基因敲除小鼠的构建图文
- 格式:pptx
- 大小:956.62 KB
- 文档页数:15

基因敲除小鼠(Knockoutmice)制备技术方法基因敲除小鼠,人们使用复杂的方法使小鼠体内的某一个基因不表达,从而使小鼠呈现这个基因缺失的状态,可用于研究这个基因的功能。
但如果某个基因功能特别重要,这个基因缺失可能具有胚胎致死性,那我们就无法得到这种基因敲除的小鼠了,于是人们发明了条件性基因敲除技术。
这一技术可以实现在特定的时间、特定的细胞或组织内使某个基因沉默。
方法是首先在目的基因(就是打算敲除的那个基因)的两侧分别插入一段名为LoxP的DNA序列(LoxP序列是一段34bp的DNA序列,两端的13个碱基为回文序列,中间的8个碱基决定LoxP的方向。
然后我们需要用到一种带有Cre酶的转基因小鼠了。
Cre重组酶于1981年从P1噬菌体中发现,属于λ Int酶超基因家族。
Cre重组酶基因编码区序列全长1029bp(EMBL数据库登录号X03453),编码38kDa蛋白质。
是一种位点特异性重组酶,能介导两个LoxP位点(序列)之间的特异性重组,使LoxP位点间的基因序列被删除或重组。
LoxP(locus of X-over P1)序列来源于P1噬菌体,是有两个13bp反向重复序列和中间间隔的8bp序列共同组成,8bp的间隔序列同时也确定了LoxP的方向。
Cre 在催化DNA链交换过程中与DNA共价结合,13bp的反向重复序列是Cre酶的结合域。
其序列如下:5' - ATAACTTCGTATA - ATGTATGC - TATACGAAGTTAT - 3'3' - TATTGAAGCATAT - TACATACG - ATATGCTTCAATA - 5'Cre-LoxP系统的特性Cre重组酶介导两个LoxP位点间的重组是一个动态、可逆的过程,可以分成三种情况:1、如果两个LoxP位点位于一条DNA链上,且方向相同,Cre重组酶能有效切除两个LoxP位点间的序列;2、如果两个LoxP位点位于一条DNA链上,但方向相反,Cre重组酶能导致两个LoxP位点间的序列倒位;3、如果两个LoxP位点分别位于两条不同的DNA链或染色体上,Cre酶能介导两条DNA链的交换或染色体易位。
![建立基因敲除小鼠颈动脉粥样硬化模型[新版]](https://uimg.taocdn.com/7e07c982866fb84ae55c8d02.webp)
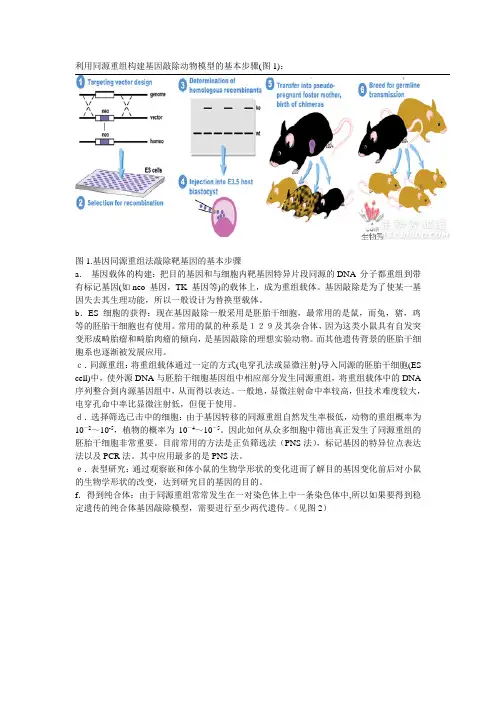
利用同源重组构建基因敲除动物模型的基本步骤(图1):图1.基因同源重组法敲除靶基因的基本步骤a.基因载体的构建:把目的基因和与细胞内靶基因特异片段同源的DNA 分子都重组到带有标记基因(如neo 基因,TK 基因等)的载体上,成为重组载体。
基因敲除是为了使某一基因失去其生理功能,所以一般设计为替换型载体。
b.ES 细胞的获得:现在基因敲除一般采用是胚胎干细胞,最常用的是鼠,而兔,猪,鸡等的胚胎干细胞也有使用。
常用的鼠的种系是129及其杂合体,因为这类小鼠具有自发突变形成畸胎瘤和畸胎肉瘤的倾向,是基因敲除的理想实验动物。
而其他遗传背景的胚胎干细胞系也逐渐被发展应用。
c.同源重组:将重组载体通过一定的方式(电穿孔法或显微注射)导入同源的胚胎干细胞(ES cell)中,使外源DNA与胚胎干细胞基因组中相应部分发生同源重组,将重组载体中的DNA 序列整合到内源基因组中,从而得以表达。
一般地,显微注射命中率较高,但技术难度较大,电穿孔命中率比显微注射低,但便于使用。
d.选择筛选已击中的细胞:由于基因转移的同源重组自然发生率极低,动物的重组概率为10-2~10-5,植物的概率为10-4~10-5。
因此如何从众多细胞中筛出真正发生了同源重组的胚胎干细胞非常重要。
目前常用的方法是正负筛选法(PNS法),标记基因的特异位点表达法以及PCR法。
其中应用最多的是PNS法。
e.表型研究:通过观察嵌和体小鼠的生物学形状的变化进而了解目的基因变化前后对小鼠的生物学形状的改变,达到研究目的基因的目的。
f.得到纯合体:由于同源重组常常发生在一对染色体上中一条染色体中,所以如果要得到稳定遗传的纯合体基因敲除模型,需要进行至少两代遗传。
(见图2)图2. 由嵌合体得到基因敲除的纯合体小鼠。


Dkk3基因敲除小鼠模型构建技术原理有研究通过同源重组的方式构建了Dkk3基因缺陷小鼠模型。
等位基因缺陷纯合子的小鼠可正常生存、可育的、甲状腺正常,但表现出亢奋的精神状态,自然杀伤细胞比例显著下降,IgM、血红蛋白和红细胞压积水平显著升高[4]。
在小鼠中,Dkk3缺陷会促进动脉粥样硬化的发生。
研究人员将Dkk3缺陷小鼠和ApoE缺陷型小鼠杂交,用以产生Dkk3和ApoE双基因缺陷型小鼠。
接着,主动脉的油红染色切片的分析显示(图2C),DKK3+/+/ApoE‒/‒小鼠的病变程度比DKK3‒/‒/ApoE‒/‒小鼠要小(图2A和2B)。
免疫荧光分析显示,与DKK3‒/‒/ApoE‒/‒小鼠相比,DKK3+/+/ApoE‒/‒小鼠(图2A和2D)病灶中的αSMA染色显著增加。
此外,对CD68标记物的染色显示DKK3+/+/ApoE‒/‒小鼠中损伤的巨噬细胞的数量减少(图2A和2D)。
这些数据共同表明,DKK3可能对ApoE‒/‒小鼠的动脉粥样硬化具有保护作用[5]。
图. 在20周龄时对接受正常饲料喂养的小鼠进行安乐死,并采集其心脏和主动脉。
切开主动脉根部,固定主动脉用油红染料进行染色并测量和量化主动脉窦及表面的病变区域。
切片分别用抗α平滑肌动蛋白(SM α-actin)和CD68抗体进行免疫染色,之后在显微镜下对阳性细胞进行计数。
A、使用油红染色主动脉窦具有代表性的切片图片,α-平滑肌肌动蛋白抗体标识出平滑肌细胞,CD68抗体标识出巨噬细胞;B、在表面着色病变区域中量化αSMA和CD68阳性细胞数量。
C、病变区占总表面积的百分比。
D、总细胞中αSMA和CD68阳性细胞所占百分比。
研究人员发现,Dkk3缺陷型小鼠在缺血时梗死面积增大。
在大脑中动脉闭塞手术处理之后,Dkk3−/−组小鼠脑部梗死体积都明显增大(图3A,B),这表明内源性DKK3对大脑缺血的损伤具有一定的抵抗作用。
还能观察到,在缺血病灶周围的大脑皮质内侧和下部检测到Dkk3免疫反应性增加,说明该区域可能包含缺血半暗带。
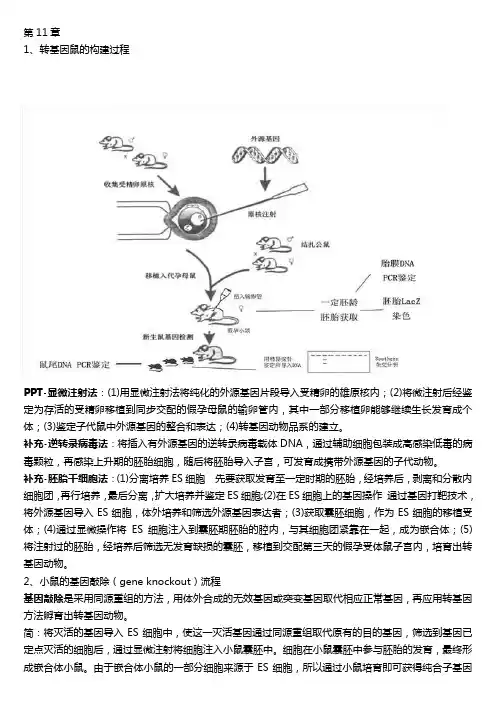
第11章1、转基因鼠的构建过程PPT-显微注射法:(1)用显微注射法将纯化的外源基因片段导入受精卵的雄原核内;(2)将微注射后经鉴定为存活的受精卵移植到同步交配的假孕母鼠的输卵管内,其中一部分移植卵能够继续生长发育成个体;(3)鉴定子代鼠中外源基因的整合和表达;(4)转基因动物品系的建立。
补充-逆转录病毒法:将插入有外源基因的逆转录病毒载体DNA,通过辅助细胞包装成高感染低毒的病毒颗粒,再感染上升期的胚胎细胞,随后将胚胎导入子宫,可发育成携带外源基因的子代动物。
补充-胚胎干细胞法:(1)分离培养ES细胞先要获取发育至一定时期的胚胎,经培养后,剥离和分散内细胞团,再行培养,最后分离,扩大培养并鉴定ES细胞;(2)在ES细胞上的基因操作通过基因打靶技术,将外源基因导入ES细胞,体外培养和筛选外源基因表达者;(3)获取囊胚细胞,作为ES细胞的移植受体;(4)通过显微操作将ES细胞注入到囊胚期胚胎的腔内,与其细胞团紧靠在一起,成为嵌合体;(5)将注射过的胚胎,经培养后筛选无发育缺损的囊胚,移植到交配第三天的假孕受体鼠子宫内,培育出转基因动物。
2、小鼠的基因敲除(gene knockout)流程基因敲除是采用同源重组的方法,用体外合成的无效基因或突变基因取代相应正常基因,再应用转基因方法孵育出转基因动物。
简:将灭活的基因导入ES细胞中,使这一灭活基因通过同源重组取代原有的目的基因,筛选到基因已定点灭活的细胞后,通过显微注射将细胞注入小鼠囊胚中。
细胞在小鼠囊胚中参与胚胎的发育,最终形成嵌合体小鼠。
由于嵌合体小鼠的一部分细胞来源于ES细胞,所以通过小鼠培育即可获得纯合子基因敲除小鼠。
繁:a.获取ES细胞b.基因载体的构建:把目的基因和与细胞内靶基因特异片段同源的DNA分子都重组到带有标记基因的载体上,此重组载体即为打靶载体。
c.将基因打靶载体通过一定的方式(常用电穿孔法)导入同源的胚胎干细胞(EScell)中,使外源DNA与胚胎干细胞基因组中相应部分发生同源重组,将打靶载体中的DNA序列整合到内源基因组中从而得以表达。