11 抗体及基因重排
- 格式:ppt
- 大小:11.34 MB
- 文档页数:67


第五章 免疫分子第一节 免疫球蛋白( Immunoglobulin, Ig )免疫球蛋白(immunoglobulin,Ig)包括成熟B 细胞的膜表面Ig(SmIg)和某个B 细胞克隆受抗原刺激、激活、分化、转化为浆细胞后分泌的Ig 。
由于Ig 具有与相应抗原发生特异结合的功能,故称为抗体(antibody,Ab)。
在人体内约有1×107个B 细胞克隆,各个B 细胞克隆的SmIg 分子的VL 、VH 结构都不一样,能识别差异极其微小的各种抗原决定簇,产生相应的Ig 分子。
分泌于血清中的Ig 分子结构极不均一,与抗原结合的特异性千差万别,Ig 自身表现的抗原性(血清型)也多种多样。
关于Ig 产生的遗传控制及多样性的起源,遗传学家们一直在进行研究,并存在分歧,提出的学说主要有胚系学说、体细胞突变学说和基因重排学说。
随着分子生物学技术的发展,基因重排学说不断被实验证实和完善,受到普遍赞同。
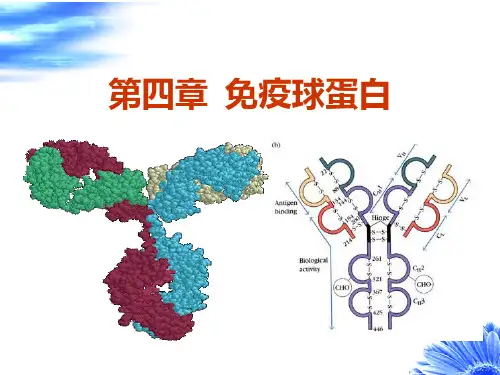
一、免疫球蛋白分子的基本结构Ig 分子的基本结构为,两条相同的轻链(light chain, L 链)在两条相同的重链(heavy chain, H 链)外侧,L 链和H 链之间,H 链与H 链之间,由二硫键连接成一个“Y ”字型。
每条L 链由210~230氨基酸组成,分子量约23ku ,每条H 链由420~446个氨基酸组成,分子量50~70ku 。
所有Ig 的L 链和H 链从氨基端起的100多个氨基酸的组成和顺序多变,称为可变区(variable region, V 区),L 链的V 区(VL)由108个氨基酸组成,H 链的V 区(VH)由107~130个氨基酸组成,VL 和VH 共同组成与相应抗原决定簇结合的部位。
在VL 和VH 中氨基酸组成的变化仅集中3个区域(占氨基酸总量的15%~20%),称为超变区(hypervariable region, HV)。
超变区是Ig 与抗原结定簇结合的互补决定区(complementarity-determining region,CDR)。

抗体多样性的遗传学原理摘要日本分子生物学家利根进川凭借抗体多样性遗传机制的发现,获得了1987年诺贝尔生理学或医学奖。
他主要运用限制酶酶切和重组DNA技术,通过演示一个DNA分子的突变和重组或重新排列证明了体细胞突变理论。
本文将就该成就的取得过程、实验原理和实验经过进行详细阐述。
关键词利根进川体细胞突变理论限制酶酶切重组DNA1、问题的产生二十世纪是生命科学迅猛发展的时代,免疫学是其中一个飞速发展的领域。
免疫学研究的基本问题之一是机体识别“自我”和“非我”。
生物体受到外源物质感染后,会启动体液免疫而产生某些特殊的蛋白质进行抵御,即抗体。
[1]由于可作为抗原刺激机体产生免疫应答的物种成千上万,理论而言可产生相应数量的抗体。
但一个物种只有数量有限的编码基因,因此20世纪70年代前,抗体多样性的产生机制一直是免疫学家争论不休的问题。
主要分歧为生殖系理论和体细胞突变理论。
生殖系理论认为,所有抗体都有专一基因负责,但该理论问题在于生物体内基因数目无法满足众多的抗体;体细胞突变理论认为,抗体基因可以发生突变和重组,该理论可以解释很少基因数能够产生大量微小差异的抗体,但缺乏有力的实验支持。
1971年,日本分子生物学家利根进川加入巴塞尔研究所工作,他相信凭借自己的分子生物学基础并应用当时的新技术——限制酶酶切和重组DNA等能够解决这个难题。
[2]2、抗体多样性产生遗传机制的发现2.1实验基础60年代中期抗体结构已被阐明,即一个免疫球蛋白分子包含两个相同的轻链(L 链)和两个相同的重链(H链),链链之间通过二硫键相连,两链各有一个恒定部分一个变异部分。
抗体分子氨基酸顺序分析表明,从氨基端起的大约110个氨基酸构成了可变区(V区),功能主要是抗原结合部位,抗体多样性的结构基础存在于此;而其余同源性较高的氨基酸顺序则称为稳定区(C区)。
可变区的氨基酸并非全部易变,变化最大的集中区域称之为高变区;而可变区其余部分的氨基酸变化很小,称之为骨架区。

收稿日期:20061028基金项目:国家自然科学基金重点项目资助(30230270)作者简介:张 红(1967),女,河南光山人,副教授,在读博士研究生,研究方向:分子免疫学。
通讯作者:张改平(1960),男,河南内黄人,研究员,博士生导师,博士,主要从事动物免疫学与生物技术研究。
鸡免疫球蛋白基因结构与抗体多样性产生的分子机制张 红1,2,3,刘钧珂3,张改平2*,王 丽2,席 俊2,宋海涛1,2(1.河南农业大学,河南郑州450002; 2.河南省动物免疫学重点实验室,河南郑州450002;3.河南教育学院,河南郑州450003)摘要:鸡具有独特的免疫球蛋白基因结构,抗体多样性产生机制有别于啮齿动物和灵长类动物。
种系基因重排产生了有限的抗体基因类型,抗体多样性的产生机制主要源于体细胞的基因转变,一种由胞嘧啶核苷脱氨酶(AID)启动的同源重组,V 基因上游的假基因取代了重排后有同源序列的可变区基因片段。
基因重排和基因转变过程受启动子、增强子和沉默子序列的调控,通过碱性螺旋-环-螺旋结构的转录因子E2A 进行调节。
关键词:鸡;免疫球蛋白基因;基因转变;抗体多样性;AID;E2A 中图分类号:S858.31 文献标识码:A 文章编号:10043268(2007)02010204鸡免疫系统具有十分独特的结构和抗体产生机制。
法氏囊是鸡重要的中枢免疫器官,在B 淋巴细胞分化增殖和抗体形成过程中发挥着至关重要的作用;抗体基因数量少且结构独特,抗体多样性机制与哺乳动物及人类有根本性差异。
所有这些特征激发了科研工作者对鸡免疫系统研究的兴趣,也使鸡成为抗体产生和B 细胞发育研究重要的模式生物物种。
文中阐述了有关鸡抗体基因结构及抗体多样性机制的最新研究进展。
1 免疫球蛋白功能性基因数量少,结构简单鸡抗体基因结构与啮齿动物、灵长类动物相比有很大差异。
小鼠抗体有3个独立的基因座编码,2个编码轻链( 和 )、1个编码重链。

抗体种类千千万,你知道原因么?人类生活在复杂多变的环境中,每时每刻都会接触到各种各样的微生物,受到一些类似抗原物质的侵扰,从而使机体致病。
为了抵御这些外来侵扰,使自身得以继续生存,人体必须形成几十万、几百万甚至更多种相应的特异性抗体以抵抗外界的抗原物质,才能免遭其害,保护自己。
我们会从抗体的产生及多样性进行其原因的阐述与分析。
1.抗原的呈递抗原呈递细胞(antigen presenting cell, APC)的抗原呈递作用是一个涉及抗原摄取、处理与呈递的复杂过程[1]。
最主要的抗原呈递分子是主要组织相容性复合物(majorhistocompatibility complex, MHC)[2]。
MHC分为两个大类:MHC-I和MHC-II,它们呈递的抗原蛋白来源不同,降解抗原的方式不同,结合肽段长度也不同,可以分别将抗原呈递给细胞毒性T细胞 (CD8 T cell) 和辅助型T细胞 (CD4 T cell)[3,4]。
结合现以MHC-II呈递外源性蛋白为例简述抗原提成过程。
外源性抗原经APC吞噬或吞饮作用,被摄入胞内形成吞噬体,后者与溶酶体融合形成吞噬溶酶体。
抗原在吞噬溶酶体内酸性环境中被蛋白水解酶降解为小分子多肽,其中具有免疫原性的称为抗原肽[3,5]。
内质网中合成的MHC-II类分子进入高尔基体后,由分泌小泡携带,通过与吞噬溶酶体融合,使抗原肽与小泡内MHC-Ⅱ类分子结合形成抗原肽-MHCⅡ类分子复合物。
所形成的复合物可能在高尔基复合体参与下被转运到细胞膜表面,被T细胞受体(TCR)识别并呈递给TH细胞[6]。
供TCR识别的先决条件是两种细胞的直接接触并相互作用。
这种细胞间的相互作用涉及APC与TH表面多种分子。
除了TCR特异性地同时识别多肽-MHC-Ⅱ分子的复合物外,某些粘附分子也参与抗原呈递过程。
活化的TH细胞可分泌各种细胞因子,用于B细胞、Tc细胞等的激活过程,产生可清除抗原的特异性抗体。

一、名词解释1.半抗原2.佐剂3.顺序决定簇4.Arthus反应5.H因子6.CD7.Ig8.ELISA9.AID10.MLR 11.Haplotype 12.Cytokine 13.mitogen 14.Isotype switch 15.Positive selection 16.免疫突触17.调节性T细胞18.抗原调变19.阴性选择20.MHC多态性21.AICD 22.GVHR23.KIR24.MHC 25.Affinity 26.Allergins 27.Autoimmunity 28.Clonal anergy 29.Molecular mimicry 30.Self tolerance 31.Chemokine 32.Epitope 33.SCID 34.GVHR 35.Hypersensitivity 36.MHC 37.Class switch 38.KIR39.CD40.Affinity四、问答题1.何谓免疫?简述免疫的功能?2.何谓抗原?半抗原及抗原决定簇?3.何谓人工自动免疫及人工被动免疫,并比较二者主要不同点?4.简述细胞因子的主要生物学作用?5.简述I型超敏反应产生的过程。
6.免疫球蛋白有哪些生物学功能?7.简述肿瘤的免疫逃逸机制。
8.简述T细胞介导的免疫应答过程。
9.抗体再次免疫应答有哪些特点?10.简述TCR基因重排的过程并试述其生物学意义。
11.试述注射青霉素后,机体可能产生超敏反应的类型、过程及其机制。
12.试比较同种异型器官移植过程中,抗原的直接识别与间接识别的机制有何不同。
13.图示补体活化的过程,并简要说明其活化的调控机制。
14.什么是T细胞活化的双信号系统?如何利用这一系统解释肿瘤细胞逃避免疫应答的现象?15.简述固有免疫应答的特点,以及机体固有免疫系统防御病原微生物入侵中过程中参与固有免疫细胞和分子的功能。
16.简述补体生物学作用。
17.比较人工自动免疫与人工被动免疫。

1、SD序列shine-Dalgarnoa sequence:存在于原核生物mRNA起始密码上游7-12个核苷酸的富含嘌呤的保守片段,能与16SrRNA3’端富含嘧啶的区域进行反向互补,所以将mRNA 的AUG起始密码子置于核糖体的适合位置以便起始翻译作用。
2、分子伴侣:他是细胞中一类能够识别并结合到不完全折叠或装备的蛋白质上帮助这些多肽链正确折叠、转运或防止它们聚集的蛋白质。
3、简并性:由一种以上的密码子编码同一个氨基酸的现象,对应于同一个氨基酸的密码子称为同义密码子。
4、遗传密码:指mRNA上每三个核苷酸翻译成多肽链上的一个氨基酸,这三个核苷酸就称为一个密码子。
5、终止密码子:不代表任何氨基酸,任何tRNA分子都不能识别的,但可以被终止因子或释放因子并引起新和成多肽链从核糖体上释放的密码子。
UGA UAA UAG6、密码子偏爱性:指不同种属的生物对简并密码具有不同的使用频率。
7、非编码区:不编码目标蛋白质的mRNA序列。
8、蛋白质糖基化:一种翻译后修饰,是氨基酸侧链共价键修饰中的一种,包括O-糖基化:侧链通过GalNAc连接在蛋白质的Ser或The的羟基处;N-糖基化:糖链连接在蛋白质的天冬酰胺的氨基侧链处。
9、反密码子:tRNA分子的反密码子环上的三联体核苷酸残基序列。
10、密码子变偶性:处于密码子3’端的碱基和与之互补反密码子5’端的碱基之间的配对有一定的自由度。
如I可以和密码子上3’端的U C A配对。
这种现象称为密码子的变偶性。
11、开放阅读框:指一组连续的含有三联密码子的能被阅读翻译成多肽链序列的DNA序列。
由起始密码子开始,到终止密码子结束。
12、多聚核糖体:核糖体在细胞内并不是单独执行功能的,而是由多个甚至几十个核糖体串联在一条mRNA分子上高效的进行多肽链的合成,这种具有特殊功能与形态结构的核糖体与mRNA的聚合体称为多聚核糖体。
13、移码突变frame shift mutation:由于单个碱基或者非三的整倍数的碱基的插入或缺失一起的从突变位点整个可读框的改变,从而产生完全不一样的氨基酸序列。



抗体的发现史及结构特征时间:2016-08-16 文章来源:抗体圈打印字号:大中小抗体(Antibody)这个词首次出现在保罗·埃尔利希1891年10月公布的《免疫力的试验性研究》这篇文章中,德语的抗体“Antikörper”出现在该文章的结论部分。
其中指出了“如果两种物质导致两种不同抗体的产生,那么这两种物质必然是不同的”。
然而这一术语并没有立即被接受,还有被建议使用的其它几个术语,如:免疫体(Immunkörper)、介体受体(Amboceptor)、介体(Zwischenkörper)、物质敏感体(substance sensibilisatrice)、连接体(copula)、德氏体(Desmon)、白细胞素(philocytase)、介体固定体(fixateur)以及免疫素(Immunisin)等。
抗体和抗毒素(Antitoxin)字面结构相似,概念则和免疫体(Immunkörper)类似。
根据E. Padlan公布的抗体结构图,朱利·安沃斯-安德烈*为斯克里普斯研究所佛罗里达分部创作了雕塑《西方天使》(2008年)塑像仿照列奥纳多·达·芬奇的维特鲁威人,将抗体放置到一个圆环的中间,寓意其对于人类的重要意义。
针对抗体的研究始于1890年,埃米尔·阿道夫·冯·贝林及北里柴三郎首次描述了抗体对白喉及破伤风痉挛毒素的抵抗作用。
他们两人将体液免疫理论往前推进了一步,提出了血清中存在一种可以与外来抗原相反应的某种介质的假设。
保罗·埃尔利希受到了他们的这一想法的启发,于1897年提出了抗体与抗原互动的侧链理论假说。
他假设道,在细胞的表面存在能和特定毒素发生一把钥匙对应一把锁类似的特异结合作用的感受器,而结合反应则会进一步导致相关抗体的生产。
其他研究人员在之后的研究中认为,抗体可以在血液中稳定独立存在。