基因的连锁互换规律
- 格式:ppt
- 大小:537.00 KB
- 文档页数:20

遗传学三个基本规律的主要内容
遗传规律有三大规律,分别是基因分离定律,基因自由组合定律,和基因连锁、交换定律。
第一规律,分离定律是遗传学中最基本的一个规律,它从本质上阐明了控制生物性状的遗传物质是以自成单位的基因活动的,基因作为遗传单位在体细胞中是成双的,它在遗传上具有高度的独立性,因此在减数分裂的配子形成过程中,成对的基因在杂种细胞中能够彼此互不干扰,独立分离,通过基因重组,在子代继续表现各自的作用,这一规律从理论上说明了生物界由于杂交和分离所出现的变异的普遍性。
第二规律,是自由组合定律,就是当具有两对或者更多对相对性状的亲本杂交,在此一代产生配子时,在等位基因分离的同时,非同源染色体上的非等位基因表现为自由组合。
第三个定律,就是连锁与互换定律,连锁与互换定律是指原来为同一亲本所具有的两个性状,在f2中常常有连系在一起遗传的倾向,这种现象成为连锁遗传。
连锁遗传定律的发现,证实了染色体是控制性状遗传基因的载体,通过交换的测定,进一步证明了基因在染色体上具有一定的距离的顺序,呈直线排列。

基因的连锁定律
基因的连锁定律是指在遗传过程中,位于同一染色体上的不同基因在生殖细胞形成(减数分裂)时,往往连在一起作为一个单位进行传递。
这个定律主要由孟德尔的遗传规律发展而来,其中包括基因的分离定律和自由组合定律。
基因的连锁定律主要包括以下三个方面:
1. 基因的连锁:在减数分裂过程中,同源染色体上的等位基因随着非姐妹染色单体的交换而发生互换,从而形成基因的重组。
2. 基因的自由组合:在减数分裂的过程中,同源染色体上的等位基因彼此分离。
在分离之前,可能发生部分染色体的交叉互换。
在此基础上,非同源染色体上的非等位基因进行自由组合,形成各种组合的配子。
3. 基因的交换:位于同一染色体上的基因在生殖细胞形成过程中,一对同源染色体上的不同对等位基因之间可以发生交换,这种现象称为交换律或互换律。
基因的连锁定律和自由组合定律是遗传学的基本定律,它们共同解释了生物体在遗传过程中基因的传递和组合规律。
这些定律在生物多样性和遗传育种等领域具有重要的应用价值。

连锁与互换定律1、连锁遗传:原来在亲本中组合在一起的两个性状在F2中有连在一起遗传的倾向,称连锁遗传。
连锁相包括互引相〔AB、ab〕、互斥相〔Ab、aB〕。
2、亲本型:与两亲本一样的性状表现型称为亲本型;不同的称为重组型。
3、完全连锁遗传:仅有亲本型,缺少重组型,eg:仅见于雄果蝇、雌家蚕。
4、不完全连锁遗传:在连锁遗传的同时发生性状的交换和重组;绝大多数生物为不完全连锁遗传。
5、利用测交法验证连锁遗传现象:特点:连锁遗传的表现为: 两个亲本型配子数是相等,> 50%; 两个重组型配子数相等,< 50%。
亲组合类型多, 重组合类型出现少6、交换值(Cv):指不完全连锁的两基因间发生交换的频率(百分率,平均次数) 。
重组值(Rf):不完全连锁的双杂合体产生的重组型配子数占总配子数的比率(百分率)。
通常又把交换值称为重组值。
但严格说,交换值不能等同于重组值,因为假设两个基因座之间相距较远,其间发生偶数次多重交换时,结果不形成重组型配子,用重组值代表交换值会造成偏低的估计。
7、连锁群:不能进展自由组合的基因群〔位于同一染色体上的基因群)。
特点:一种生物连锁群的数目与染色体的对数是一致的。
即有n对染色体就有n个连锁群。
8、染色体作图:把染色体的多种基因相互之间的排列顺序确定下来。
连锁遗传的特征1〕摩尔根连锁互换是经典遗传学第三定律,是孟德尔自由组合定律的补充;2〕发生在两对或以上基因间,且基因在染色体上线性排列;3〕连锁基因发生在同一对同源染色体上;4〕减数分裂偶线期,同源染色体联会,非姐妹染色单体间的互换是形成重组型的分子根底;5〕两对基因座间距离越大,交换概率越大、连锁性越弱;6〕完全交换即为自由组合,完全不交换即为完全连锁情形;染色体作图〔基因定位〕方法包括两点测交法和三点测交法计算基因间相对距离(1)非等位基因在染色体上排列的直线距离与基因间的互换率大小有关;(2)遗传学上规定,以互换率的1%作为一个遗传单位将基因定位在一条直线上。

第五章连锁与互换规律1910年摩尔根将果蝇白眼基因定位于X上后,又发现一些伴性遗传基因,证明X上确有许多基因(人有23对染色体和十万个基因)。
提出问题:(1)同一染色体上众多基因如何排列(2)遗传传递有何规律经研究得到结论:(1)基因论(2)连锁交换规律第一节基因的连锁与交换一、连锁遗传现象(一)连锁遗传现象发现1906年英国学者贝特森(Bateson)和潘耐特(Pannett)研究香豌豆两对性状遗传时,首先发现的。
分离比不符花颜色紫色P对红色p显性,花粉粒形状长形L对圆形l显性。
F2合9:3:3:1,亲组合类型较多,重组合类型偏少。
原为同一亲本的两个性状,在F中常常有联系在一起的倾向,这说明来自同2一亲本的基因,有较多的在一起传递的可能。
但贝特森和潘耐特未能提出科学的产生配子数找原因,4种配子数目不等,亲组合类型多,解释。
摩尔根认为应从F1重组类型少。
(二)摩尔根的实验果蝇翅的长短,复眼的颜色长翅Vg,残翅vg,红色复眼Pr,紫色复眼pr解释相引组杂交红眼雄果蝇产生两种数目相同的配子,测交后代1:1,红眼雌果蝇产生四种配子,但亲组型配子Vg Pr和vg pr多,而重组型配子Vg pr和vgPr少。
同理可解释相斥组杂交结果。
果蝇翅的长短和眼的颜色是两对相对性状,这两对相对性状在杂交后代中具有某种程度的相关性或相连性,把这种现象叫连锁。
把这两对相对性状叫连锁性状。
控制连锁性状的基因叫连锁基因,连锁基因位于同一对同源染色体上。
连锁基因之间能够发生交换的连锁叫不完全连锁(雌果蝇),不能发生交换的连锁叫完全连锁(雄果蝇、雌家蚕)。
目前人们经过研究发现仅有两种动物雄果蝇和雌蚕属于完全连锁,不发生交换。
相引组:两个显性在一起,两个隐性在一起配成的杂交组合。
相斥组:一个显性一个隐性和一个隐性一个显性在一起配成的杂交组合。
(三)连锁的细胞学基础同一染色体上非等位基因随染色体一起遗传——完全连锁,又能按一定比例发生交换而重组——不完全连锁。


基因连锁互换定律1. 引言基因连锁互换定律是遗传学中重要的定律之一。
它描述了基因在染色体上的连锁分布以及连锁分离的频率。
本文将探讨该定律的基本概念、实验验证以及其在遗传研究中的应用。
2. 基本概念2.1 基因连锁基因连锁指的是在染色体上,位于同一条染色体上的基因之间的相对位置关系保持恒定。
这意味着这些基因在遗传上很有可能一起传递给下一代。
2.2 互换互换是指两条同源染色体之间的交叉互换事件。
在互换事件中,两条染色体上的相同位点上的DNA片段通过交换破裂重连的方式重新组合。
互换可以导致基因连锁的打破,使得原本连锁在一条染色体上的基因出现重新组合。
3. 实验验证基因连锁互换定律最早是通过托马斯·亨特·摩尔根(Thomas Hunt Morgan)的果蝇实验得出的。
通过对果蝇基因型的观察和分析,摩尔根发现某些基因总是共同遗传,这与经典遗传学中的独立性原则相违背。
3.1 果蝇实验摩尔根在实验中选择了具有明显表型差异的果蝇,例如翅膀的颜色。
他通过交配不同表型的果蝇,观察它们后代的表现,并推断基因的连锁关系。
3.2 结果与结论通过大量的实验证明,果蝇的一对基因位点可以遗传的同时,也可以发生连锁互换。
这一发现支持了基因连锁互换定律的存在。
4. 基因连锁互换定律的意义基因连锁互换定律的发现对于遗传学的发展产生了重要影响。
4.1 进化的证据基因连锁互换定律的存在证明了生物进化过程中的基因发生重组,从而导致物种多样性的增加。
4.2 遗传图谱的构建基因连锁互换定律为遗传学家构建遗传图谱提供了重要依据。
通过观察基因连锁互换事件的频率,可以推断基因之间的距离以及相对位置。
4.3 基因定位和基因治疗基因连锁互换定律也为研究基因定位和开展基因治疗提供了基础。
通过对基因连锁的研究,科学家可以确定染色体上具体基因的位置,从而更好地理解基因功能和相关疾病。
5. 总结基因连锁互换定律是遗传学的重要定律之一。
通过果蝇实验和大量实验证据,科学家证明了基因之间的连锁关系以及连锁互换的发生。


必修本第36课时 基因的连锁和交换定律知识精华完全连锁 现象:F 1灰身长翅♂(BbVv)×黑身残翅♂(bbvv) →1灰身长翅(BbVv):1黑身残翅(bbvv)原因: (只有亲本类型,没有重组类型) ♂)−−−→−减数分裂1:1 , F 1雄果蝇产配子时,同一条染色体上的不同基因常连在一起不分离,所以只有亲本类型的配子没有重组类型的配子。
不完全连锁现象:F 1灰身长翅雌(BbVv)×黑身残翅雄(bbvv)↓灰长(BbVv):黑残(bbvv):灰残(Bbvv):黑长(bbVv)42 : 42 : 8 : 8(亲本类型特别多,重组类型特别少)原因:B V B V b v B v b V(♀) : :F 1雌果蝇产配子时,部分初级卵母细胞的同一条染色体上不同基因因同源染色体的非姐妹染色单体之间的交叉互换,即产生亲本类型的配子又产生了重组类型的配子。
理论意义:基因的不完全连锁即互换能引起基因重组,使生物产生变异实践意义:根据育种目标选择杂交亲本时,必须考虑性状间的连锁关系题例领悟例题:基因型为的精原细胞120个,其中若有30个在形成精子过程中发生互换,在正常发育下能产生亲本基因型和重组基因型的精子数目依次是( ) A 、480、480、60、60 B 、210、210、30、30C 、90、90、30、30D 、240、240、60、60解析:120个精原细胞最多产生480个精子,所以A 、D 选项错误。
由于交叉(互换)发生在同源染色体的两条非姊妹染色单体之间,一个基因型为 B v b vB v b vb v 42 42 8B D b dB D精原细胞由于发生了互换,产生的四个精子中(BD ,bd ,Bd ,Bd),只有两个是重组基因型的精子(Bd ,Bd)。
30个精原细胞形成的120个精子中,重组基因型的精子(Bd ,bD)应各有30个。
而其余则是两亲本基因型的配子。
该精原细胞的重组率(或交换值)为:(30+30)/480 ×100%=12.5% 答案:B自我评价一、选择题1、在两对相对性状的遗传实验中,如果F 1的测交后代的表现型有两种且比为1:1,那么这个遗传的类型应属于 ( )A 、基因的分离定律B 、基因的自由组合定律C 、基因的完全连锁D 、基因的连锁和交换2、基因型为 的个体,在形成配子过程中,有20%的初级精母细胞发生了互换。

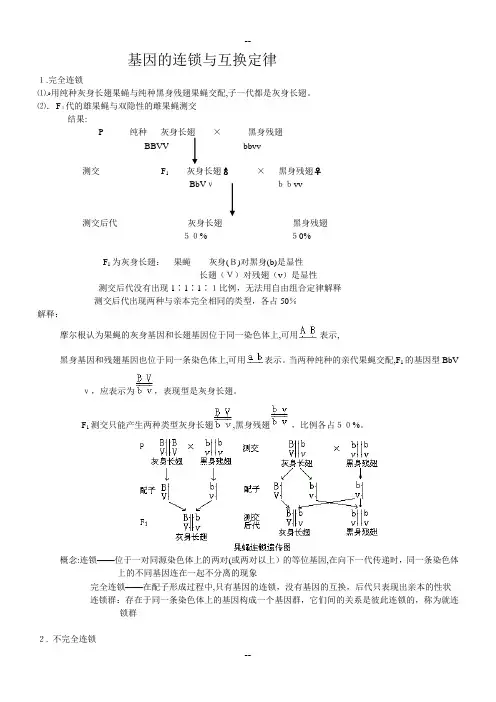
基因的连锁与互换定律之答禄夫天创作1.完全连锁⑴.用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,子一代都是灰身长翅。
⑵.F1代的雄果蝇与双隐性的雌果蝇测交结果:P 纯种灰身长翅×黑身残翅BBVV bbvv测交 F1灰身长翅♂×黑身残翅♀BbVv bbvv测交后代灰身长翅黑身残翅50% 50%F1为灰身长翅:果蝇灰身(B)对黑身(b)是显性长翅(V)对残翅(v)是显性测交后代没有出现1∶1∶1∶1比例,无法用自由组合定律解释测交后代出现两种与亲本完全相同的类型,各占50%解释:摩尔根认为果蝇的灰身基因和长翅基因位于同一染色体上,可用暗示,黑身基因和残翅基因也位于同一条染色体上,可用暗示。
当两种纯种的亲代果蝇交配,F1的基因型BbVv,应暗示为,表示型是灰身长翅。
F1测交只能发生两种类型灰身长翅,黑身残翅,比例各占50%。
概念:连锁——位于一对同源染色体上的两对(或两对以上)的等位基因,在向下一代传递时,同一条染色体上的分歧基因连在一起不分离的现象完全连锁——在配子形成过程中,只有基因的连锁,没有基因的互换,后代只表示出亲本的性状连锁群:存在于同一条染色体上的基因构成一个基因群,它们间的关系是彼此连锁的,称为就连锁群2. 不完全连锁用子一代雌性个体进行测交实验结果:P 纯种灰身长翅×黑身残翅BBVV bbvv测交 F1灰身长翅♀×黑身残翅♂BbVv bbvv测交后代灰身长翅黑身残翅42% 42%灰身残翅黑身长翅8% 8%后代出现四种性状,其中亲本类型占多数,新组合类型占少数。
解释:细胞在进行减数分裂形成配子的过程中,减数分裂第一期前期,同源染色体联会,形成四分体。
联会复合体中同源染色体间的非姐妹染色单体间会发生染色单体的交叉互换,在交换区段上的基因随染色体发生交换,这种交换发生新的基因组合。
交叉互换后形成四种配子,其数量相同;其中有两种配子是亲本类型,两种配子是重组合类型,各占一半。
基因的连锁与互换定律1.完全连锁⑴.ﻩ用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,子一代都是灰身长翅。
⑵. F1代的雄果蝇与双隐性的雌果蝇测交结果:P纯种灰身长翅×黑身残翅BBVV bbvv测交F1灰身长翅♂×黑身残翅♀BbVvbbvv测交后代灰身长翅黑身残翅50%50%F1为灰身长翅:果蝇灰身(B)对黑身(b)是显性长翅(V)对残翅(v)是显性测交后代没有出现1∶1∶1∶1比例,无法用自由组合定律解释测交后代出现两种与亲本完全相同的类型,各占50%解释:摩尔根认为果蝇的灰身基因和长翅基因位于同一染色体上,可用表示,黑身基因和残翅基因也位于同一条染色体上,可用表示。
当两种纯种的亲代果蝇交配,F1的基因型BbV v,应表示为,表现型是灰身长翅。
F1测交只能产生两种类型灰身长翅,黑身残翅,比例各占50%。
概念:连锁——位于一对同源染色体上的两对(或两对以上)的等位基因,在向下一代传递时,同一条染色体上的不同基因连在一起不分离的现象完全连锁——在配子形成过程中,只有基因的连锁,没有基因的互换,后代只表现出亲本的性状连锁群:存在于同一条染色体上的基因构成一个基因群,它们间的关系是彼此连锁的,称为就连锁群2. 不完全连锁用子一代雌性个体进行测交实验结果:P纯种灰身长翅×黑身残翅BBVVbbvv测交F1灰身长翅♀×黑身残翅♂BbVv bbvv测交后代灰身长翅黑身残翅42% 42%灰身残翅黑身长翅8% 8%后代出现四种性状,其中亲本类型占多数,新组合类型占少数。
解释:细胞在进行减数分裂形成配子的过程中,减数分裂第一期前期,同源染色体联会,形成四分体。
联会复合体中同源染色体间的非姐妹染色单体间会发生染色单体的交叉互换,在交换区段上的基因随染色体发生交换,这种交换产生新的基因组合。
交叉互换后形成四种配子,其数量相同;其中有两种配子是亲本类型,两种配子是重组合类型,各占一半。
但是在生殖细胞形成过程中,发生交叉互换的性母细胞并不多,所以,减数分裂产生的配子中亲本类型的配子最多,重组合类型的配子占少数。

基因连锁互换定律基因连锁互换定律是指在同一染色体上的基因在遗传过程中会联合遗传,即它们之间存在连锁关系。
这一定律是由托马斯·亨特·摩尔根和他的同事们在20世纪初期通过对果蝇的研究发现的。
该定律对于遗传学和进化生物学的发展产生了深远影响,也为人类遗传疾病的研究提供了理论基础。
一、基因连锁互换定律的发现1.1 摩尔根实验托马斯·亨特·摩尔根是美国哥伦比亚大学生物学教授,他和他的同事们通过对果蝇(Drosophila melanogaster)进行实验,发现了基因连锁互换定律。
他们选取了两组显性突变体(眼睛颜色和翅膀形态),并将它们杂交,得到了F1代。
然后再将F1代进行自交,得到F2代。
他们发现,F2代中出现了不同于父母代的表型比例,并且这些表型出现的频率并不相等。
1.2 连锁关系的发现通过对F2代的分析,摩尔根和他的同事们发现,这些表型不是独立地遗传的,而是存在某种联系。
他们将这种联系称为连锁关系。
具体来说,他们发现了两个显性突变体(红眼和长翅膀)在同一染色体上,并且它们之间的距离很近,因此它们在遗传过程中总是一起遗传。
1.3 基因连锁互换定律的提出通过对果蝇遗传实验的研究,摩尔根和他的同事们提出了基因连锁互换定律:在同一染色体上的基因会联合遗传,即它们之间存在连锁关系。
这意味着,在一些情况下,不同基因之间并不是完全独立地遗传。
二、基因连锁互换定律的解释2.1 染色体结构要理解基因连锁互换定律,首先需要了解染色体结构。
每个细胞核中都有一定数量的染色体,其中包含了DNA分子。
DNA分子由四种碱基组成:腺嘌呤、胸腺嘧啶、鸟嘌呤和胞嘧啶。
这些碱基按照一定顺序排列,形成了基因。
每个染色体上都有许多基因,它们的位置是固定的。
2.2 染色体的配对在有性生殖过程中,每个个体都会从父母那里继承一条染色体。
因此,在一个细胞中,每条染色体都有与之对应的另一条染色体。
这两条染色体组成了一对同源染色体。
基因的连锁和交换定律
基因的连锁定律和交换定律是遗传学中重要的定律。
基因的连锁定律是指,当两种不同基因位于同一染色体上时,它们会一起遗传。
这意味着,如果一对基因位于同一染色体上,它们不会在细胞分裂过程中分离,而是一起遗传给下一代。
基因交换定律是指,当两种不同基因位于不同染色体上时,它们会在细胞分裂过程中进行交换。
这意味着,如果一对基因位于不同染色体上,它们会在细胞分裂过程中互相交换,从而使它们能够在下一代中分离遗传。
这两个定律是遗传学中重要的基础,它们有助于解释基因遗传的规律,并为研究遗传疾病和基因工程等领域奠定了基础。
基因的连锁定律是由英国遗传学家Thomas Hunt Morgan在20世纪初发现的。
他通过对果蝇(Drosophila melanogaster)的研究发现,当两种不同基因位于同一染色体上时,它们会一起遗传。
这种现象被称为基因连锁。
基因交换定律也是由Thomas Hunt Morgan发现的。
他发现,当两种不同基因位于不同染色体上时,它们会在细胞分裂过程中进行交换。
这种现象被称为基因
交换。
这两个定律是遗传学中重要的基础,它们有助于解释基因遗传的规律,并为研究遗传疾病和基因工程等领域奠定了基础。
连锁定律和交换定律解释了基因遗传的规律,这对基因工程和分子生物学的研究具有重要意义。
例如,在基因工程中,利用这两个定律可以更好地设计基因治疗策略。
在分子生物学中,这两个定律可以帮助研究者更好地理解基因组学和基因调控机制。
基因的连锁与互换规律从图中可以看出:F 1雌果蝇(灰身长翅)在减数分裂产生配子时,绝大多数的初级卵母细胞中连锁基因不发生互换,即为连锁遗传,所以减数分裂时产生两种数量相等的亲本类型配子:BV :bv=1:1(多数)。
少数的初级卵母细胞中的连锁基因发生互换,则会产生四种配子为:BV :bv :Bv :bV=1:1:1:1(少数)。
注意:其中有50%是亲本类型配子,50%是重组类型配子。
所以把上述两种情况相加,F 1雌果蝇共产生四种配子:BV :bv :Bv :bV=多:多:少:少。
如把初级卵母细胞总数看成100,假设不发生交叉互换的初级卵母细胞为68个(68%),则产生的卵细胞为BV 和bv 各34个。
发生互换的初级卵母细胞为32个(32%),则产生的卵细胞为BV 、bv 、Bv 、bV 各8个。
两种情况相加,F 1产生的卵细胞为BV :42%、bv :42%、Bv :8%、Bv :8%。
这样就很容易理解F 1灰身长翅(雌)与黑身残翅(雄)测交后产生的后代的种类及比例:灰身长翅:黑身残翅:灰身残翅:黑身长翅=42%:42%:8%:8%。
学生根据此图也比较容易理解交换值的含义及公式在应用中的变化。
交换值= F 1重组型配子数/ F 1总配子数×100%=测交后代中的重组型数/测交后代总数×100% =1/2发生交换的性母细胞的百分比以此图为例,F 1雌果蝇的初级卵母细胞有32%发生了互换,则连锁基因间的交换值为16%(8%+8%)。
(如果其它生物的精母细胞发生了互换,与卵母细胞同理)练习1:在100个精母细胞的减数分裂中,有50 的细胞的染色体发生了一次互换,在所形成的配子中,换型的百分率占 ( C )A 、5%B 、15%C 、25%D 、35%解析:根据上述公式,重组配子(换型)率为发生互换的精母细胞百分比的一半,即50%÷2=25% 练习2:基因型为 的精原细胞120个,其中若有30 个在形成精子的过程中发生了交换,在正常发育的情况下能产生亲本基因型和重组基因型的精子数目依次是 ( B )测交配子测交后代灰身长翅 黑身残翅 灰身残翅黑身长翅 灰身长翅黑身残翅F 1♀ ♂ (多数)42% 42%8% 8%×图二A B a bA 、480、480、60、60、B 、210、210、30、30、C 、90、90、30、30、D 、240、240、60、60解析:30个精原细胞发生了互换,产生四种精子为:BV :30、bv :30、Bv :30、bV :30。