第二节膜表面受体介导的信号转导
- 格式:doc
- 大小:738.00 KB
- 文档页数:46



细胞生物学中的膜受体结构与信号转导细胞膜受体是生物学研究中的一个重要课题,它们对于细胞的正常生理功能以及疾病的形成都具有重要的影响。
膜受体是指位于细胞膜上的蛋白质,能够响应外界信号分子的结合并传递信号,从而控制细胞的生理活动。
这些外界信号分子可以是激素、神经递质、细胞因子等。
膜受体的结构及其信号转导机制是近年来生物学研究的热门领域之一。
一、膜受体结构膜受体分为离子通道型、酪氨酸激酶型、酰基酶型、G蛋白偶联型等几种类型。
其中,G蛋白偶联型受体在人体中是最广泛存在的一类受体。
它们的结构都由七个跨膜螺旋结构组成,可使蛋白质从细胞膜内部贯穿到膜外,每个螺旋结构中具有不同的氨基酸序列。
这些螺旋结构组成一个类似于枪托的外形,被称为“7TMB”(seven transmembrane helices bundle)结构。
7TMB结构是G蛋白偶联型受体与其他一些受体分子的独特标志。
二、信号转导信号转导是指外界信号分子与膜受体结合后,膜受体内部启动级联反应的过程,将信号传递给细胞内部,引起细胞活动的变化。
G蛋白偶联型受体是信号转导过程中重要的参与者。
它们通过膜上的七个跨膜结构与G蛋白相互作用,从而引起以下信号转导事件:1.激活腺苷酸酶(adenylyl cyclase):当一个激动剂分子与负责激动剂作用的G蛋白偶联型受体结合时,该受体活跃化,离开G蛋白并促进腺苷酸酶的活化,催化cAMP(环磷酸腺苷)的形成,cAMP 激活蛋白激酶A(PKA)而引发细胞内的后续反应过程。
2.使钙离子脱敏:肌肉收缩的调节和其他生理过程中,钙离子扮演了重要的角色。
G蛋白偶联型受体上的钙离子通道蛋白与 G蛋白偶联型受体互作,导致钙离子水平在细胞内缓慢下降。
这就使得钙离子接收体在没有外部钙离子贡献的情况下,难以响应内源性信号分子的作用。
3.激活酪氨酸激酶:一些受体是酪氨酸激酶受体。
受到激活剂分子的激励后,它们会召集酪氨酸激酶进入细胞内。
该过程又分为两次活化,第一次活化发生在受体和酪氨酸激酶之间,第二次则是在酪氨酸激酶与其底物的内部反应。


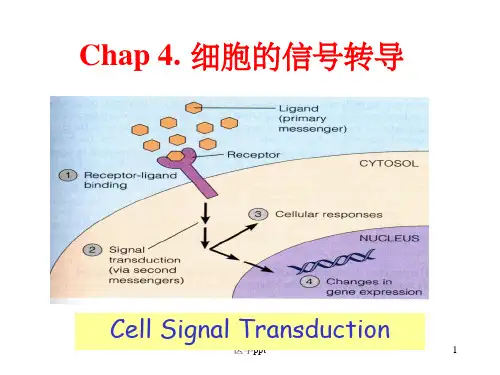

细胞信号传导的途径细胞信号传导是维持生命活动的重要过程,它使得细胞能够感知和响应外界环境的各种刺激。
一旦信号被传递到细胞内部,它将引发一系列分子事件,最终导致特定的细胞行为或生物学功能的改变。
本文将针对细胞信号传导的主要途径进行探讨。
一、细胞膜上的受体介导的信号传导细胞膜上的受体是感知外界刺激的主要途径之一。
它们可以分为离子通道受体和酪氨酸激酶(RTK)受体两大类。
离子通道受体可以通过改变细胞膜上的离子通道通透性,使离子进出细胞,从而改变细胞内离子浓度和电位,引发信号传导。
而RTK受体则可以激活下游的信号转导蛋白,如Ras/MAPK通路和PI3K/AKT通路等,进一步调控细胞的增殖、分化和存活等生理功能。
二、胞内受体介导的信号传导除了细胞膜上的受体,胞内受体也起着重要的作用。
胞内受体主要指核受体家族,如雌激素受体(ER)和孕酮受体(PR)等。
这些受体位于细胞核内,当配体如激素结合到受体上时,受体会发生构象改变,形成激活状态。
激活的受体-配体复合物将与DNA结合,调控特定基因的转录,进而影响细胞的生物过程和功能。
三、细胞内信号传导通路在细胞内部,信号通常通过一系列信号转导通路进行传递。
其中,Ras/MAPK通路和PI3K/AKT通路是最为经典和重要的两条通路。
Ras/MAPK通路的激活依赖于RTK受体的激活,它能够调节基因表达、细胞生长和分化等过程。
PI3K/AKT通路则与细胞的存活、增殖和细胞周期调控密切相关。
四、细胞间的信号传导除了细胞内信号传导,细胞间的信号传导也是维持组织和器官功能的重要手段。
细胞间的信号传导可以通过细胞间直接接触的方式,如通过细胞间连接和细胞间通道进行信号传递。
此外,细胞还可以通过释放信号分子,如细胞因子、神经递质等,在周围环境中扩散,并作用于距离较远的细胞,实现信号传导。
细胞信号传导是一个复杂而精密的过程,涉及多个信号分子和通路的相互作用。
了解细胞信号传导的途径对于揭示细胞功能和生物学过程具有重要意义。

受体介导的信号转导随着现代科学技术的不断发展,受体介导的信号转导已经成为细胞学研究的一个重要领域。
对于细胞内外环境的信息传递以及细胞生命活动的调节、控制和调控,信号转导扮演着至关重要的角色,而受体介导的信号转导则是信号转导中最为重要的环节之一。
一、受体介导的信号转导的定义和作用受体介导的信号转导是指生物体内分子信号在外界刺激下,通过细胞膜上的受体分子传递到细胞内,从而激活一系列的生化反应,最终调控细胞的生理功能和代谢活动。
在这个过程中,受体分子扮演着关键的角色,它们既能识别具体的外界信号,又能启动一系列下游的信号传递分子,促进生物体对外界环境变化的适应和反应。
二、受体介导的信号转导的类型目前,关于受体介导的信号转导存在着多种分类方式,其中常见的有以下几种:1、离子通道型受体介导的信号转导这种类型的受体介导的信号转导,传递过程中虽然不需要通过多种信号转导蛋白,但是具有快速、短暂和在静息态下低能耗的特点,常见的受体有神经递质受体和离子通道,在激活时通过控制物体表面区域的离子浓度来促进离子的通道形成,从而实现信号的快速传递。
2、酶联受体介导的信号转导酶联受体介导的信号转导常见于生长因子和激素家族,通过它们,能够引发一系列的生化反应过程,从而调节生物体的生长发育和细胞增殖等生命活动。
酶联受体介导的信号转导机理比离子通道类型更为复杂,需要在激活过程中依赖其他多种信号转导蛋白的共同作用。
3、G蛋白偶联受体介导的信号转导这种类型的信号转导,可以在细胞表面上识别多种化学物质,包括激素、神经递质、细胞杀伤因子等,并能启动一系列下游的信号传递过程,以实现对于外界环境的适应和反应。
在G蛋白偶联受体介导的信号转导中,G蛋白的激活是关键的一环,它能够调节去极化和建立离子梯度等一系列过程,从而影响下游的生化反应过程。
三、受体介导的信号转导的应用由于受体介导的信号转导在细胞生物学和微生物学中的重要作用,其相关研究也已经成为了生物技术和医学研究的热点之一。

第二节膜表面受体介导的信号转导亲水性化学信号分子:* 有神经递质、蛋白激素、生长因子等* 它们不能直接进入细胞只能通过膜表面的特异受体,传递信号使靶细胞产生效应膜表面受体主要有三类(图8-7):①离子通道型受体(ion-channel-linked receptor)存在于可兴奋细胞②G蛋白耦联型受体(G-protein-linked receptor)③酶耦联的受体(enzyme-linked receptor)后2类存在于大多数细胞在信号转导的早期表现为一系列蛋白质的逐级磷酸化使信号逐级传送和放大。
图8-7 膜表面受体主要有3类一、离子通道型受体离子通道型受体(图8-8):* 离子通道的受体即,配体门通道(ligand-gated channel)* 主要存在于神经、肌肉等,可兴奋细胞其信号分子为神经递质* 神经递质+受体,而改变通道蛋白的构象离子通道,开启or关闭改变质膜的离子通透性瞬间(1/1000秒),胞外化学信号→电信号继而改变突触后细胞的兴奋性* 位于细胞膜上的受体,一般4次跨膜位于质网上的受体,一般6次跨膜* 离子通道型受体分为阳离子通道,如乙酰胆碱、谷氨酸、五羟色胺的受体阴离子通道,如甘氨酸&γ-氨基丁酸的受体* 如:乙酰胆碱受体(图8-9、10)以三种构象存在2分子乙酰胆碱的结合使通道处于开放构象但受体处于通道开放构象状态,时限十分短暂在几十毫微秒,又回到关闭状态然后,乙酰胆碱与受体解离受体恢复到初始状态做好重新接受配体的准备图8-8 离子通道型受体synaptic cleft:突触间隙图8-9 乙酰胆碱受体结构模型图8-10 乙酰胆碱受体的三种构象图8-11 神经肌肉接触点处的离子通道型受体二、G蛋白耦联型受体G蛋白* 3聚体GTP结合的调节蛋白,简称G蛋白(trimeric GTP-binding regulatory protein)由α、β、γ,3个亚基组成位于质膜胞质侧* α 和γ亚基,通过共价结合的脂肪酸链尾部,与细胞膜结合* G蛋白在信号转导过程中,起着分子开关的作用(图8-12)当α亚基与GDP结合时,处于关闭状态与GTP结合时,处于开启状态* α亚基具有GTP酶活性其GTP酶的活性能被RGS(regulator of G protein signaling)增强RGS也属于GAP(GTPase activating protein)图8-12 G蛋白分子开关G蛋白耦联型受体,为7次跨膜蛋白(图8-13)* 受体胞外结构域,识别胞外信号分子,并与之结合* 胞结构域,与G蛋白耦联调节相关酶活性在细胞产生第二信使将胞外信号跨膜→胞* G蛋白耦联型受体包括,多种神经递质、肽类激素&趋化因子的受体在味觉、视觉&嗅觉中,接受外源理化因素的受体亦属G蛋白耦联型受体图8-13 G蛋白耦联型受体为7次跨膜蛋白由G蛋白耦联受体介导的细胞信号通路主要包括:cAMP信号通路磷脂酰肌醇信号通路(一)cAMP信号途径在cAMP信号途径中胞外信号&相应受体结合→调节腺苷酸环化酶活性→通过第二信使cAMP水平的变化→将胞外信号→胞信号1、cAMP信号的组分①激活型激素受体(Rs)or 抑制型激素受体(Ri)②活化型调节蛋白(Gs)or 抑制型调节蛋白(Gi)③腺苷酸环化酶相对分子量为150KD的糖蛋白跨膜12次在Mg2+ 或Mn2+的存在下腺苷酸环化酶,催化ATP→cAMP(图8-14)。

膜受体介导的信号转导途径及信号分子下载提示:该文档是本店铺精心编制而成的,希望大家下载后,能够帮助大家解决实际问题。
文档下载后可定制修改,请根据实际需要进行调整和使用,谢谢!本店铺为大家提供各种类型的实用资料,如教育随笔、日记赏析、句子摘抄、古诗大全、经典美文、话题作文、工作总结、词语解析、文案摘录、其他资料等等,想了解不同资料格式和写法,敬请关注!Download tips: This document is carefully compiled by this editor. I hope that after you download it, it can help you solve practical problems. The document can be customized and modified after downloading, please adjust and use it according to actual needs, thank you! In addition, this shop provides you with various types of practical materials, such as educational essays, diary appreciation, sentence excerpts, ancient poems, classic articles, topic composition, work summary, word parsing, copy excerpts, other materials and so on, want to know different data formats and writing methods, please pay attention!膜受体介导的信号转导途径及信号分子引言膜受体介导的信号转导途径是细胞内外信息传递的重要机制之一,其参与调节细胞的生长、分化、凋亡等生命活动。
12.2细胞膜受体介导的信号转导同学们好!上一讲我已经学习了细胞信号转导系统中的关键成员:细胞膜表面受体。
今天我们来进一步学习细胞膜受体介导的信号转导通路。
下面给同学介绍两条最主要的细胞信号转导“热线”,即:G蛋白偶联受体(GPCR)信号通路和受体酪氨酸激酶(RTK)信号通路。
一、GPCR信号通路GPCR信号通路是类型最多样和作用最普遍的细胞信号转导途径。
主要包括以下环节:(一)第一信使的跨膜信号转导胞外各种信号分子或称为配体(ligand)作为第一信使,可与相应的细胞膜表面GPCR特异性结合,触发受体蛋白构象变化,引起胞内区结合的G蛋白发生结构与活性变化,进而诱导邻近的效应靶蛋白发生功能转换,实现胞外信号的跨膜转导。
上一讲我们已经讲过GPCR受体的基本结构和特点,也学习了G蛋白的分子开关机制。
在此我们来看看GPCR与配体结合后是如何引起G蛋白变构与活性变化的。
1、G蛋白变构。
G蛋白由α、β、γ三个亚基构成,其中α很独立,而β与γ则形成异二聚体,它们三个在安静状态下聚在一起均通过共价结合的脂分子锚定在细胞膜上。
此时,α结合GDP使G蛋白处于非活化状态。
G蛋白作为分子开关,一旦α结合上GTP,则导致α与βγ二聚体解离,呈现出活化状态。
2、G蛋白活性变化。
大体上来讲,G蛋白活性变化分为3步:1)受体激活。
即配体结合GPCR后,引起受体构象变化,胞内区与G蛋白结合,诱导α与GDP结合力大大下降。
2)G蛋白激活。
当GDP从α上脱离,并在鸟苷交换因子GEF的作用下结合上GTP后,整个G蛋白即被激活解离,特别是游离的α可找到其附近的效应靶分子,如腺苷酸环化酶(adenylyl cyclase,AC),两者结合,激活AC活性。
3)G蛋白失活。
α还具有GTPase酶活性,α结合GTP仅仅数秒之后即可把GTP水解成GDP,α与AC脱离,又恢复到与βγ二聚体结合的失活状态。
上述步骤顺序进行,循环往复,完成胞外信号的跨膜信号转导。
细胞表面受体介导的信号通路步骤细胞表面受体介导的信号通路是细胞内外信息传递的重要机制之一。
当外界刺激物(如激素、神经递质等)结合到细胞表面受体上时,会触发一系列的信号转导步骤,最终导致细胞内特定反应的发生。
本文将详细介绍细胞表面受体介导的信号通路步骤。
1. 受体激活与配体结合信号通路的起始点是外界刺激物(配体)与细胞表面受体结合。
配体可以是激素、神经递质或其他分子。
当配体与受体结合时,会引发受体构象变化,从而激活受体。
2. 受体激活后的自磷酸化受体激活后,其内部区域会发生自磷酸化作用。
这意味着受体上存在磷酸化位点,并且在激活状态下会被磷酸化。
这种自磷酸化可以通过多种方式实现,如自身蛋白激酶活性或与其他蛋白激酶的相互作用等。
3. 激活下游信号分子受体的自磷酸化会导致下游信号分子的激活。
一般来说,这些下游信号分子是蛋白激酶,它们可以进一步传递信号,将信息传递到细胞内部。
4. 信号放大与传导激活的下游信号分子会进一步放大和传导信号。
这通常通过级联反应实现,其中一个被激活的蛋白激酶可以磷酸化和激活另一个蛋白激酶,从而形成一个信号放大和传导的链式反应。
5. 调节因子介入在信号通路中,还存在一些调节因子,它们可以增强或抑制信号传导。
这些调节因子可以是其他蛋白质、离子或小分子。
它们通过与下游信号分子相互作用,进一步调控整个信号通路的效果。
6. 下游效应发生在信号通路中发生了一系列级联反应后,会触发细胞内的下游效应。
这些效应可以是细胞内的生化反应、基因表达的改变或细胞行为的改变等。
下游效应是信号通路最终的结果。
7. 负反馈调节为了保持信号通路的平衡和稳定,通常会存在一些负反馈调节机制。
这些机制可以通过抑制受体活性、降解信号分子或调节信号分子的磷酸化状态等方式实现,从而限制信号传导的强度和持续时间。
8. 信号终止一旦下游效应发生并达到所需的程度,信号通路需要被终止。
这可以通过多种方式实现,如受体内外区域结构的变化、磷酸化位点去磷酸化等。