拟南芥突变体种子库
- 格式:xlsx
- 大小:94.58 KB
- 文档页数:38

拟南芥发育异常突变体的基因克隆与表型分析摘要植物发育在器官、组织和细胞等不同层次上受到复杂的遗传互作网络调控。
本实验室的研究方向是利用正向遗传学方法探索植物非生物逆境响应相关发育性状形成的遗传基础和分子机制。
实验室在前期工作中,在激活标签突变体库中筛选获得一个拟南芥显性突变体abs3-1D(abnormal shoot3-1 dominant)。
abs3-1D突变体呈现株型小,叶柄短,叶片紧凑,颜色偏黄,开花明显提前,次生花序簇生,主花序茎矮化,短角果且种子的育性下降。
在黑暗诱导条件下,abs3-1D表现出叶片衰老加速的特征。
为进一步研究ABS3在植物发育过程的功能,以abs3-1D为遗传背景进行EMS(乙基磺酸甲酯, ethyl-methane sulphonate)诱变。
在诱变体库中筛选到能够逆转abs3-1D早花表型的突变体F08-69。
F08-69表现出晚花表型,并且F08-69single营养生长期延长,黑暗条件下叶片衰老速度减慢。
全基因组重测序鉴定出At3g10390基因+678bp处G变成A,导致甘氨酸(G)替换为丝氨酸(S)。
另外还发现叶色、叶形和表皮毛发育缺陷的突变,其中一株隐性突变体abt3-1(aberrantly branched trichome 3),其突变表型为植株小,叶形宽,叶色浓绿,尤其是其出现了高比例的多分支的表皮毛,并且根的生长存在缺陷。
abt3-1根的伸长速率仅为Col-0的三分之一。
通过对根的分生组织中皮层细胞数目和细胞长度的测定,表明abt3-1根的分生区变短。
该研究利用图位克隆技术发现abt3-1中At1g66510的第8个外显子的+33bp处G突变为A,提前引入终止密码子。
回复验证表明At1g66510(ABT3)的突变导致了abt3-1的表型。
ABT3基因编码一个目前功能未知的AAR2(a1- 2 repression 2)家族蛋白。
我们同时对ABT3蛋白的亚细胞定位进行了观察,发现ABT3蛋白定位于细胞质和细胞核中。

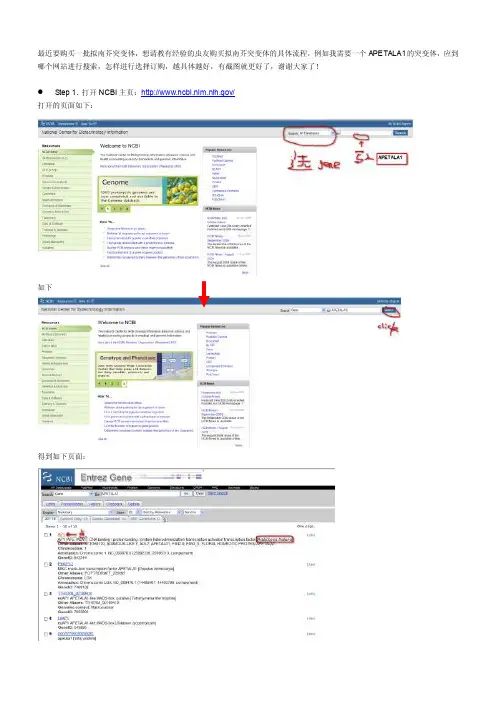
最近要购买一批拟南芥突变体,想请教有经验的虫友购买拟南芥突变体的具体流程,例如我需要一个APET ALA1的突变体,应到哪个网站进行搜索,怎样进行选择订购,越具体越好,有截图就更好了,谢谢大家了!Step 1. 打开NCBI主页:/打开的页面如下:如下得到如下页面:进一步获得该基因在NC BI里面的基因信息,到此我为什么要做这一步呢,主要是想获得该gene在拟南芥中的系统名,见下图:记住这个名称:AT1G69120这个就是APE TALA1(AP1)基因接下来开始查找 APETAL A1(AT1G69120)的突变体,拟南芥突变体库世界上有很多,公开的没有公开私用的都有,突变的方法也不尽相同,有DS的,T-DNA插入的,Tos17,EMS方法突变的等等。
但是,我们通常用美国SALK研究所的突变体库,这个突变体库比较权威,从这里可以找到几乎现有的所有拟南芥突变体,包括T-DNA插入,RIKENFST等等各种不同的突变类型,而且有详细的突变位点介绍和购买方法它的搜索界面一目了然,使用也很方便。
下面介绍SA LK突变体库的使用方法:Step 2:打开SALK主页:http://signa/点击 T-DNA Expres s 进入(红圈处点击),如下显示:显示如下,所有信息全在如下窗口中从上述窗口中可以获得很多不同gr oup制得的突变体,有SALKT-DNA,CSHL FST(冷泉港实验室的)等等,我个人建议使用SALK 的突变体,订购比较方便,听同学说好像一百美元一个,上图中,蓝色下划线的那两个,以SALK_冠名的那个,两个显示的是不同的插入位置,和T-DNA插入方向(看在图中的位置和箭头方向)点击其中一个进入信息页,比如点击SA LK_056708,得到如下页面:我们主要是从ABRC 订购,点击进入页面,填写要求的相关信息,万事大吉。


《拟南芥耐铯突变体atbe1-5的筛选及其机理的研究》篇一一、引言随着人类对环境资源的需求日益增长,重金属污染问题日益严重。
其中,铯(Cs)作为一类典型的重金属元素,因其高度的可移动性和长寿命对生态环境构成威胁。
对于植物的抗性研究,尤其是对于模式植物拟南芥耐铯突变体的筛选和其耐铯机理的研究,成为了近年来的研究热点。
本文就拟南芥耐铯突变体atbe1-5的筛选及其机理进行深入探讨,旨在为重金属污染环境下的植物耐性机制研究提供新的视角。
二、研究方法本部分主要介绍了在实验室环境中,如何筛选拟南芥耐铯突变体atbe1-5,以及其筛选的具体方法和实验过程。
(一)突变体的筛选通过一系列的突变体筛选方法,包括物理诱变、化学诱变和分子遗传操作等手段,得到了一个拟南芥耐铯突变体atbe1-5。
详细介绍了铯离子浓度梯度的设定,突变的测定方式,如基因型和表现型的鉴定等。
(二)分子生物学方法在实验过程中,通过使用分子生物学方法如基因克隆、基因序列分析等手段,深入解析了该突变体的遗传特性和耐铯机制。
三、实验结果(一)atbe1-5突变体的筛选结果经过一系列的筛选和鉴定,成功筛选出拟南芥耐铯突变体atbe1-5。
该突变体在较高浓度的铯离子环境下仍能正常生长和发育。
(二)atbe1-5突变体的遗传特性分析通过基因克隆和序列分析,发现atbe1-5突变体与野生型相比在某个关键基因上存在单碱基或单核苷酸差异,这些差异导致突变体的表达方式和表型特性发生变化。
这种改变赋予了突变体更强的耐铯能力。
(三)atbe1-5突变体的耐铯机理研究研究发现,atbe1-5突变体在铯离子胁迫下能更好地激活抗逆机制。
在转录水平和翻译水平上表现出不同的调控方式,同时涉及一些重要酶和基因的活跃性增强,帮助植株抵御高浓度的铯离子带来的压力。
此外,该突变体在铯离子的吸收和转运方面也表现出独特的特点。
四、讨论与结论本研究通过系统的筛选和分析拟南芥耐铯突变体atbe1-5的遗传特性和耐铯机理,深入探讨了其可能的生物学意义和应用价值。

拟南芥突变体的鉴定摘要在真核细胞的细胞核中,基因组DNA缠绕在组蛋白上,形成核小体的结构。
核心组蛋白的末端会发生一系列翻译后水平上的共价修饰,主要包括甲基化、乙酰化、磷酸化、泛素化和ADP-核糖基化等。
这些共价修饰在染色质的结构与功能及基因的转录调控方面起着重要的作用。
组蛋白的甲基化发生在赖氨酸和精氨酸位点,这些位点的甲基化参与异染色质形成,X染色体失活和基因转录的激活与沉默等许多重要的生物学功能。
早先认为组蛋白上的甲基化是不可逆的。
然而近年来在哺乳动物和酵母中相继发现了许多组蛋白去甲基化酶。
其中最大的一个基因家族是包含有JMJC结构域的组蛋白去甲基化酶家族。
在植物中还没有报道有组蛋白去甲基化酶。
我们通过基因组注释以及序列比对从拟南芥中鉴定出21个编码包含有JMJC结构域蛋白的基因。
并从SALK突变体库中筛选鉴定了相应的T-DNA插入突变体。
利用分子标记,我们筛选出了37个纯合T-DNA插入株系。
给我们以后分析这些基因在植物生长发育中所起的作用打下了坚实的基础。
关键词组蛋白密码, 甲基化,去甲基化,T-DNA插入突变体1 背景介绍1.1 “组蛋白密码”理论与表观遗传学在真核细胞的细胞核中,基因组DNA与组蛋白和非组蛋白相互结合,构成遗传信息的物质载体——染色质。
染色质由其基本单位核小体重复连接而成,每个核小体包含一个组蛋白八聚体(H2A/H2B-H3/H4-H3/H4-H2A/H2B)和围绕其上的两圈超螺旋DNA(146KB),核小体之间通过不同长度的超螺旋DNA和H1组蛋白前后相接。
组蛋白八聚体的成员作为核小体的基本组成元件,在所有真核生物中高度保守。
尽管核心组蛋白均具有一个三螺旋的结构严整的组蛋白折叠区,其两个末端却没有形成固定结构。
然而,这些末端却会发生一系列翻译后水平上的共价修饰,主要包括甲基化、乙酰化、磷酸化、泛素化和ADP-核糖基化等。
这些共价修饰在染色质的结构与功能及基因的转录调控方面起着重要的作用(1,2)。


《拟南芥耐铯突变体atbe1-5的筛选及其机理的研究》篇一一、引言随着现代工业的快速发展,重金属污染问题日益严重,特别是铯等重金属的污染已经成为了一个严重的环境问题。
为了更好地解决这一难题,研究人员一直致力于寻找耐铯能力强且能在铯污染土壤中正常生长的植物种类和机制。
近年来,在多种植物中筛选出的耐铯突变体为我们提供了新的研究思路。
本文以拟南芥耐铯突变体atbe1-5为研究对象,对其筛选方法及耐铯机理进行深入研究。
二、材料与方法1. 实验材料本实验采用拟南芥野生型及耐铯突变体atbe1-5为实验材料。
2. 实验方法(1)突变体的筛选:通过化学诱变法获得拟南芥突变体库,利用铯处理法筛选出耐铯能力强的突变体。
(2)生理生化分析:对筛选出的耐铯突变体进行生长曲线、生物量、叶绿素含量等生理指标的测定。
同时,对铯处理后植物的根部形态和铯的分布进行分析。
(3)基因表达分析:采用RNA-Seq等基因组学技术对atbe1-5突变体在铯处理前后的基因表达水平进行分析,筛选出与耐铯性相关的基因。
(4)转录因子和调控网络分析:对筛选出的基因进行转录因子预测和调控网络分析,探讨其耐铯机理。
三、结果与分析1. 突变体的筛选结果通过化学诱变法和铯处理法,我们成功筛选出了一批耐铯能力强的拟南芥突变体,其中atbe1-5的耐铯能力尤为突出。
与野生型相比,atbe1-5在铯处理下生长更为旺盛,生物量也更大。
2. 生理生化分析结果生理生化分析表明,atbe1-5突变体在铯处理后,其叶绿素含量、光合作用等生理指标均表现出较强的抗逆性。
此外,其根部形态也发生了明显变化,根毛增多、根系更发达,有利于植物在重金属污染土壤中的生长。
3. 基因表达分析结果基因表达分析结果显示,atbe1-5突变体在铯处理后,与野生型相比,一些与重金属抗性、细胞代谢和能量代谢等相关的基因表达水平发生了显著变化。
这些基因的差异表达可能参与了atbe1-5突变体的耐铯过程。

拟南芥T-DNA插入突变体的鉴定摘要:我们用CTAB法提取拟南芥的T-DNA插入突变体的DNA,然后用三引物法进行PCR和琼脂糖凝胶电泳来判断其为突变纯合体还是突变杂合体。
通过这次实验,我们掌握了如何来判断纯和突变和杂合突变。
关键字:拟南芥 T-DNA插入突变突变体的鉴定前言:拟南芥拟南芥是十字花科的植物,它是植物遗传学、发育生物学和分子生物学的模式植物,其具有以下这些特点:①植株形态个体小,高度只有30cm左右;②生长周期快,从播种到收获种子一般只需8周左右;③种子多,每株可产生数千粒种子;④形态特征简单,生命力强,用普通培养基就可作人工培养;⑤遗传转化简单,转化效率高;⑥基因组小,只有5对染色体,125MB;⑦在2000年,拟南芥成为第一个基因组被完整测序的植物。
突变体突变体在植物基因分离及遗传学研究的最重要材料,通过自然突变或者人工诱变同源重组、基因沉默以及插入突变等方法都可以用来构建突变体,人工诱变是指利用物理因素(X射线,Y射线,紫外线,激光等)或化学诱变(如亚硝酸,硫酸二乙酯)来处理生物,使生物发生基因突变,这种方法可提高突变率,创造人类需要的变异类型。
目前,人工诱变拟南芥常用的方法有EMS诱变、T-DNA 插入突变、激活标签等。
T-DNA插入突变Ti质粒是土壤农杆菌的天然质粒,该质粒上有一段特殊的DNA区段,当农杆菌侵染植物细胞时,该DNA区段能自发转移,插入植物染色体DNA中,Ti质粒上的这一段能转移的DNA被叫做T-DNA。
人们根据这一现象,将Ti质粒进行改造,将感兴趣的基因放进T-DNA区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化。
T-DNA插入到植物染色体上的什么位置,是随机的。
如果T-DNA插入某个功能基因的内部,特别是插入到外显子区,将造成基因功能的丧失。
所以利用农杆菌Ti质粒转化植物细胞,是获得植物突变体的一种重要方法。
T—DNA插入突变最大的用处是构建突变体库,在此基础上构建侧翼序列库;目前在拟南芥中已经建立了接近饱和的T—DNA插入突变体库,该突变体库包含超过225 000个独立的T—DNA插入株系,在预测的29 454个基因中有21 700个基因发生了插入突变[6]。

拟南芥突变体的相关研究遗传学摘要:本文列举了利用正向遗传法对拟南芥突变体的筛选、遗传群体的初步遗传群体及初步遗传图谱的构建和基因的图位克隆、遗传分析及相关基因的功能分析的流程,为拟南芥的研究提供更明确更清晰的思路。
关键词:拟南芥突变体;筛选;图位克隆;功能分析1 拟南芥突变体的筛选拟南芥是十字花科拟南芥属植物,近年来拟南芥以其个体小、生长周期短以及基因组小等特点而成为分子遗传学研究的模式植物。
拟南芥的另一优点是很容易被诱变,目前已从拟南芥中分离得到了几千种突变体,这些突变体的获得为揭示植物生长发育规律起了非常重要的作用。
拟南芥突变体的筛选已成为许多重要理论问题得以解决的前提,而筛选方法是突变体筛选成败的关键。
这里拟南芥耐低钾突变体的筛选为例,介绍一种简单、灵敏、通用的拟南芥突变体的筛选方法。
1.1植物材料诱变以拟南芥为材料,诱变方法如下:称取250mg(约5000粒)野生型种子置于50ml烧杯内并加入25ml 重蒸水,搅拌30 分钟;在4℃,下放置12小时后,把种子转移到盛30ml100mmol/L 磷酸缓冲液(PH6.5)的100ml三角瓶中,加入0.2%(V / V)的甲基磺酸乙酯(EMS),封口后放在水浴(25℃)振荡器上振荡12h。
然后用50ml 蒸馏水漂洗种子4 次,每次15min。
将漂洗好的种子置于4℃下春化3天后种植。
1.2诱变植株培养将经EMS诱变处理后的拟南芥种子(M1)播种于1/4Hoagland 营养液浸透的混有蛭石的营养土中,然后覆膜保湿。
18-22℃、光照强度120umol/m2s-1、光周期16h/8h条件下培养,待种子成熟后分行采收种子。
1.3 突变体的筛选拟南芥种子用0.5 %(v/ v)次氯酸钠加0.1%(v/ v)Tri-tonX-100表面消毒10 分钟,再用无菌水冲洗3遍。
接种前种子与0.4%(w/ v)低熔点琼脂糖混和,然后用吸管将种子吸出,成行地涂于MS 培养基上;将培养皿置于4℃冰箱春化48小时,之后转入光照培养,培养皿垂直放置。

拟南芥属植物分子遗传学和突变体筛选研究方法拟南芥(Arabidopsis thaliana)是目前广泛应用于分子遗传学和突变体筛选的模式植物。
它具有小型体积、短生命周期、易于培养和遗传变异等优点,使其成为研究植物基因功能的理想模型。
下面将介绍拟南芥属植物的分子遗传学和突变体筛选研究方法。
一、拟南芥分子遗传学研究方法2. 基因组学方法:包括全基因组测序(Whole Genome Sequencing, WGS)、基因芯片(Microarray)和下一代测序(Next Generation Sequencing, NGS)等技术,用于分析和比较拟南芥基因组的序列、结构和功能。
3.双杂交法:通过构建酵母杂交系统,研究和鉴定拟南芥基因间的物理和功能相互作用关系,进而揭示拟南芥基因调控网络和信号转导途径。
4. RNA干扰(RNA interference, RNAi)技术:利用沉默诱导的RNA (siRNA)或者镰刀状RNA(hairpin RNA)介导靶向基因的沉默,从而研究和验证拟南芥基因的功能。
二、拟南芥突变体筛选方法1. EMS化学诱变:使用化学诱变剂EMS(Ethyl methanesulfonate),处理拟南芥种子,让其发生突变,形成突变种子库。
进一步筛选和鉴定突变体,识别和研究拟南芥基因的突变功能。
2. 插入序列突变法:通过插入转座子(Transposon)或者T-DNA转座子,将外源序列插入拟南芥基因组,产生随机或特异性的基因突变,进行筛选和分析。
3.含有T-DNA插入的突变体库:使用含有T-DNA插入的突变体库,通过筛选和分离带有T-DNA插入的个体,鉴定和研究拟南芥基因的功能和表达调控。
4.突变体数据库查询:拟南芥基因突变体数据库中收集了大量已经鉴定和命名的突变体信息,可以通过数据库查询,寻找和鉴定具有特定表型的突变体。
最近要购买一批拟南芥突变体,想请教有经验的虫友购买拟南芥突变体的具体流程,例如我需要一个APETALA1的突变体,应到哪个网站进行搜索,怎样进行选择订购,越具体越好,有截图就更好了,谢谢大家了!
Step 1. 打开NCBI主页:/
打开的页面如下:
如下
得到如下页面:
进一步获得该基因在NCBI里面的基因信息,到此我为什么要做这一步呢,主要是想获得该gene在拟南芥中的系统名,见下图:
记住这个名称:AT1G69120这个就是APETALA1(AP1)基因
接下来开始查找APETALA1(AT1G69120)的突变体,拟南芥突变体库世界上有很多,公开的没有公开私用的都有,突变的方法也不尽相同,有DS的,T-DNA插入的,Tos17,EMS方法突变的等等。
但是,我们通常用美国SALK研究所的突变体库,这个突变体库比较权威,从这里可以找到几乎现有的所有拟南芥突变体,包括T-DNA插入,RIKEN FST等等各种不同的突变类型,而且有详细的突变位点介绍和购买方法它的搜索界面一目了然,使用也很方便。
下面介绍SALK突变体库的使用方法:
Step 2:打开SALK主页:/
点击T-DNA Express 进入(红圈处点击),如下显示:
显示如下,所有信息全在如下窗口中
从上述窗口中可以获得很多不同group制得的突变体,有SALK T-DNA,CSHL FST(冷泉港实验室的)等等,我个人建议使用SALK的突变体,订购比较方便,听同学说好像一百美元一个,上图中,蓝色下划线的那两个,以SALK_冠名的那个,两个显示的是不同的插入位置,和T-DNA插入方向(看在图中的位置和箭头方向)
点击其中一个进入信息页,比如点击SALK_056708,得到如下页面:
祝实验顺利!。
拟南芥突变体的相关研究遗传学摘要:本文列举了利用正向遗传法对拟南芥突变体的筛选、遗传群体的初步遗传群体及初步遗传图谱的构建和基因的图位克隆、遗传分析及相关基因的功能分析的流程,为拟南芥的研究提供更明确更清晰的思路。
关键词:拟南芥突变体;筛选;图位克隆;功能分析1 拟南芥突变体的筛选拟南芥是十字花科拟南芥属植物,近年来拟南芥以其个体小、生长周期短以及基因组小等特点而成为分子遗传学研究的模式植物。
拟南芥的另一优点是很容易被诱变,目前已从拟南芥中分离得到了几千种突变体,这些突变体的获得为揭示植物生长发育规律起了非常重要的作用。
拟南芥突变体的筛选已成为许多重要理论问题得以解决的前提,而筛选方法是突变体筛选成败的关键。
这里拟南芥耐低钾突变体的筛选为例,介绍一种简单、灵敏、通用的拟南芥突变体的筛选方法。
1.1植物材料诱变以拟南芥为材料,诱变方法如下:称取250mg(约5000粒)野生型种子置于50ml烧杯内并加入25ml 重蒸水,搅拌30 分钟;在4℃,下放置12小时后,把种子转移到盛30ml100mmol/L 磷酸缓冲液(PH6.5)的100ml三角瓶中,加入0.2%(V / V)的甲基磺酸乙酯(EMS),封口后放在水浴(25℃)振荡器上振荡12h。
然后用50ml 蒸馏水漂洗种子4 次,每次15min。
将漂洗好的种子置于4℃下春化3天后种植。
1.2诱变植株培养将经EMS诱变处理后的拟南芥种子(M1)播种于1/4Hoagland 营养液浸透的混有蛭石的营养土中,然后覆膜保湿。
18-22℃、光照强度120umol/m2s-1、光周期16h/8h条件下培养,待种子成熟后分行采收种子。
1.3 突变体的筛选拟南芥种子用0.5 %(v/ v)次氯酸钠加0.1%(v/ v)Tri-tonX-100表面消毒10 分钟,再用无菌水冲洗3遍。
接种前种子与0.4%(w/ v)低熔点琼脂糖混和,然后用吸管将种子吸出,成行地涂于MS 培养基上;将培养皿置于4℃冰箱春化48小时,之后转入光照培养,培养皿垂直放置。
拟南芥基因突变体筛选和分类的研究拟南芥(Arabidopsis thaliana)是植物研究中的常用材料,因其具有许多优点:生长快、体形小、基因组测序完成、适应广泛等等。
而生物学研究中最为基础的就是基因研究,因此引发人们对拟南芥基因的探索和研究。
本文将会介绍拟南芥基因的突变体筛选和分类的研究。
一、拟南芥的基因突变体筛选在拟南芥中,基因突变体是非常重要的,由于拟南芥基因组的测序已经完成,因此,科研人员可以利用现代高通量筛选技术,来快速建立大规模的拟南芥基因突变体资源库。
1.1 传统筛选法最常用的传统筛选方法是化学诱变、X射线、γ射线和紫外线辐射等方法,其基本原理是:通过人为或其他因素对植物种子或幼苗进行处理,使其基因发生变异,最终产生突变体。
其中,化学诱变是使用化学物质诱导植物基因发生变异,这种方法有较大优势,因为可以在不依赖实验室设备的情况下大规模筛选。
但是,化学诱变方法可能会引起伪突变和失败的突变率较高。
1.2 高通量筛选法随着科学技术的发展,高通量筛选方法得到了广泛应用,尤其是基于基因编辑技术的筛选方法。
目前流行的高通量筛选法有:T-DNA插入、CRISPR/Cas9、RNA干扰等。
其中,T-DNA插入是在拟南芥基因组中随机插入T-DNA,每个T-DNA都能够导致拟南芥基因表达的改变。
因此,T-DNA插入是一种高效、简单、易于筛选和操作的方法,并且在拟南芥研究中应用广泛。
二、拟南芥基因突变体分类拟南芥基因突变体分类是指将筛选得到的突变体按照突变部位和性质分为不同的类型,以方便基因功能的研究和应用。
2.1 快速分离突变位点的方法首先,需要快速、准确地确定突变体的位点,以此进行后续的基因功能分析,现在,基因突变体的位点鉴定方法已经得到了较大的改进,常见的方法包括PCR-RFLP、dCAPS和PCR-sequencing等。
其中,PCR-RFLP是一种快速、简便的检测方法,基本原理是:通过PCR扩增突变体和野生型基因区段,然后将PCR产物限制性酶切,从而分辨突变体和野生型基因。
拟南芥化学变异体库在基因功能研究中的应用一、引言拟南芥(Arabidopsis thaliana)作为模式植物,被广泛应用于植物基因功能研究中。
随着基因组测序技术的发展,拟南芥的基因组信息已经得到了完整的描绘,但基因序列信息本身并不能完整反映出基因的生物学功能。
化学变异体库是一种通过化学处理改变拟南芥遗传信息的方法,可以帮助研究者更深入地了解基因生物学功能。
本文将从化学变异体库的产生、特点和应用角度来阐述其在基因功能研究中的重要作用。
二、化学变异体库的产生化学变异体库产生的第一步是对拟南芥种子进行化学处理。
不同的化学剂会引起不同的变异,例如:硝基苯(2,4-DNP)、桉叶酸(EMS)、硝基硫酸(NaN3)等。
这些化学剂能使拟南芥发生遗传变异,形成各种各样的变异体。
然后将这些变异体收集起来进行命名和分类,形成化学变异体库。
三、化学变异体库的特点化学变异体库的特点是具有遗传多样性和突变类型的单独性。
不同的化学处理方式会引起不同的突变,从而得到多样的遗传变异体。
每个变异体都是独立的,具有单独的突变类型和特征。
四、化学变异体库的应用化学变异体库已经被广泛应用于拟南芥基因功能研究中。
1. 突变体筛选化学变异体库中的变异体可以用于筛选出具有特定性状或表现型的突变体。
通过对化学变异体库进行筛选,可以获得一批具有特定性状或表现型的拟南芥突变体。
这些突变体可以进一步用于基因功能的研究。
2. 基因互补化学变异体库也可以用于基因互补研究。
比如,我们可以通过化学变异体库得到一个特定基因的不同突变体,然后将这些突变体与同一基因的其它突变体进行互补研究。
通过这样的研究,可以揭示基因的不同区域的作用和功能。
3. 信号转导研究化学变异体库的应用还可以有效地揭示信号转导通路。
比如,我们可以通过化学变异体库筛选出一个与某个特定信号转导通路有关的突变体,并将这个突变体作为研究对象。
然后通过对该突变体的研究,揭示与该信号转导通路相关的基因和基因功能。