地理性, 另一些不同来源的菌株则不表现出地理差异r4,.我国学者刘兆伟,郭好礼等于1993年率先用Sanger法测定了几个种的绿僵菌的部分rDNA序列.结果表明,25S一635区域比其它2个区域具较高的突变率,可以准确地反映遗传关系[6].Drive和Milne的研究,是迄今为止最为详尽和完整的,基本解决了绿僵菌属的分类难题.他们共测定了121株绿僵菌菌株的ITS卜5.8S—ITS2 rDNA序列,其中26株有代表性菌株的序列数据在文中予以应用.综合Curran和他们的研究结果,建议将37株绿僵菌划分为3个种(金龟子绿僵菌,黄绿绿僵菌和白色绿僵菌),其中黄绿绿僵菌下分5个变种,金龟子绿僵菌下分4个变种.值得一提的是,他们将ITS1和ITS2的序列作为唯一的性状来进一74一行拉丁文描述,使ITS1和ITS2的序列成为绿僵菌分类最可靠的依据Ⅲ.2绿僵茵入侵过程中相关基因的研究昆虫病原真菌约有1000种,其中一些种类已成功用于蝗虫,粉虱,白蚁,蟑螂等害虫生物防治.近年研究较多的是金龟子绿僵菌(Ma),该菌含多种可降解昆虫外表皮的酶,已从中克隆到多种致病相关基因.2.1Prl基因有研究结果表明,Ma产生的昆虫表皮水解酶在高水平情况下与害虫发病机理有很大的关系.胞外弹性凝乳蛋白酶Prl基因是最早被克隆的昆虫表皮降解蛋白基因.该基因表达的2种蛋白PrlA和PrlB具有很强的分解昆虫体壁的功能,它主要降解酪蛋白,凝乳弹性蛋白,降血凝乳蛋白和胶原质蛋白等,在对昆虫的致病力中起着重要的作用.PrlA和PrlB分别位于金龟子绿僵菌不同染色体上,所编码的氨基酸序列有54%的同源性[8--10]. PrlA基因的一级结构和丝氨酸肽链内切酶——枯草杆菌蛋白酶具有相当高的同源性.PrlA与枯草杆菌蛋白酶K的氨基酸序列相似性达61%,但PrlA降解烟草天蛾体壁的能力是蛋白酶K的2倍…]. Screen等分析了PrlA基因启动子的序列,发现存在CREA和AREA结合位点.条带移位实验证实了构巢曲霉GST—CREA融合蛋白能结合CREA上3个位点中的2个.Joshi等利用差异显示RT—PCR得到1个新的类枯草杆菌蛋白酶基因(PrlB),核型分析表明,二者位于不同的染色体上,通过对绿僵菌白僵菌Prl基因的cDNA分析,发现PrlA和PrlB均首先被合成为1个大的前体蛋白,包括1个信号肽,1个前体肽和1个成熟的蛋白分子,它们的前体蛋白和成熟蛋白分子量大小均不相同.StLeger于1987年发现并于1993年克隆到了Prl基因.Prl蛋白一直被认为在绿僵菌侵染昆虫过程中具有不可代替的重要作用,但王成树等人于2000年首次报道了缺失Prl基因的野生型菌株依然保持了其毒力特性.野生型Ma菌株V275在初期单孢分离过程中发现了3株缺失弹性凝乳蛋代鹏等绿僵菌的研究进展白基因PrlA和PrlB的突变株,经嵌套PCR扩增和Southern杂交实验,证明了该基因的确已丢失.28srDNA保守序列分析比对证明了该突变体都是金龟子绿僵菌,但形态学和遗传与野生型差异明显.RAPD分析表明,二者之间的整体遗传相似性低于70%.生物测定发现,突变菌株对Tenebno molitor致死能力仅比野生型菌株低20%,而对Galleriamellone的致死能力却相当.因此,王成树指出,在人造培养基上培养的绿僵菌缺失Prl基因是一种常见现象,应该得到足够的重视[1.2.2Pr2基因在昆虫病原真菌中另一个重要的内切蛋白酶是丝氨酸类胰蛋白酶,也是一组碱性蛋白同工酶. Smithson等从绿僵菌液体培养基中通过阴离子交换色谱得到丝氨酸类胰蛋白酶Pr2,并对N一末端序列进行了测定[1.利用寡脱氧核糖核酸探针,从绿僵菌的基因文库中钓出该蛋白酶基因.经研究发现,该基因编码一个由254个氨基酸组成的蛋白质,含有2个长度分别为94和40bp的内含子.信号序列断裂分析表明,Pr2的同工型Tryl在它的N末端含有21个氨基酸的信号肽,产生一个含226个氨基酸成熟蛋白.Pr2基因编码的丝氨酸蛋白酶可以水解酪蛋白和白蛋白,但不能降解凝乳弹性蛋白.StLeger等人的研究结果表明,Pr2是Prl活性的4%~10%,而Cole等人的研究结果是5.4%~26%.液体培养发现,在接种绿僵菌后48h,即可检测到Pr2的活性,而到72h才检测到Prl的活性,因此认为,Pr2可能在Prl的诱导和激活方面起着重要作用n.2.3Pr3及Pr4基因StLeger在研究过程中发现,Pr3是一种酸性内切水解蛋白酶,pH5.0~5.5,等电点低,目前对其组成及作用机制仍不太清楚.Pr4表现出胰岛素的特性,但根据其特性和活性研究,将其划分为半胱氨酸蛋白酶,其活性是Prl的51%,远高于Pr2.2.4毒素毒素基因是编码"环状六肽毒素"合成酶的基因.通过研究不同来源的绿僵菌的"破坏素"的合成酶的基因序列,发现该类毒素中每一个氨基酸都有一个酶化的区域,每个域内有许多的保守序列.目前已发现了不少昆虫病原真菌毒素,但尚未开发出与毒素相关的商业制剂[1.Mollier将这些毒素分为两大类:低分子量毒素和高分子蛋白毒素.破坏素(destruxin)是Kodaira从金龟子绿僵菌[anisopliae(Metschn.)Sorokin]的培养液中分离到的低分子量的环缩肽.当时从滤液中分离出6 种环缩肽,其中2种的杀虫活性高,分别命名为destruxinA和destruxinB,二者结构类似,均由分丙氨酸,丙氨酸,撷氨酸,异亮氨酸和脯氨酸这5种氨基酸组成.破坏素对竹节虫,大蜡螟等昆虫表现出毒性.用破坏素进行体腔注射后,不同昆虫表现的症状不同,多数鳞翅目(Lepidoptera)昆虫表现为强直性麻痹.从金龟子绿僵菌中分离出的细胞松弛素(cytochalasins)是一种吲哚环结构毒素,细胞松弛素对哺乳动物有急性毒性,可抑制血细胞的运动,降低血细胞的吞噬能力[1.提纯破坏素主要是采用吸附法,即将绿僵菌培养液的滤液用7g/L的活性炭吸附,再用体积分数为50%的丁醛水溶液将活性物质从活性炭上洗脱下来,然后分离黄色相,真空喷雾浓缩,用苯充分萃提.苯液过中性氧化铝柱子,然后分别用含5%和10%(体积分数)乙醇的苯洗脱,分开收集,便可得到破坏素A(简称DA)和破坏素B(简称DB),苯油经重结晶即可得无色的DA和DB的晶体.李农昌以金龟子绿僵菌小孢变种(anisopliaeSorokinvat. anisopliaeTulloch)为材料,将其菌丝体捣碎,过滤浓缩后得黄色粘稠状粗提物,然后对粗提物进行薄层层析,最终也纯化到了破坏素[1.目前对昆虫病原真菌毒素研究较多的是小分子环缩肽类和可降解寄主体壁的酶类,对其研究主要还是集中在实验室内,直接应用于防治田间害虫的实例尚未见报道,主要是因为其发酵含量低,对哺乳动物有急性毒性,并且提取和制造工艺繁琐,目前还不能产生较好的经济效益.3绿僵菌剂型的开发研究3.1绿僵菌生产工艺的研究75—2005年4月热带农业科学第25卷第2期在绿僵菌生产工艺方面,单纯的液体发酵生产的孢子或菌体,因活性和耐储性降低而受到限制.主要采用传统的液固双相发酵生产工艺生产真菌孢子粉,固相发酵阶段存在搅拌不均,通透性差,发酵周期长和设备价格偏贵等问题.国内多采用浅盘,窗纱或无纺布开放式生产模式,空间利用率低,容易感染杂菌,产品质量不稳定,成为阻碍真菌生防制剂产业化发展的重要原因之一.另外,太阳光紫外线是一种影响金龟子绿僵菌田间使用效果的主要影响因子.Drauzio等人的研究结果表明,绿僵菌对紫外线相当敏感,且用PDA,PDAY等人工培养基生长的绿僵菌对紫外线的敏感度要低于直接用昆虫体培养的绿僵菌,其中稻谷类人工培养基能显着提高绿僵菌对紫外线的耐受性[1B]. Braga等提出,即使室外紫外线强度低于致死剂量,也会对真菌孢子的致病力及萌发速率产生一定影响n9j.3.2绿僵菌剂型的开发研究在长期的剂型研究过程中,真菌杀虫剂剂型研究取得了较快的发展.到目前为止,真菌杀虫剂主要剂型有粉剂,可湿性粉剂,乳剂,油剂,干菌丝,微胶囊等.其中,已大面积应用的绿僵菌剂型主要是粉剂,干菌丝和油剂等.据国外报道,醇烷氧基化合物也是一种有效的乳化剂.与水剂相比,油剂在低的相对湿度下更有利于孢子的萌发,同时在高温下也更能延长孢子的寿命.油剂还有利于孢子对疏水基质的吸附,如昆虫体壁或植物表面]. Inyang等研究了溶剂油对黄绿绿僵菌(anisopli oe)分生孢子活性的影响,结果表明,菜子油,花生油或色拉油是较好的溶剂油,制成的油剂(加有抗氧化剂)在17℃下贮藏127周后的孢子萌发率仍达到60%C.目前国外研究的真菌生防制剂大概有100多种,其中主要是真菌杀虫剂.除草剂,杀菌剂和杀线虫剂也在不断增加.杀虫制剂包括白僵菌14个,绿僵菌7个.其中,影响最大且最成功的范例,是英国CABIBioscience蝗虫生物防治国际合作项目(LUBILOSA)研制的杀蝗绿僵菌生物农药,已在非洲注册登记,投产,大面积用于沙漠蝗的防治,一76一施药l0~15d后防效达到90%以上,成为有效的蝗虫生防手段.澳大利亚蝗虫生物防治研究项目(CSIRO)研制出了杀蝗绿僵菌生物农药,并用于大面积草原蝗虫的防治,对蝗虫天敌等非目标生物安全.EcoScience绿僵菌产品(Bio—Path,Bio—Blast)用于防治蟑螂,白蚁;美国Ecoscien的绿僵菌产品(Bio—Path,Bio—Blast)防治蟑螂和白蚁;巴西和委内瑞拉的绿僵菌产品(Metaquino,Cobican)防治甘蔗沫蝉等等.我国对杀虫真菌制剂的研究,主要是重庆真菌农药研制工程中心获得的杀蝗绿僵菌孢子母粉和杀蝗生物制剂"100{L/mL杀蝗绿僵菌油悬浮剂",已在我国北方部分蝗虫发生省区使用和推广.参考文献1TullochM.ThegenusMetarhizium.Transactionof theBritishMycologicalSociety,1976,66:407~4112黄勃,樊美珍,李增智.绿僵菌属系统分类的研究进展.安徽农业大学,2002,29(2):169~1723LealSCM,BertioliDJ,ButtTM,eta1.Amp1ifi—cationandrestrictionendonucleasedigestionof thePrlgeneforthedetectionandcharactization ofMetarhiziumstrains.MycologyResearch,1997,101:257~2654CobbBD,ClarkonJM.Detectionofmolecular variationintheinsectpathogenicfungusMetarhiz—iumusingRAPD-PCR.FEMSMicrobiologyLetter, 1993,112:319~3245LealSCM.Characterizationofisolatesoftheen- tomopathogenicfugusMetarhiziumanisopliaeby RAPD-PCR.MycologyResearch,1994,98(9):1077~10886刘兆伟,郭好礼,KurtzmanCP.由部分核糖体RNA序列确定的绿僵菌属种系统发育关系.真菌,1994,13(2):139~1517DriverF,MilnerRJ,TruemanWH.Ataxonomis revisionMetarhiziumofbasedonaphylogenetic analysisofrDNAsequencedata.MycologyResearch, 2000.104:134~1508裴炎,冀志霞,杨星勇,等.绿僵菌分解昆虫外壳蛋白酶MAP一21的纯化与特性.微生物,2000,40 (3):306~31I代鹏等绿僵菌的研究进展9ColeSCJ,CharnleyAK,CooperRM.Purification andparticalcharacterizationofanoveltrypsiir一1ikecysteineproteasefromMtarhiziumanisopliae. FEMSMicrobio1ogyLetters,1993,113:189~19610StLegerRJ,BidochkaMJ,RobertsDW.Construc—tionofanimprovedmycoinsecticideoverexpress—ingatoxicprotease.ProceedingsoftheNational AcademyofSciencesoftheUnitedStatesofAmeri—ca,1996,93(13):6349~635411StLegerRJ,RobertsDW.Engineeringimproved mycoinsecticides.TrendsinBiotechno1ogy,1997,15(3):83~8512SmithsonSL,PatersonIC,BaileyAM,eta1. Cloningandcharacterizationofageneencodinga cuticle—degradingproteasefromtheinsect pathogenicfungusMetarhiziumanisopliae.Gene, 1995,166(1):161~16513StLegerRJ,JoshiL,RobertsD.AmbientpHisa majordeterminantintheexpressionofcuti—cle—degradingenzymesandbydrophobinby Metarhiziumanisopliae.AppliedandEnvironmental Microbiology,1998,64(2):709~71314WangChengshu,MiItonA,TariqM,eta1.Detec—tionandcharacterizationofPrlvirulentgene deficienciesintheinsectpathogenicMetarhizium anisopliae.FEMSMicrobiologyLetter,2002,213:25l~25515沈坤发,茅绍雄,林光明,等.主要虫生真菌的分子生物学研究概述.江西农业大学,2001,5(23): 140~14416李建庆,张永安,张星耀,等.昆虫病原真菌毒素的研究进展.林业科学研究,2003,16(2):233~23917李农昌,樊美珍,朱玮,等.绿僵菌毒素的提取方法.林业科技通讯,1989(1O):30~3118DrauzsioENRangel,GibertoULBraga,StephanD, eta1.Variationsin[IV—Btoleranceandgermina—tionspeedofMetarhiziumanisopliaeconidiapro—ducedoninsectsandartificalsubstrates.Jour—nalofInvertebratepathology,2004,115:207~21119BragaGUL,F1intSD,Mi1lerCD,eta1.Both solarUV AandUvBradiationconidialculturabi1i—tyanddelaygerminationintheentomopathogenic fugusMetarhiziumanisopliae.Photochemistryand Photobio1ogy,2001,74:734~73920王滨,樊美珍,李增智.真菌杀虫剂剂型的研究与应用.安徽农业大学,2003,30(2):206~20921InyangEN,McCartneyHA,OyejolaB,eta1.Ef—fectofformulation,applicationandrainonthe persistenceoftheentomogenousfungusMetarh~ium anisopliaeonoilseedrape.MycologyResearch,2000,104(6):653~66(上接第52页)27NinS,BenniciA,RoselliG,eta1.Agrobcteri—am—meadiatedtransformationofArtemisiaabsir~hium L.(wormwood)andproductionofsecondarymetabo一1ites.PlantCellReports,1997,16(1O):725~73028赵亚华,何平,高向阳.根癌农杆菌介导的mMT—I cDNA转化枸杞及其表达的研究.中国农业科学, 2000,33(2):92~9729费厚满,梅康凤,沈听,等.发根农杆菌对绞股蓝的转化及毛状根中皂甙的产生.植物,1993,353233(8):626~631盛世红,陈惠明.防风悬浮细胞的原生质再生植株培养.植物,1994,32(4):268~273谭红,龚革,李志东,等.利用原生质体诱变技术筛选脱落酸高产菌株.应用与环境生物,1998, 4(3):281~285周延清,苑保军,张根发,等.决明原生质体的分离与培养研究.华北农,1998,13(3):107~1l1孙如勇,安锡培.植物原生质体培养.北京:科学出版社,1991.108~116—77—。
离体培养条件下莱氏绿僵菌致病形态和非致病形态的转录组分析作者:刘守柱张希鹏郝轩卉任宪銮王世贤聂玉恒吴蕊来源:《山东农业科学》2023年第11期关键词:莱氏绿僵菌;转录组分析;致病形态;非致病形态;形态转变;离体培养莱氏绿僵菌[Metarhizium rileyi(Farlow)Kep-ler](Hypocreales:Clavicipitaceae)原名莱氏野村菌(Nomuraea rileyi),是一种重要的昆虫生防真菌,可在田间自然发生并对60多种鳞翅目害虫,尤其是夜蛾科害虫,如甜菜夜蛾(Spodopteraexigua)、棉铃虫(Helicoverpa armigera)、斜纹夜蛾(Spodoptera litura)及近年入侵我国的草地贪夜蛾(Spodoptera frugiperda)等有明显的致病作用,是一种具有广阔应用前景的生防真菌。
虽然莱氏绿僵菌在害虫生物防治领域有突出的优点,但较慢的致死速度是制约其大规模利用的瓶颈。
例如,用1x108个孢子/mL的莱氏绿僵菌孢子悬液处理斜纹夜蛾3龄幼虫,半致死时间(LT50)为5.5~6.6d;处理4、5龄幼虫时,LT50分别延长至9.8、11.0d。
莱氏绿僵菌的致病机制是一个复杂的生物学过程,可以简单归纳为孢子在昆虫体壁上的粘附、萌发、体壁穿透等体外侵染过程,以及侵入寄主血腔后的增殖、变形、分泌毒素、组织破坏等体内发育过程。
体外过程决定侵染成功率,体内过程决定致死速度。
研究表明,莱氏绿僵菌成功侵入昆虫血腔后,菌体在发育初期以小而短的酵母状形态存在,称为虫菌体(hyphal body,Hb),以芽殖的方式大量增殖,此时寄主正常进食、蜕皮和生长;当菌体数量达到一定密度阈值后,酵母状的Hb不再进行芽殖,而是向两端生长,形成细长的菌丝,并分泌毒素杀死寄主。
莱氏绿僵菌的体内发育过程表明该菌是一种二型性真菌(di-morphism fungus),虫菌体是非致病形态,不会影响寄主的生长,只有当其转变为菌丝后,才能表现出杀虫效果,是致病形态。