第二章 免疫分子 6 MHC参与的抗原递呈
- 格式:pdf
- 大小:280.30 KB
- 文档页数:11

简述细胞免疫的过程
细胞免疫是一种免疫响应过程,主要涉及T淋巴细胞和细胞因子的参与。
其过程可以分为以下几个步骤:
1. 抗原递呈:免疫细胞(如抗原呈递细胞、树突状细胞等)摄取并处理外源抗原,将其片段展示在细胞表面的主要组织相容性复合物(MHC)上。
2. T细胞激活:这些展示抗原的MHC分子与特异性受体(T 细胞受体,TCR)上的T细胞结合,从而通过信号传导通路激活T细胞。
3. T细胞增殖和分化:活化的T细胞通过增殖和分化产生大量的效应细胞。
其中,CD4+ T细胞分化为不同亚群(如辅助性T细胞1型、2型、17型、调节性T细胞等),而CD8+ T细胞则分化为细胞毒性T细胞。
4. 效应细胞杀伤:细胞毒性T细胞识别并杀伤感染细胞或异常细胞(如肿瘤细胞)。
它们通过释放细胞毒素(如穿孔素、颗粒酶等)或通过Fas/FasL通路诱导细胞凋亡来发挥杀伤作用。
5. 细胞因子产生:辅助性T细胞分泌细胞因子(如干扰素γ、白细胞介素等),促进炎症反应、调节免疫细胞活性和协调免疫应答。
6. 调节与终止:调节性T细胞通过分泌调节性细胞因子,抑
制其他免疫细胞的活性,维持免疫的平衡。
一旦病原体被清除,免疫应答将被终止,避免过度炎症反应。
细胞免疫通过调控免疫细胞的活性和杀伤作用,起到防御感染和清除异常细胞的作用。
这个过程中多个细胞和分子参与,相互协同发挥作用,维护机体的免疫平衡和健康。
第二章MHC分子与移植第一节MHC 分子一、概述主要组织相容性抗原系统(major histocompatibility system, MHC),是指在哺乳动物体内受遗传控制的,代表个体特异性的主要组织抗原系统。
其编码基因是由紧密连锁的一组基因群组成,每个基因都有40多个等位基因。
MHC编码的产物表达在多种细胞表面,称为MHC 分子或MHC抗原。
MHC分子有两种类型:MHC-I类和MHC-II类。
起初,人们认为MHC 分子在T细胞对异体移植的组织进行排斥反应中起作用,即当机体从其他个体接受移植物后,如果受体的MHC与供体不同,就会对移植物产生强烈的排斥反应;后来证明,MHC除参与器官移植排斥外,还参与免疫应答和免疫识别的调控。
小鼠MHC命名为H-2(histocompatibility antigen-2),位于第17对染色体。
人类的MHC命名为HLA(Human Leukocyte Antigen),位于第6对染色体短臂。
多态性(Polymorphism):同一种属不同个体间在基因或蛋白水平上的变异。
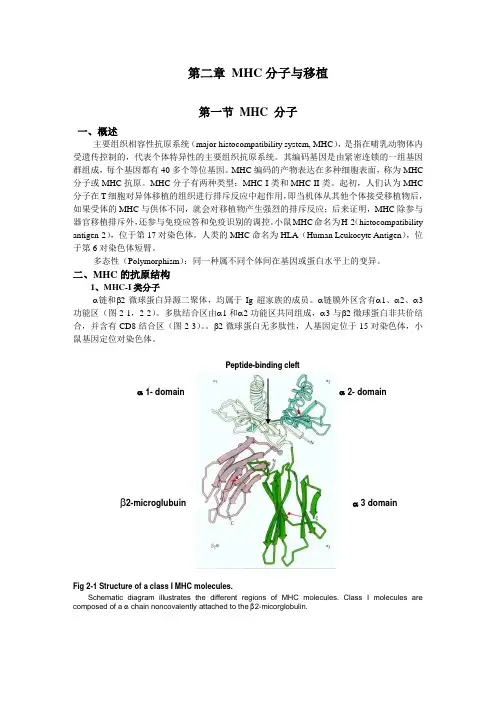
二、MHC的抗原结构1、MHC-I类分子α链和β2微球蛋白异源二聚体,均属于Ig超家族的成员。
α链膜外区含有α1、α2、α3功能区(图2-1,2-2)。
多肽结合区由α1和α2功能区共同组成,α3与β2微球蛋白非共价结合,并含有CD8结合区(图2-3)。
β2微球蛋白无多肽性,人基因定位于15对染色体,小鼠基因定位对染色体。
P e p t i d e-b i n d i n g c l e f tα 1- domain α 2- domain β2-microglubuin α 3 domainFig 2-1 Structure of a class I MHC molecules.Schematic diagram illustrates the different regions of MHC molecules. Class I molecules are composed of a α chain noncovalently attached to the β2-micorglobulin.Fig2-2 T h e α1 and α2 domains as viewed from topSchematic diagram Showing the peptide-binding cleft consisting of a base of antiparallel β strands and sides of α helices. This cleft in Class I molecules can accommodate peptides containing 8-10 residues.HLA- I APCFig 2-3 The α3 segment of the α chain folds into an Ig domain that serves as the binding site for CD 8.2、MHC-II 类分子:αβ异源二聚体,α链膜外区含有α1和α2功能区;β链膜外区含有β1和β2功能区。


抗原的加工呈递和主要组织相容性复合体学习要求:掌握:主要组织相容性复合体的概念人类经典MHC基因编码的分子结构、组织分布及其功能特征熟悉:MHC基因结构及其特性MHC分子与抗原肽的相互作用了解:免疫功能相关基因HLA在医学中的应用和意义第一节MHC(对应书上第三节)一、简介1. MHC的概念及MHC限制性(掌握)主要组织相容性复合体(Major histocompatibility complex):是一组决定移植组织是否相容,与免疫应答密切相关,紧密连锁的基因群。
MHC限制性(MHC restriction):2. MHC的发现(了解)二、MHC基因结构及功能1. MHC 基因结构和分布(掌握人类经典MHC基因编码的分子结构、组织分布)I类基因区:编码MHC I类分子的基因:II类基因区:编码MHC II类分子的基因:2. MHC 的多基因性和多态性(熟悉)•单体型haplotype•连锁不平衡linkage disequilibrium3. MHC分子及与抗原肽的结合(掌握经典HLA分子的功能特征)HLA分子的功能:(参考PPT P67-68)1. 作为抗原提呈分子参与适应性免疫应答2. 作为调节分子参与固有免疫应答4. 非经典MHC 分子(了解)CD1分子呈递脂质抗原三、HLA与疾病的关系(了解,参考书P77)(熟悉几个关联基因)✧强直性脊柱炎AS——HLA-B27阳性✧乳糜泻CD——HLA-DQ第二节抗原呈递细胞(对应书上第一节)✧抗原呈递细胞概念:✧抗原提呈细胞的分类(专职性和非专职性)✧抗原呈递细胞的特性:•活化初始化和效应性T细胞•呈递抗原给T细胞(第一信号),并提供第二信号•PAMP分子增强APC的抗原呈递能力•APC 和T 细胞的双向作用一、细胞(掌握APC类型)(一)树突状细胞1.DC的类型(cDC,pDC)2.经典DC的成熟(了解)未成熟DC(抗原的摄取加工);成熟DC(抗原呈递)(参考书P64,表5-1)3.DC作为专职APC的重要特性:4.DC能够活化初始T细胞的原因:(高海峰老师在复习讲座中提及,可帮助理解)初始T细胞活化条件:起始APC必须表达足够多的MHC分子、黏附分子和协同刺激分子,还必须有足够的机会与初始T细胞相遇。


抗原递呈的名词解释抗原递呈是指免疫系统中一种重要的生物学过程,它在机体抵御各种病原微生物和异常细胞的免疫应答中起到至关重要的作用。
本文将对抗原递呈的概念、机制和意义进行解释和探讨。
一、抗原递呈的概念抗原递呈是指细胞表面的抗原被免疫细胞摄取、分解,并通过特定分子的结合和展示,呈现给免疫系统中的T细胞,以引发针对该抗原的免疫反应。
这个过程是免疫系统有效应对外来病原体的关键环节。
二、抗原递呈的机制抗原递呈的机制主要涉及抗原提呈细胞(APC)和T淋巴细胞之间的相互作用。
APC通常是树突状细胞(DC)、B淋巴细胞和巨噬细胞等,它们具有递呈抗原的能力。
在抗原递呈过程中,APC首先摄取外源性或内源性抗原,并将其分解成小片段。
然后,这些片段与主要组织相容性复合体(MHC)分子结合,形成MHC抗原复合物。
这样,抗原被呈现在APC表面的MHC分子上。
一旦抗原呈现在MHC分子上,它就可以与T细胞上的特异性受体,即T细胞受体(TCR)结合。
这个结合会触发一系列的信号转导事件,最终导致T细胞的活化。
被活化的T细胞可以释放细胞因子,调节和增强免疫应答。
这种抗原递呈和T细胞激活的机制保证了机体对病原体和异常细胞的有效识别和清除。
三、抗原递呈的意义抗原递呈对于维持机体免疫状态和应对病原微生物非常重要。
通过抗原递呈的过程,机体可以识别和区分自身抗原和非自身抗原,从而避免自身免疫反应。
同时,抗原递呈还能够引发免疫耐受,抑制过度免疫反应和自身免疫疾病的发生。
抗原递呈还对疫苗设计和免疫治疗具有重要意义。
通过合理选择和设计递呈抗原的载体、调节抗原递呈细胞的活性和改变抗原递呈方式,可以增强或抑制机体特定免疫反应的程度和效力。
这为疫苗的研发和免疫疗法的应用提供了理论依据和技术手段。
最后,抗原递呈还是免疫学研究的重要领域。
深入研究抗原递呈的机制和调控方式,可以探索机体免疫系统的复杂性和多样性,为疾病发生、发展和治疗提供新的认识和思路。
综上所述,抗原递呈作为免疫系统中的一种重要过程,具有广泛的意义和应用前景。

免疫系统中的抗原递呈与识别免疫系统对于维护我们的健康和抵御病原体的攻击起着重要的作用。
在这个系统中,抗原递呈与识别是其中的重要环节之一。
它可以使机体识别出敌人,发动免疫反应,从而防止疾病的发生。
在本文中,我们将探讨免疫系统中的抗原递呈与识别这一重要的过程的机制和作用。
1. 抗原递呈与识别的基本原理在免疫系统中,抗原是指能够诱导机体免疫反应的物质。
它可以是细胞表面的分子,如病毒和细菌表面的蛋白质或糖类,也可以是自身分子失去了正常结构,其形态发生了改变,被免疫系统错误地视为敌人的物质,如肿瘤细胞和转移细胞。
抗原递呈与识别的基本原理是:在抗原递呈细胞(APC)中,抗原与主要组织相容性复合物(MHC)分子结合,形成MHC-抗原复合物,并能与T细胞表面的受体结合,从而激活T细胞的免疫应答。
MHC分子是一类主要存在于APC表面的分子,它的主要作用是展示抗原分子,使其与T细胞表面的受体结合,从而激活T细胞。
MHC分子主要分为两类:MHC-I分子和MHC-II分子。
MHC-I分子主要存在于所有核细胞和部分细胞表面,可以呈现内源性抗原,如突变的自身细胞或病毒感染细胞的抗原分子;MHC-II分子主要存在于APC表面,可以呈现外源性抗原,如细菌、病毒等抗原分子。
2. 抗原递呈与识别的过程在抗原递呈与识别的过程中,抗原递呈细胞首先通过吞噬、胞吐、自发捕获等方式获取外源性或内源性抗原分子。
然后,这些抗原分子会与MHC分子结合,形成复合物。
对于MHC-I分子来说,抗原分子一般先在细胞内被加工、分解成小片段,并被转运到内质网中与MHC-I分子结合形成复合物,再被转运到细胞表面。
对于MHC-II分子来说,抗原先在吞噬囊泡内被加工、切割成小片段,然后与MHC-II分子结合,形成复合物,再经过胆囊泡转运到APC表面。
在这个过程中,影响抗原递呈与识别的因素非常复杂,包括抗原分子自身的结构、抗原递呈细胞的类型和状态、MHC分子的类型和亚型、以及T细胞的表面受体等。

MHC与抗原处理和递呈抗原处理和呈递是适应性免疫的基石,没有T细胞的帮助,B细胞不能产生高亲和力抗体。
而CD4+T细胞通过细胞表面的抗原特异性受体提供这种帮助,这些受体识别与肽段结合的主要组织相容性复合体(MHC)分子,从而激活T 细胞,然后T细胞执行效应器功能,如细胞毒性、和产生细胞因子。
同样,负责清除病毒感染的细胞毒性CD8+T细胞也依赖于对肽-MHC复合物的识别来发挥作用。
负责抗原呈递的两大类糖蛋白包括MHC I类和II类分子,它们分别向CD8+T细胞和CD4+T细胞呈递抗原肽。
我们对于抗原呈递的理解,很重要的是:它为适应性免疫系统提供了一种调查宿主蛋白质及其修饰的方法,任何可能表明存在病毒和细菌等入侵者或导致恶性肿瘤突变的变化都可以被检测到。
而在没有感染源或癌细胞的情况下,抗原呈递为免疫系统诱导自身耐受提供了参考。
先天性免疫反应和适应性免疫反应在功能上相互依存,它们以不同的方式保持对自我的非反应状态。
先天免疫反应使用模式受体识别微生物成分的简约解决方案,而适应性免疫反应的的解决方案则基于对高度多样化、躯体生成的抗原特异性受体的严格选择。
因此,抗原呈递与建立功能正常、自我耐受的T细胞库有着不可磨灭的联系。
MHC分子的结构编码人类主要组织相容性复合物的基因位于第6号染色体短臂。
1999年,HLA(人类白细胞抗原,即人类的MHC)基因序列分析工作完成并揭示,它是迄今为止人体中最复杂、多态性最丰富的遗传系统,拥有极大数量等位基因,赋予种群对各种病原体显示最合适免疫应答的巨大潜力,以适应多变的内外环境。
经典的HLA分子包括HLAⅠ类分子(HLA-A、B、C)和HLAⅡ类分子(HLA-DR、DP、DQ),均为糖蛋白。
HLA Ⅰ类分子表达在绝大多数有核细胞表面,但不同的组织和细胞的表达水平不同。
HLAⅡ类分子表达范围较窄,主要分布在单核细胞/单核巨噬细胞、DC细胞、B细胞以及激活的T 细胞和胸腺上皮细胞表面;以上均为结构性表达。

第一章绪论1名词解释:免疫:从狭义上讲,免疫是人体对病原体(病菌、病毒)和有害物质的抵抗力。
从广义上讲,免疫是机体识别“自身”与“非己”抗原,对自身抗原形成免疫耐受,对非己抗原产生排斥反应的一种生理功能。
2.机体免疫系统识别和清除突变细胞的功能称为(C)A.免疫耐受B.免疫自稳C.免疫监视D.免疫防御E.免疫调节3.最早提出克隆选择学说的科学家是(D)A.JennerB.BorderC.PorterD.BurnetE.Pasteur4.最早用牛痘预防天花的科学家是(D)A.BurnetB.BorderC.PorterD.JennerE.Pasteur5.免疫指(ABCDE)A.机体排除病原微生物的功能。
B.机体清除损伤和衰老细胞的功能。
C.机体识别和排除抗原性异物的功能。
D.机体抗感染的防御功能E.机体识别和清除自身突变细胞的功能。
第二章免疫器官1.简述淋巴细胞再循环的概念及其意义淋巴细胞再循环:定居在外周免疫器官的淋巴细胞,由输出淋巴管进入胸导管,经上腔静脉进入血液循环,在毛细血管后微静脉处穿越血管内皮细胞表面(HEV),重新分布于全身淋巴器官和组织。
淋巴细胞在血液、淋巴液、淋巴器官和组织间周而复始循环的过程即为淋巴细胞再循环。
淋巴细胞再循环的意义:(1)淋巴细胞在淋巴组织和器官中分布更为合理(2)淋巴组织不断从循环池中补充新的淋巴细胞有助于增强整个机体的免疫功能(3)有利于淋巴细胞与抗原和抗原递呈细胞接触(4)有利于动员效应淋巴细胞迁移至炎症部位;(5)定居在外周免疫器官的记忆性细胞也参与再循环,其接触相应抗原后进入淋巴组织,并迅速发生活化、增殖和分化,产生再次免疫应答。
2.骨髓的主要功能包括(ABDE)A.各类血细胞和免疫细胞发生的场所B.B细胞分化成熟的场所C.T细胞分化成熟的场所D.再次体液免疫应答发生的场所E.既是中枢免疫器官,又是外周免疫器官3.T细胞分化成熟的场所是(B)A.骨髓B.胸腺C.腔上囊D.淋巴结E.脾4.B细胞分化成熟的场所是(A)A.骨髓B.胸腺C.腔上囊D.淋巴结E.脾第三章抗原1、TD2抗原:一类能刺激机体免疫系统使之产生特异性免疫应答,并能与相应的应答产物在体内外发生特异性结合的物质。


抗原递呈细胞(APC)与MHC 在免疫应答过程中,除T细胞和B细胞起核⼼作⽤外,单核巨噬细胞和树突状细胞也参加发挥作⽤,主要是处理和递呈抗原,故称抗原递呈细胞(antigen presenting cells,APC),亦可称为辅佐细胞(accessory cells,A cells)或A细胞。
APC能通过吞噬或胞饮作⽤摄取和处理抗原,并将经过处理得到的含有抗原决定簇的多肽⽚段与MHCⅡ类分⼦结合,然后表达于细胞表⾯递呈给CD4+TH细胞。
具有抗原递呈作⽤的细胞有单核巨噬细胞,树突状细胞和B细胞三类。
虽然有核细胞均表达MHCⅠ类分⼦,也能将胞浆内的蛋⽩抗原处理降解为多肽⽚段,与Ⅰ类分⼦结合后表达在细胞表⾯递呈给CD8+TC细胞,有递呈抗原作⽤,但习惯上不将这些细胞归类于专职APC,⽽称其为靶细胞。
⼀、单核吞噬细胞 ⾎液中的单核细胞(monocytes)和组织中的巨噬细胞(macrophages,Mφ)统称为单核吞噬细胞系统(mononuclear phagocyte system)。
单核吞噬细胞有较强的粘附玻璃或塑料表⾯的特性,⽽淋巴细胞⽆此能⼒,可利⽤该特点分离和获取单核吞噬细胞。
单核细胞和巨噬细胞表⾯有多种受体。
与免疫功能有关的重要受体有IgG的Fc受体(CD64)和补体C3b受体,以及某些淋巴因⼦受体。
巨噬细胞表⾯有较多的MHCⅠ类和Ⅱ类分⼦,与抗原递呈有关。
单核吞噬细胞在免疫应答中的功能如下: 1.吞噬和杀伤作⽤ 巨噬细胞可吞噬较⼤的病原微⽣物和衰⽼损伤细胞。
已被抗体(IgG)和补体(C3b)结合的细菌等抗原异物,更易被巨噬细胞吞噬,称为抗体和补体的调理作⽤。
被巨噬细胞吞噬的细菌等异物在吞噬体内被杀伤或消化降解。
也可通过Fc受体与被IgG抗体结合的靶细胞发⽣结合,发挥ADCC作⽤杀伤靶细胞。
IFNγ可激活巨噬细胞,增强其杀伤细胞内寄⽣菌和肿瘤细胞的活性。
但有时巨噬细胞对伤寒杆菌和结核杆菌等杀伤⼒有限,特别在未经上述细菌免疫的机体内,这些细菌可能存活并在巨噬细胞内增殖,造成感染的扩散或迁延。
免疫学mhc名词解释MHC(主要组织相容性复合物)是一种重要的分子机制,它在免疫系统中起着关键的作用。
MHC分子是一种膜结合的蛋白质,存在于几乎所有的脊椎动物细胞表面。
它们的主要功能是在免疫系统中呈递抗原给免疫细胞,以及确定自身细胞和非自身细胞之间的差异。
MHC分子有两种主要类型:MHC-I和MHC-II。
MHC-I分子主要存在于几乎所有核细胞表面,包括细胞核形成细胞(如肌肉细胞和神经元)。
MHC-I分子的主要功能是呈递细胞内产生的抗原给CD8+ T细胞。
这种呈递能力使得免疫系统能够识别被感染的细胞,进而引发针对感染源的免疫反应。
MHC-II分子主要存在于特定免疫细胞,如抗原递呈细胞(如树突状细胞、B细胞和巨噬细胞)。
MHC-II分子的主要功能是呈递外源性抗原给CD4+ T细胞。
这种呈递能力使得CD4+ T细胞能够识别外来抗原,进而引发免疫应答。
MHC分子有多个亚型,其具体功能和特性在不同物种和个体间可能不同。
不同的MHC分子会结合不同的肽段,并呈递给免疫细胞进行识别。
这种选择性的呈递能力使得免疫系统能够识别和应对多样性的病原体。
MHC分子在移植医学中扮演重要角色。
由于MHC分子是高度多态的,移植过程中的MHC配型需要尽可能的匹配。
如果移植接受者和供体的MHC之间存在较大的差异,有可能导致宿主对移植物产生免疫应答,从而使移植物被宿主排斥。
总之,MHC是一组重要的分子机制,在免疫系统中起着关键的作用。
它们通过呈递抗原给免疫细胞,使免疫系统能够识别和应对感染和外来抗原。
同时,MHC也在移植医学中至关重要,其匹配度的差异可能导致移植物的排斥。
第二章免疫分子6MHC参与的抗原递呈第六节 MHC参与的抗原递呈(MHC Mediated Antigen Presentation) 近年来,对抗原识别的研究一直是免疫学的热点,特别是对抗原加工和递呈(antigen processing and antigen presentation)的研究。
目前已初步揭开了免疫应答中抗原信息产生和传导的秘密,即抗原的加工和递呈受控于主要组织相容性复合体(major histocompatibity complex,MHC)系统。
尽管关于MHC分子的很多细节尚有待深入研究,但目前已明确MHC 产物行使着将抗原递呈给T细胞的重要作用。
抗原的加工和递呈有两条不同的途径:一是内源性抗原途径,抗原在内质网(ER)和高尔基器内加工,并与MHC-Ⅰ类分子结合后,被递呈到细胞表面,加工后的抗原能被CD8+T细胞识别;二是外源性抗原途径,在内吞体(endosome) 抗原被加工降解,形成的抗原片段与MHC-Ⅱ类分子结合后,转运到细胞表面,它可被CD4+T细胞识别。
MHC分子的抗原递呈功能是免疫应答和免疫调节的关键,MHC 分子是免疫细胞间信息沟通、相互协作的基础。
从分子水平研究MHC 结构和功能,对揭示抗原递呈的复杂机理有重要意义。
一、MHC参与抗原递呈的分子结构MHC分子在内质网产生并装配,与加工后的抗原肽特异性地结合、保护、运输、递呈抗原肽。
MHC分子主要分为两类,即MHC-Ⅰ类分子和MHC-Ⅱ类分子,这两类MHC分子的结构及在抗原递呈中的作用存在一定的差异。
应用X射线衍射晶体分析技术确定了两类MHC分子的结构特征,即在MHC分子的抗原结合部有一个深的肽结合槽。
它由两条α螺旋和8股β折叠片组成。
两条α螺旋位于槽的上部形成两个侧面,8股β折叠位于槽的下部形成底面,这样就形成一个深槽状的三维空间结构。
MHC-Ⅰ类分子能结合8~9个氨基酸残基的短肽,MHC-Ⅱ类分子能结合13~17个残基的短肽。
第六节 MHC参与的抗原递呈(MHC Mediated Antigen Presentation)近年来,对抗原识别的研究一直是免疫学的热点,特别是对抗原加工和递呈(antigen processing and antigen presentation)的研究。
目前已初步揭开了免疫应答中抗原信息产生和传导的秘密,即抗原的加工和递呈受控于主要组织相容性复合体(major histocompatibity complex,MHC)系统。
尽管关于MHC分子的很多细节尚有待深入研究,但目前已明确MHC 产物行使着将抗原递呈给T细胞的重要作用。
抗原的加工和递呈有两条不同的途径:一是内源性抗原途径,抗原在内质网(ER)和高尔基器内加工,并与MHC-Ⅰ类分子结合后,被递呈到细胞表面,加工后的抗原能被CD8+T细胞识别;二是外源性抗原途径,在内吞体(endosome) 抗原被加工降解,形成的抗原片段与MHC-Ⅱ类分子结合后,转运到细胞表面,它可被CD4+T细胞识别。
MHC分子的抗原递呈功能是免疫应答和免疫调节的关键,因为MHC分子是免疫细胞间信息沟通、相互协作的基础。
从分子水平研究MHC结构和功能,对揭示抗原递呈的复杂机理有重要意义。
一、MHC参与抗原递呈的分子结构MHC分子在内质网产生并装配,特异性地与加工后的抗原肽结合、保护、运输、递呈抗原肽。
MHC分子主要分为两类,即MHC-Ⅰ类分子和MHC-Ⅱ类分子,这两类MHC分子的结构及在抗原递呈中的作用存在一定的差异。
应用X射线衍射晶体分析技术确定了两类MHC分子的结构特征,即在MHC分子的抗原结合部有一个深的肽结合槽。
它由两条α螺旋和8股β折叠片组成。
两条α螺旋位于槽的上部形成两个侧面,8股β折叠位于槽的下部形成底面,这样就形成一个深槽状的三维空间结构。
MHC-Ⅰ类分子能结合8~9个氨基酸残基的短肽,MHC-Ⅱ类分子能结合13~17个残基的短肽。
为什么会有这样的差异? Rudensky等证实,MHC-Ⅰ类分子的肽结合槽两个末端是关闭的,其中一端有一个恒定的酪氨酸闭合,另一端有一个恒定的盐桥。
而MHC-Ⅱ类分子的一个末端是关闭的,即有一个盐桥,另一端既没有潜在的盐桥,也没有恒定的酪氨酸,而有许多侧链。
也有报道MHC-Ⅱ类分子的肽结合槽两端均是打开的。
因此,较长的具有抗原性的短肽片段可以优先被MHC-Ⅱ类分子递呈。
一些研究表明未折叠肽能够被MHC-Ⅱ类分子递呈而不需要进一步加工,因为这些线带样的蛋白质分子可能适合于两端开放的MHC-Ⅱ类分子的结合槽。
事实说明,MHC分子与抗原多肽形成复合物的特殊构型是T细胞激活的必要条件。
20世纪90年代以来,在已知编码与抗原递呈有关的MHCⅠ类及Ⅱ类分子的基因聚集排列的基础上,人们又在MHC基因区域发现了两种新的参与内源性抗原加工和提呈的基因,一种是低分子多肽复合物-2及低分子多肽复合物-7(low molecular mass polypeptide,LMP-2、LMP-7)基因,与内源性抗原(如细胞内的病毒抗原、肿瘤抗原)在细胞浆内降解为抗原片段有关;另一种为抗原加工相关载体(transporter associated with antigen processing,TAP)基因,其编码产物将降解好的不带信号肽的抗原片段以非分泌途径输送到内质网。
抗原片段在内质网中与新合成的MHC-Ⅰ类分子重链的肽结合槽结合,折叠形成一定的空间构型,随后β2微球蛋白结合上来,形成稳定的肽-MHC分子复合体,经高尔基体以囊泡形式转运到细胞表面,被细胞毒T细胞识别,引发随后一连串的免疫反应。
由于MHC-Ⅰ类基因、TAP基因、LMP-2及LMP-7基因位置靠近,转录活性同时受γ干扰素的诱导,能够协调三者基因产物的量,有效地递呈抗原,故三者构成的系统被比喻成真核生物MHC-Ⅰ类分子抗原加工和递呈的“操纵子”。
TAP及LMP-2、LMP-7基因处于MHC-Ⅱ基因区域,LMP-2及LMP-7分别位于TAP-1及TAP-2基因的左侧,与TAP基因的位置紧邻。
目前发现的TAP 基因分为2种:TAP-1及TAP-2。
它们各自因生物种类不同而有许多别称,1991年国际HLA命名会议将它们统一定名为TAP-1及TAP-2。
来自不同种属的TAP基因产物氨基酸顺序呈现高度的同源性,如小鼠TAP-1基因产物的氨基酸顺序与大鼠及人对应产物的顺序同源性为72%~89%。
二、T淋巴细胞对MHC限制性抗原识别的现象在Th细胞和CTL识别抗原的认识上有一个关键性的进展,就是自身MHC限制现象的发现。
MHC限制性要求APC必须表达能被T细胞识别的自身MHC分子,T细胞才能识别表达在该APC上的外源蛋白质抗原并产生应答。
T细胞识别的自身MHC分子是指在胸腺中的前体T细胞成熟过程中遭遇到的MHC分子。
“自身MHC”并非指T细胞本身所表达的MHC分子,而是指表达在APC细胞或靶细胞表面的MHC分子。
通常,因为T细胞与APC细胞发生在同一个体,是同基因的,APC细胞上的所有MHC分子都被同个体的所有T细胞视为自身MHC。
在实验系统中,T细胞可对某一特定APC所表达的抗原发生应答。
这两种细胞类型至少有一部分的共同基因。
尤其是当它们来源于共有一个或多个MHC等位基因的个体或近交系时,APC表达的MHC分子,在T细胞的成熟过程中已经遭遇并将之视为自身MHC。
将来自某一近交系动物的T细胞与不同近交系的APC细胞混合,并检查T细胞的应答,可能发现自身MHC限制性现象。
以下三组实验证实了Th 细胞和CTL抗原识别的MHC限制性。
①经抗原免疫的近交系豚鼠的T细胞,只有当同系巨噬细胞存在时,才能在试管内对抗原发生应答并分裂增殖。
这种增殖的T细胞大多是Th细胞。
进一步分析近交系和同基因系鼠显示,为了将抗原呈递给Th细胞,APC必须表达能被T细胞识别为自身的MHC-Ⅱ类分子。
这类实验表明,Th细胞对抗原的识别是MHC-Ⅱ类限制性的。
②利用过继转移技术,在近交鼠体内所做的实验和体外抗体产生的研究中显示,只有当B细胞表达能被T细胞识别的自身的MHC-Ⅱ类分子时,辅助T细胞与B细胞才能协同作用并对蛋白质抗原产生抗体应答,由此,辅助T细胞是MHC-Ⅱ类限制性的结论得到进一步支持。
③对于MHC限制性,最有力的证明是来自CTL对鼠和人的病毒感染靶细胞的特异性杀伤作用的分析。
当病毒感染靶细胞表达的MHC-Ⅰ类分子被T细胞识别为自身MHC时,该靶细胞才被杀伤。
这就证实了CTL识别病毒抗原是MHC-Ⅰ类限制性的。
这些实验证明,参与T细胞抗原识别的MHC基因产物必须表达在APC 上或CTL介导溶细胞作用的靶细胞上。
现有一些实验证实确实如此,并且必要的MHC分子不必表达在T细胞自身表面。
①一种A型MHC限制的抗原特异性单克隆T细胞,对抗原的应答需要表达A型MHC产物的APC存在。
在源于(A×B)F1个体的抗原特异性T细胞中,有些是MHC A限制性的,另一些是MHC B限制性,有很少一部分细胞亚群可能被(A×B)F1鼠上表达的MHC杂交分子所限制。
关键在于, MHC A限制性的T细胞即使是取自(A×B)F1鼠,也对表达MHC A的APC呈递的抗原发生应答。
②MHC A-限制性T 细胞对抗原的应答可被A型MHC分子特异性抗体所阻断,CD8+T细胞的杀伤活性可被针对MHC-Ⅰ类分子的抗体所抑制,而抗MHC-Ⅱ类分子的抗体可抑制CD4+T细胞的抗原激活;这种抑制仅发生在抗体结合至APC上时,而结合在T细胞上则不受抑制。
③不表达限制性MHC分子的APC不能刺激抗原特异性T细胞。
例如,在上述例子中,MHC-限制性T细胞对MHC A阴性的APC不发生应答;如果将MHC A基因转移至APC细胞,使之成为MHC A阳性细胞,它就会获得刺激T细胞的能力。
将抗原递呈给抗原特异性CD4+T细胞需要MHC-Ⅱ类分子的表达。
该实验中,来自H-2K鼠(表达MHC-Ⅰ类分子,而不表达MHC-Ⅱ类分子)的鼠成纤维细胞株3T3,不能将作为抗原的细胞色素C呈递给对该抗原特异的I-E K限制性T细胞杂交系。
把编码分子I-E Kα和β链的功能基因转移至3T3细胞中,它就能向该T细胞系呈递抗原。
这些实验引出一个基本的概念,抗原特异性的辅助性T淋巴细胞和溶细胞性T淋巴细胞同时识别表达在APC或靶细胞表面的两种结构--外来抗原和自身MHC分子。
上述实验提示Th细胞和CTL分别受到MHC-Ⅱ类分子和MHC-Ⅰ类分子的限制。
实际上,T细胞的MHC-Ⅰ类或MHC-Ⅱ类限制性更多地与CD8或CD4分子的表达有关,而非细胞的作用能力。
这样,所有CD4+T细胞为MHC-Ⅱ类分子所限制,事实上CD4分子本身就结合MHC-Ⅱ类分子。
大多数CD4+T细胞是辅助性细胞,尽管在人类和小鼠中已经证实了一种CD4+CTL(亦为MHC-Ⅱ类限制)。
相反,所有CD8+T细胞都是MHC-Ⅰ类限制性,CD8分子结合MHC-Ⅰ类分子。
大多数CD8+T细胞为CTL,也可能具有某些辅助细胞产生细胞因子的作用。
三、MHC-Ⅰ类分子参与的抗原递呈(一) MHC-Ⅰ类分子递呈抗原的过程 MHC-Ⅰ类分子递呈的抗原是细胞内源性多肽,它们往往来自胞浆或核蛋白,如细胞自身蛋白,病毒蛋白等,均是细胞内合成的蛋白质产物。
它们在胞浆中受蛋白水解酶的作用,裂解成为小的片段或多肽前体。
多肽与MHC-Ⅰ类分子的组装发生在内质网内。
存在于胞浆里的多肽必须经过专门通道进入内质网,这一通道就是与 MHC相关的多肽转运的基因产物-TAP(transporter associated with antigen processing)。
TAP属于ATP结合蛋白转运基因家族的成员,它形成多个跨膜结构,在不同的细胞均有表达,包括原核细胞和哺乳类动物细胞。
TAP为二分子的异二聚体,TAP1和TAP2存在于内质网膜上的跨膜结构,每个亚单位贯穿细胞膜6次,形成一个“孔”样结构。
它的基因座位与LMP-2,LMP-7相邻,均位于MHC-Ⅱ类基因群内,推测这是生物进化的结果。
至于多肽如何通过TAP进入内质网,目前仍不清楚,可能是多肽片段和TAP“孔”状结构胞浆端的结合位点相互作用,依靠TAP中ATP 的水解引起TAP二聚体的构型改变,导致TAP对应的多肽结合位点暴露,随后在内质网腔内多肽的释放。
目前已肯定TAP转运器提供来自胞浆的抗原多肽并传递给MHC-Ⅰ类分子,而且研究表明,TAP2所介导的多肽转移是一特异事件,因为实验发现多肽的结构构成和TAP2结构多态性的变化将阻断这一转运作用,从而,人们认为多肽羧基端残基决定的TAP选择性转运和MHC-Ⅰ类分子特异的多肽结合关键基序是受控于MHC的。