数量遗传(QTL)定位的原理及研究进展
- 格式:ppt
- 大小:3.66 MB
- 文档页数:68

主要农作物QTL定位和克隆研究进展摘要:随着遗传图谱的日趋饱和QTL定位分析方法的日益完善,近年来农作物QTL的研究发展非常迅速。
本文首先从总体上对水稻、玉米、小麦、棉花、大豆这几个农作物的QTL定位作了简要的介绍,然后详细介绍了番茄、水稻、玉米被克隆出的几个QTL的克隆过程及基因功能,对整个QTL的分析方法作了系统的介绍。
关键词:QTL、定位、克隆、基因作物的许多农艺性状和经济性状是数量性状,研究作物数量性状的遗传对农作物育种具有十分重要的意义。
近几年,作物QTL定位和克隆发展迅速,本文详细阐述了主要农作物近几年的发展状况。
1.QTL定位研究进展QTL定位是检测分子标记和QTL间的连锁关系,估计QTL的效应利用分子标记进行遗传连锁分析,检测出QTL。
分子遗传学的发展和RFLP,RAPD,SSR,AFLP等分子标记技术的完善,加上日趋饱和的遗传图谱为工具和日益完善的QTL定位分析方法,已在许多作物上定位了不少QTL,并分析了各QTL的效应。
1.1 水稻水稻QTL的研究近年来发展非常迅速,已进行QTL定位的性状很多。
自Wang等[1]利用RFLP连锁图定位了水稻对稻瘟病有部分抗性的14个QTL以来,有关水稻QTL 定位的研究报道不断增加.目前,世界各国的科学家应用不同的群体,对水稻大多数性状进行了QTL定位,这些性状包括水稻的生育期、株高及其组成性状、产量及产量构成性状、谷粒外观品质、食味和营养品质等农艺性状、以及水稻种子的休眠性、水稻叶片叶绿素和过氧化氢含量等生理性状。
目前的数据表明水稻遗传图谱上的分子标记数已超过6000个,平均间距为75-100 kb,基本覆盖了水稻基因组的所有区域,为进一步精细定位及克隆提供了便利。
1.2玉米玉米的许多产量相关性状[2],如穗长、穗粗、行数、行粒数等,国内外都进行了较为深入的研究,定位了大量的QTL位点,并分析了它们的遗传规律,为玉米育种提供了很好的指导意义。

植物的大部分农艺性状、产量性状和品质性状属于数量遗传性状[1,2]。
数量遗传性状由多基因调控,在分离群体中表现为连续分布,且易受到环境影响。
数量遗传性状的深度解析与现代分子生物学技术的发展密切相关,分子标记、作图群体以及统计分析方法的应用和发展显著提高了数量遗传性状基因定位效率。
1分子标记分子标记反映不同个体间DNA 序列的变异,可以较为直观地反映DNA 水平的遗传多态性,具有共显性的特点[3]。
与利用表型进行目标性状筛选相比,分子标记具有不影响目标性状表达、不受环境影响、数量极多、能够对隐性遗传性状准确筛选、与基因变异直接相关、不与其他性状连锁等优点。
分子标记检测手段简单、迅速,因此作为作物遗传改良的重要工具广泛应用于遗传育种、基因挖掘、基因定位、基因库的设计与构建等方面。
根据其基础技术发展的过程,分子标记可分为3代:第1代:以分子杂交技术为基础的RFLP 标记。
DNA 序列改变时酶切位点会同时发生变化,从而产生RFLP 标记扩增条带的多态性。
RFLP 标记因其要求DNA 模板量大、分析过程繁琐、价格高昂、灵敏度不高等问题,目前逐渐被新兴分子标记所取代[4]。
摘要:解析数量遗传性状基因是作物遗传改良的重要手段。
分子生物学、各种组学和基因组测序技术的不断突破促进了分子育种技术的快速发展,分子标记技术也不断更新,逐渐成为数量遗传性状基因挖掘的重要方式之一。
简要介绍了数量遗传性状基因定位方法分子标记技术的发展历程和现状,并展望了分子标记技术的发展方向。
关键词:分子标记;数量性状;基因定位中图分类号:Q943.2文献标识码:A 文章编号:1008-1631(2021)05-0088-04收稿日期:2021-08-07基金项目:国家重点研发计划专项(2017YFD0300407);河北省农林科学院创新工程项目(2019-4-1B-5);河北省农林科学院科技创新人才队伍建设项目(C21R1302);科技部科技伙伴计划项目(KY202002003)作者简介:田玉(1988-),男,河北石家庄人,助理研究员,主要从事园艺作物栽培技术研究及示范推广工作。

数量性状的分子标记(定位的原理和方法讲义)作物中大多数重要的农艺性状和经济性状如产量、品质、生育期、抗逆性等都是数量性状。
与质量性状不同,数量性状受多基因控制,遗传基础复杂,且易受环境影响,表现为连续变异,表现型与基因型之间没有明确的对应关系。
因此,对数量性状的遗传研究十分困难。
长期以来,只能借助于数理统计的手段,将控制数量性状的多基因系统作为一个整体来研究,用平均值和方差来反映数量性状的遗传特征,无法了解单个基因的位置和效应。
这种状况制约了人们在育种中对数量性状的遗传操纵能力。
分子标记技术的出现,为深入研究数量性状的遗传基础提供了可能。
控制数量性状的基因在基因组中的位置称为数量性状基因座()。
利用分子标记进行遗传连锁分析,可以检测出,即定位()。
借助与连锁的分子标记,就能够在育种中对有关的的遗传动态进行跟踪,从而大大增强人们对数量性状的遗传操纵能力,提高育种中对数量性状优良基因型选择的准确性和预见性。
因此,定位是一项十分重要的基础研究工作。
年,等发表了第一篇应用连锁图在番茄中定位的论文。
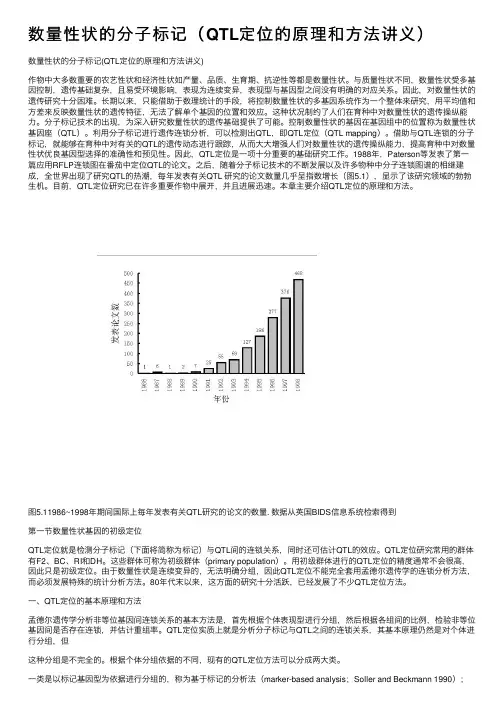
之后,随着分子标记技术的不断发展以及许多物种中分子连锁图谱的相继建成,全世界出现了研究的热潮,每年发表有关研究的论文数量几乎呈指数增长(图),显示了该研究领域的勃勃生机。
目前,定位研究已在许多重要作物中展开,并且进展迅速。
本章主要介绍定位的原理和方法。
图年期间国际上每年发表有关研究的论文的数量. 数据从英国信息系统检索得到第一节数量性状基因的初级定位定位就是检测分子标记(下面将简称为标记)与间的连锁关系,同时还可估计的效应。
定位研究常用的群体有、、和。
这些群体可称为初级群体()。
用初级群体进行的定位的精度通常不会很高,因此只是初级定位。
由于数量性状是连续变异的,无法明确分组,因此定位不能完全套用孟德尔遗传学的连锁分析方法,而必须发展特殊的统计分析方法。
年代末以来,这方面的研究十分活跃,已经发展了不少定位方法。
一、定位的基本原理和方法孟德尔遗传学分析非等位基因间连锁关系的基本方法是,首先根据个体表现型进行分组,然后根据各组间的比例,检验非等位基因间是否存在连锁,并估计重组率。

数量性状基因定位的原理及方法随着现代分子生物学的发展和分子标记技术的成熟,已经可以构建各种作物的分子标记连锁图谱。
基于作物的分子的标记连锁图谱,采用近年来发展的数量性状基因位点(QTL)的定位分析方法,可以估算数量性状的基因位点树目、位置和遗传效应。
本文介绍了数量性状基因定位的原理以及分析方法.每一种方法都有自己的优点,但也存在相应的缺陷。
1 数量性状基因定位的原理孟德尔遗传学分析非等位基因间连锁关系的基本方法是,首先根据个体表现型进行分组,然后根据各组间的比例,检验非等位基因间是否存在连锁,并估计重组率。
QTL定位实质上就是分析分子标记与QTL之间的连锁关系,其基本原理仍然是对个体进行分组,但这种分组是不完全的。
2 数量性状基因定位的方法自然界存在生物个体的性状、品质等多为数量性状,它们受多基因的控制,也易受环境影响。
.多基因及环境的共同作用结果使得数量性状表现为连续变异,基因型与表现型间的对应关系也难以确定.因此,长期以来,科学工作者只是借助数理统计方法,将复杂的多基因系统作为一个整体,用平均值和方差来表示数量性状的遗传特征,而对单个基因的效应及位置、基因间的相互作用等无法深入了解,从而限制了育种中数量性状的遗传操作能力。
20 世纪80 年代以来发展的分子标记技术为深入研究数量性状的遗传规律及其操作创造了条件, 提高了植物育种中目标数量性状优良基因型选择的可能性、准确性及预见性。
下面主要介绍了几种定位方法。
2。
1 QTL 定位方法连锁是QTL定位的遗传基础.QTL 定位是通过数量性状观察值与标记间的关联分析,即当标记与特定性状连锁时,不同标记基因型个体的表型值存在显著差异,来确定各个数量性状位点在染色体上的位置、效应,甚至各个QTL 间的相关作用。
因此,QTL 定位实质上也就是基于一个特定模型的遗传假设,是统计学上的一个概念,有可信度(如99%,95%等),与数量性状基因有本质区别(图1)。



植物分子生物技术—QTL定位的研究摘要:分子标记技术和数量遗传学的发展,使得分子遗传学与数量遗传学相互渗透和融合,从而形成了一个新的研究领域—分子数量遗传学(Molecular Quantitative Genetics)。
分子数量遗传学研究的内容,就是借助分子标记,采用适当的统计分析方法明确QTL 在染色体上的位置及其效应。
而QTL 定位的原理是:利用适当的分离群体,构建较高密度的、分布较均匀的、覆盖全基因组的分子标记连锁图。
根据遗传连锁的基本遗传学原理,对分离群体中单株的标记基因型和性状的表型值进行一定的统计分析,将决定数量性状的QTL 定位在分子标记连锁图中。
目前,QTL 定位的方法主要有单标记分析法(Edwards et al, 1987),区间作图法(Lander and Botstein, 1989)和复合区间作图法(Zeng, 1994)等。
关键词: QTL 植物分子生物技术QTL 定位就是采用类似单基因定位的方法将QTL定位在遗传图谱上,确定QTL与遗传标记间的距离( 重组率表示)。
根据标记数目的不同,可分为单标记、双标记和多标记几种方法。
根据统计分析方法的不同,可分为方差与均值分析法、回归及相关分析法、矩估计及最大似然法等根据标记区问数可分为零区间作图、单区间作图和多区间作图。
此外,还有将不同方法结合起来的缘音分析方法,如QTL复合区间作图(CIM)、多区问作图(MM)、多QTL作图、多性状作图(MTM)等等。
建立在标记与数量性状之间相互关联基础上的关联分析方法主要有两类:1)以标记为基础(MarkerBased)的平均值差异法(简称MB法);2)以性状为基础(Trait—Based method)的方法(简称TB法) 许多学者根据不同群体的遗传特性,分别提出了相应的标记座位与QTL 相互关联的检测方法,所涉及到的群体包括;近交系闸分离群体,异交系间分离群体,加倍单倍体(DH)群体,两个近交系间的重组近变系(RIL),一粒传群体(SSD),单倍体群体等。

数量性状基因定位的原理及方法随着现代分子生物学的发展和分子标记技术的成熟,已经可以构建各种作物的分子标记连锁图谱.基于作物的分子的标记连锁图谱,采用近年来发展的数量性状基因位点(QTL)的定位分析方法,可以估算数量性状的基因位点树目、位置和遗传效应。
本文介绍了数量性状基因定位的原理以及分析方法。
每一种方法都有自己的优点,但也存在相应的缺陷。
1 数量性状基因定位的原理孟德尔遗传学分析非等位基因间连锁关系的基本方法是,首先根据个体表现型进行分组,然后根据各组间的比例,检验非等位基因间是否存在连锁,并估计重组率。
QTL定位实质上就是分析分子标记与QTL之间的连锁关系,其基本原理仍然是对个体进行分组,但这种分组是不完全的。
2 数量性状基因定位的方法自然界存在生物个体的性状、品质等多为数量性状,它们受多基因的控制,也易受环境影响。
多基因及环境的共同作用结果使得数量性状表现为连续变异,基因型与表现型间的对应关系也难以确定。
因此,长期以来,科学工作者只是借助数理统计方法,将复杂的多基因系统作为一个整体,用平均值和方差来表示数量性状的遗传特征,而对单个基因的效应及位置、基因间的相互作用等无法深入了解, 从而限制了育种中数量性状的遗传操作能力。
20 世纪80 年代以来发展的分子标记技术为深入研究数量性状的遗传规律及其操作创造了条件, 提高了植物育种中目标数量性状优良基因型选择的可能性、准确性及预见性。
下面主要介绍了几种定位方法。
2.1 QTL 定位方法连锁是QTL定位的遗传基础。
QTL 定位是通过数量性状观察值与标记间的关联分析,即当标记与特定性状连锁时,不同标记基因型个体的表型值存在显著差异,来确定各个数量性状位点在染色体上的位置、效应, 甚至各个QTL 间的相关作用。
因此, QTL 定位实质上也就是基于一个特定模型的遗传假设, 是统计学上的一个概念, 有可信度(如99% , 95%等) ,与数量性状基因有本质区别( 图1)。
数量性状的分⼦标记(QTL定位的原理和⽅法讲义)数量性状的分⼦标记(QTL定位的原理和⽅法讲义)作物中⼤多数重要的农艺性状和经济性状如产量、品质、⽣育期、抗逆性等都是数量性状。
与质量性状不同,数量性状受多基因控制,遗传基础复杂,且易受环境影响,表现为连续变异,表现型与基因型之间没有明确的对应关系。
因此,对数量性状的遗传研究⼗分困难。
长期以来,只能借助于数理统计的⼿段,将控制数量性状的多基因系统作为⼀个整体来研究,⽤平均值和⽅差来反映数量性状的遗传特征,⽆法了解单个基因的位置和效应。
这种状况制约了⼈们在育种中对数量性状的遗传操纵能⼒。
分⼦标记技术的出现,为深⼊研究数量性状的遗传基础提供了可能。
控制数量性状的基因在基因组中的位置称为数量性状基因座(QTL)。
利⽤分⼦标记进⾏遗传连锁分析,可以检测出QTL,即QTL定位(QTL mapping)。
借助与QTL连锁的分⼦标记,就能够在育种中对有关的QTL的遗传动态进⾏跟踪,从⽽⼤⼤增强⼈们对数量性状的遗传操纵能⼒,提⾼育种中对数量性状优良基因型选择的准确性和预见性。
因此,QTL定位是⼀项⼗分重要的基础研究⼯作。
1988年,Paterson等发表了第⼀篇应⽤RFLP连锁图在番茄中定位QTL的论⽂。
之后,随着分⼦标记技术的不断发展以及许多物种中分⼦连锁图谱的相继建成,全世界出现了研究QTL的热潮,每年发表有关QTL 研究的论⽂数量⼏乎呈指数增长(图5.1),显⽰了该研究领域的勃勃⽣机。
⽬前,QTL定位研究已在许多重要作物中展开,并且进展迅速。
本章主要介绍QTL定位的原理和⽅法。
图5.11986~1998年期间国际上每年发表有关QTL研究的论⽂的数量. 数据从英国BIDS信息系统检索得到第⼀节数量性状基因的初级定位QTL定位就是检测分⼦标记(下⾯将简称为标记)与QTL间的连锁关系,同时还可估计QTL的效应。
QTL定位研究常⽤的群体有F2、BC、RI和DH。
这些群体可称为初级群体(primary population)。

数量性状基因座(QTL)精细定位的研究进展来源:卢超的日志本文转自博士论坛,希望与大家分享!生物界的许多性状是以数量性状为基础的,该类性状的发生不是决定于一对等位基因,而是受到两对或更多对等位基因的控制,每对等位基因彼此之间没有显性与隐性的区别,而是共显性。
这些等位基因对该遗传性状形成的作用微小,所以也称为微效基因(minor gen e),它们在染色体上的位置称为数量性状基因座(quantitative trait loci, QTL)。
数量性状就是由许多对微效基因的联合效应造成的一类具有正态分布特性的性状,具有这种性状的个体在正态分布中的位置决定于它们所具有的微效基因的多少。
数量性状之所以复杂,是因为这种性状与基因型之间没有一一对应的关系,并且环境因素对它有显着的影响[1]。
1961年,Thoday首次利用一对侧翼标记定位一个QTL[2],随后QTL的研究得到飞速发展。
现已证明,只要是控制连续分布性状的数量性状基因座,就能在实验群体或远交群体中定位[3] 。
在整个QTL的研究中,可分为发现QTL,QTL的定位估计和精细定位[4]。
Glazier认为可分成四个步骤:连锁和关联分析,精细定位,序列分析和候选基因的功能检测[5]。
在模式生物小鼠的QTL研究,已描述的实验步骤更为详细[6]:第一,把QTL定位到染色体的区段上。
典型的QTL定位方法须对几百个杂交后代进行恰当的表型检测;利用覆盖整个基因组的75-100个遗传标记进行基因组扫描,遗传标记的平均跨度为15-20个厘摩(cM);准确基因分型,然后对性状和遗传标记之间进行连锁和关联分析;计算QTL与某标记位点靠近的可能性大小。
这种方法即使经过成百上千次的表型和基因型的分析,定位的QTL在染色体上还是一个较大的区间。
第二,把一个QTL的作用与其它QTL的作用分开。
通常在同源系(congenic strain)中进行,这样就把多基因遗传的性状转化成单基因性状进行研究。

qtl原理QTL原理。
QTL(Quantitative Trait Loci)是指定位于染色体上的控制数量性状的基因或基因组区域。
数量性状是指受多个基因和环境因素影响的性状,如生长速率、产量等。
QTL分析是一种通过遗传连锁和相关分析,来确定数量性状的遗传基础的方法。
QTL的发现对于理解数量性状的遗传机制、育种改良和基因定位都具有重要意义。
QTL分析的基本原理是通过构建遗传连锁图谱,确定数量性状与分子标记之间的遗传关系。
首先,需要构建一个包含足够密度的分子标记的遗传图谱,这些分子标记可以是单核苷酸多态性(SNP)、简单重复序列(SSR)等。
然后,通过测定不同个体的数量性状表型值,结合分子标记的基因型数据,利用统计方法来分析数量性状与分子标记之间的遗传关系。
最终确定数量性状的QTL位置和效应大小。
QTL分析的关键是构建遗传图谱和选择合适的统计方法。
构建遗传图谱需要选择合适的分子标记,保证标记的分布均匀和密度足够。
同时,统计方法的选择也是至关重要的,常用的方法包括单因素方差分析、双因素方差分析、QTL定位等。
QTL分析的应用范围非常广泛,涉及到农业、医学、动植物育种等多个领域。
在农业领域,QTL分析可以帮助育种者快速筛选出具有优良数量性状的种质资源,加速育种进程。
在医学领域,QTL分析可以帮助研究者发现与疾病相关的基因,为疾病的预防和治疗提供理论依据。
在动植物育种领域,QTL分析可以帮助育种者理解数量性状的遗传机制,指导育种方向。
总之,QTL分析是一种重要的遗传分析方法,通过构建遗传图谱和统计分析,可以确定数量性状的遗传基础,为育种改良和基因定位提供重要依据。
随着分子标记技术和统计方法的不断发展,QTL分析将在更多领域发挥重要作用,推动遗传学研究和应用的进步。
猪QTL定位研究进展罗琰(华中农业大学动物科技学院,湖北武汉 430070)摘要:数量性状位点(QTL)的定位是鉴别家畜控制重要经济性状基因的第一步。
人类基因组计划和分子生物学技术的飞速发展及其应用,推动了猪基因图谱的研究进展,使猪的QTL定位研究取得长足的进展,从而为利用分子标记进行数量性状位点的鉴别和定位奠定了坚实的基础。
本文概述了影响猪繁殖、生长、肉质、胴体等主要经济性状的QTLs定位研究进展情况。
关键词:猪;QTL;定位;研究进展猪是人类最早驯养的动物之一,具有重要的经济价值,且其经济性状多受数量性状位点(quantitative trait loci,QTL)控制,因而QTL的研究已成为国际上的研究热点,并已取得了一定的成果。
目前,对猪数量性状的研究已发展为直接将研究目标指向各个数量性状位点,借助各种分子遗传标记,构建遗传连锁图谱,通过一定的统计分析方法,从而将影响数量性状的多个基因剖分开来,将其定位于特定的染色体区域,确定它们与其它基因的关系,最终分析出其DNA序列。
现今,已经检测出了与猪各种性状相关的4928个QTL,为猪的品种改良和数量性状主效基因的检测及应用研究提供了重要的参考依据和前提条件。
1 QTL及其定位QTL的概念是由Geldermann(1975年)最先提出的,是指数量性状的基因在基因组中的位置,是对某一数量性状有一定决定作用的单个基因或微效多基因簇[1]。
猪的生长速度、瘦肉率、采食量、肉质性状、产仔率和抗病性等大多数经济性状受数量性状基因座控制。
目前,猪的应激敏感基因、高产仔数基因、瘦肉率和脂肪率座位等具有重要经济价值的基因已被定位或发现了与其连锁的DNA标记,其中部分已定位的基因,在育种中发挥着重要的作用,展现出基因定位和遗传图谱研究的价值。
虽然分子标记取得了许多进展,但尚处于初始阶段,QTL的特点是多基因的微效性,且在染色体上存在着数以百计甚至千计的QTL,已显出很好的发展前景。