线粒体与叶绿体的来源
- 格式:pptx
- 大小:1.02 MB
- 文档页数:13

简述线粒体和叶绿体内共生起源的主要论据一、线粒体的共生起源线粒体是细胞内的一种器官,负责细胞的能量代谢和ATP的合成。
线粒体内含有自己的DNA和蛋白质,与细胞核不同。
线粒体的共生起源,是指线粒体和原核细胞的共同进化过程。
1.线粒体具有自主繁殖能力线粒体具有自主繁殖能力,通过二分裂方式繁殖。
这表明线粒体并非完全依赖于细胞核的支配。
2.线粒体和细菌有相似之处线粒体和细菌有许多相似之处,都具有类似的细胞壁、膜结构和基因组成等。
并且,线粒体在结构和生化反应方面也与细菌有许多相似之处。
3.线粒体和原核细胞有共同祖先线粒体的DNA序列与细菌的DNA序列有相似之处,这表明线粒体和细菌有共同的祖先。
这也支持了线粒体的共生起源理论。
二、叶绿体的共生起源叶绿体是植物和藻类细胞内的一种器官,负责光合作用和氧气释放。
叶绿体也含有自己的DNA和蛋白质,与细胞核不同。
叶绿体的共生起源,是指叶绿体和细胞核的共同进化过程。
1.叶绿体与细菌有相似之处叶绿体与细菌有相似之处,如细胞壁结构、膜结构和基因组成等。
并且,叶绿体内含有一种称为“类囊体”的结构,与细菌内的类囊体结构相似。
2.叶绿体具有自主繁殖能力叶绿体具有自主繁殖能力,通过二分裂方式繁殖。
这表明叶绿体并非完全依赖于细胞核的支配。
3.叶绿体和藻类有共同祖先叶绿体和藻类有共同的祖先,这表明叶绿体的起源和藻类的共生起源有关。
综上所述,线粒体和叶绿体的共生起源理论,是通过对细胞结构、生化反应、遗传物质等方面的比较研究,提出并支持的。
这些论据都表明,线粒体和叶绿体都具有自主繁殖能力、与细菌具有相似之处、与其宿主细胞有共同祖先等,这些都为线粒体和叶绿体的共生起源提供了有力的证据。

例析线粒体和叶绿体的内共生起源学说封开县江口中学陶勇由于线粒体和叶绿体具有独特的半自主性并与细胞核建立了复杂而协调的互作关系,它们的起源一直以来多被认为有别于其他细胞器。
在人们为这两种细胞器设计的起源假说中,内共生起源学说很好地贴合了线粒体和叶绿体的半自主性和核质关系特征,因而得到了广泛的认可和支持。
内共生起源学说认为,线粒体和叶绿体分别起源于原始真核细胞内共生的行有氧呼吸的细菌和行光能自养的蓝细菌。
该假说的提出远早于mtDNA和cpDNA的发现。
随着人们对真核细胞超微结构、线粒体和叶绿体DNA及其编码机制的认识,内共生起源学说的内涵得到了进一步充实。
1970年,Margulis在已有的资料基础上提出了一种更为细致的设想。
假设认为,真核细胞的祖先是一种体积较大、不需氧具有吞噬能力的细胞,通过糖酵解获取能量。
而线粒体的祖先则是一种革兰氏阴性菌,具备三羧酸循环所需的酶和电子传递链系统,可利用氧气把糖酵解的产物丙酮酸进一步分解,获得比糖酵解更多的能量。
当这种细菌被原始真核细胞吞噬后,即与宿主细胞间形成互利的共生关系:原始真核细胞利用这种细菌获得更充分的能量;而这种细菌则从宿主细胞获得更适宜的生存环境。
与此类似,叶绿体的祖先可能是原核生物的蓝细菌(cyanbacteria)。
当这种蓝细菌被原始真核细胞摄人后,为宿主细胞进行光合作用;而宿主细胞则为其提供其他的生存条件。
线粒体和叶绿体的内共生学说先后得到了大量的生物学研究证据的支持。
特别是近期的分子生物学和生物信息学的研究发现真核细胞的细胞核中存在大量原本可能属于呼吸细菌或蓝细菌的遗传信息,说明最初的呼吸细菌和蓝细菌的大部分基因组在漫长的共进化过程中发生了向细胞核的转移。
这种转移极大的削弱了线粒体和叶绿体的自主性,建立起稳定、协调的核质互作关系。
一、线粒体和叶绿体内共生起源学说的主要论据1.基因组与细菌基因组具有明显的相似性线粒体和叶绿体具有细菌基因组的典型特征。

例析线粒体和叶绿体的内共生起源学说————————————————————————————————作者:————————————————————————————————日期:ﻩ例析线粒体和叶绿体的内共生起源学说封开县江口中学陶勇由于线粒体和叶绿体具有独特的半自主性并与细胞核建立了复杂而协调的互作关系,它们的起源一直以来多被认为有别于其他细胞器。
在人们为这两种细胞器设计的起源假说中,内共生起源学说很好地贴合了线粒体和叶绿体的半自主性和核质关系特征,因而得到了广泛的认可和支持。
内共生起源学说认为,线粒体和叶绿体分别起源于原始真核细胞内共生的行有氧呼吸的细菌和行光能自养的蓝细菌。
该假说的提出远早于mtDNA和cpDNA的发现。
随着人们对真核细胞超微结构、线粒体和叶绿体DNA及其编码机制的认识,内共生起源学说的内涵得到了进一步充实。
1970年,Margulis在已有的资料基础上提出了一种更为细致的设想。
假设认为,真核细胞的祖先是一种体积较大、不需氧具有吞噬能力的细胞,通过糖酵解获取能量。
而线粒体的祖先则是一种革兰氏阴性菌,具备三羧酸循环所需的酶和电子传递链系统,可利用氧气把糖酵解的产物丙酮酸进一步分解,获得比糖酵解更多的能量。
当这种细菌被原始真核细胞吞噬后,即与宿主细胞间形成互利的共生关系:原始真核细胞利用这种细菌获得更充分的能量;而这种细菌则从宿主细胞获得更适宜的生存环境。
与此类似,叶绿体的祖先可能是原核生物的蓝细菌(cyanbacteria)。
当这种蓝细菌被原始真核细胞摄人后,为宿主细胞进行光合作用;而宿主细胞则为其提供其他的生存条件。
线粒体和叶绿体的内共生学说先后得到了大量的生物学研究证据的支持。
特别是近期的分子生物学和生物信息学的研究发现真核细胞的细胞核中存在大量原本可能属于呼吸细菌或蓝细菌的遗传信息,说明最初的呼吸细菌和蓝细菌的大部分基因组在漫长的共进化过程中发生了向细胞核的转移。
这种转移极大的削弱了线粒体和叶绿体的自主性,建立起稳定、协调的核质互作关系。

细胞生物学第七章线粒体与叶绿体知识点整理线粒体和叶绿体是细胞中两个重要的细胞器。
它们在细胞代谢和能量转换中发挥着重要的作用。
以下是关于线粒体和叶绿体的一些重要知识点:线粒体:1.结构:线粒体是一个由两层膜包围的细胞器。
它包含一个外膜和一个内膜,内膜形成了许多内突起,称为线粒体内膜嵴。
2.能量转换:线粒体是细胞中的能量生产中心。
它通过细胞呼吸过程中的氧化磷酸化来产生能量,将食物分子中的化学能转化为细胞可以使用的三磷酸腺苷(ATP)。
3. 基因组:线粒体具有自己的基因组,称为线粒体DNA(mtDNA)。
它主要编码细胞呼吸过程中所需的蛋白质。
mtDNA由母亲遗传给子代,因此线粒体DNA有助于研究人类的遗传和进化。
4.线粒体疾病:线粒体功能障碍可以导致许多疾病,如线粒体脑肌病、线粒体糖尿病和阿尔茨海默病。
这些疾病通常会影响能量的产生和细胞的正常功能。
叶绿体:1.结构:叶绿体是植物和一些原生生物中的细胞器。
它也是由两层膜包围,并且内膜形成了一系列叫做叶绿体嵴的结构。
2.光合作用:叶绿体是光合作用的主要场所,其中光能转化为化学能以供细胞使用。
叶绿体中的叶绿素能够吸收太阳能,并将其转化为光合作用的产物,如葡萄糖。
3. 基因组:叶绿体也具有自己的基因组,称为叶绿体DNA(cpDNA)。
它主要编码参与光合作用和叶绿体功能的蛋白质。
4.叶绿体疾病:类似于线粒体疾病,叶绿体功能障碍也会导致一系列疾病,在植物中称为叶绿体遗传病。
这些疾病通常会导致叶绿体的正常结构和功能受损。
1.起源:线粒体起源于古代原核生物,而叶绿体起源于古代蓝藻细菌。
这些细菌进化成为现代细胞中的线粒体和叶绿体。
2.功能:线粒体主要参与能量转换,而叶绿体主要参与光合作用。
它们在细胞代谢中的角色不同,但都与能量生产和细胞功能密切相关。
3.基因组:线粒体和叶绿体都有自己的基因组,具有其中一种程度的自主复制和表达能力。
不过,线粒体基因组比较小,叶绿体基因组比较大。

线粒体和叶绿体的起源和演化历程生命起源于多样的地方,而人类总是把目光聚焦于其中一种生命形式——生物。
生物有初生的细胞,也有千变万化的种类。
其中,线粒体和叶绿体就是群体中经常会被提及的细胞器。
一、线粒体的起源线粒体是一个具有独立膜结构的小细胞器。
科学家们在研究中发现,线粒体有独立、自主、复制和分裂的能力,又有自主的生物合成过程。
由此可见,线粒体和细胞是两个完全独立的实体。
现在,DNA进入线粒体中就能在里面进行复制、转录和翻译。
许多研究表明,线粒体有良好的远程活动性,并可以积极地参与细胞的代谢和能量生产。
线粒体的形态结构显示出一个功能复杂、高度耐受和实用的细胞器。
最初,科学家们普遍认为线粒体起源于细胞外侵入,但太阳能生成机理的发现揭示了线粒体的更为绝妙的起源。
现在,焦炭沉积导致的自然化学分析表明,早期的生命形式最早起源于化学环境中的光合作用,并赖以生存。
自繁殖和依附机体的形态再次出现,这意味着早期的生物形式已经具备了完整的生命活动。
但是,回到线粒体的问题上,它是否可以类比现代光合作用的调控保留下来呢?二、叶绿体的起源叶绿体同样是一个细胞器,具有独立膜壳和固有的DNA,能够参与自主的生物合成活动。
它的主要功能就是接收太阳能,进行光合作用,将其中的光能转化为化学能,为细胞提供生命所需的物质和能量。
早期的研究表明,叶绿体起源于一种原初的光合作用菌类——兰氏菌(Cyanobacteria),这个研究理论目前也被大多数科学家所认同。
兰氏菌的DNA已被分析出来,它的基因与叶绿体有着非常相似的序列,这也进一步证明了叶绿体起源自一个内共生过程。
这个内共生过程发生在生命起源的早期,或者是早期的环境中,它与线粒体的起源过程类似。
现在,科学家对这个过程及其演化道路和机制已有了比较清晰的认识。
不论生命进化的方向是否符合我们的预期,能够形成生命、维护生命和传承生命的原则和规律似乎都是丝毫不变的。
三、线粒体和叶绿体的演化历程线粒体和叶绿体都经历过一个演化过程,这个演化过程跨越了数百万年的时间,是由朴素的单细胞生物向越来越复杂的多细胞生物的过渡。

泡桐叶绿体和线粒体的提取与分离一、试验目的:1、将泡桐叶片中的叶绿体和线粒体提取出来。
2、掌握植物叶片中叶绿体和线粒体提取与分离的基本技术。
3、学习蔗糖沉淀差速离心法和差速离心法提取线粒体的方法。
二、实验材料与仪器:实验材料:泡桐新鲜叶片。
实验试剂:蒸馏水;液氮;缓冲液A(50 mmo l/L Tr is,,25 mmo l /L EDTA,1.25 mo l/L N aC l ,10 mmo l/Lβ-巯基乙醇, ψ=0. 1% (W /V ) BSA (牛血清蛋白) , ψ= 2% (W /V) CTAB, 10 g /L PVP, pH = 8. 0);缓冲液B (Tris2HCl 0.105 mol/ L, 蔗糖0.15 mol/ L, EDTA 0.1005 mol/ L, BSA 0.11% , 巯基乙醇0.11%, pH =7.15);缓冲液C( Tris2HCl 0.105 mol/ L, 蔗糖0.13 mol/L, MgCl2 0.101 mol/ L, pH 7.15);缓冲液D( 0. 2 mol/ L 蔗糖、0. 3mol/ L 甘露醇、50 mol/ L Tris2Cl、1 mol/ L EDTA、0. 1% BSA 、0. 6% PVP、0. 1% B2巯基乙醇,pH 7. 5);缓冲液E( 蔗糖0. 3 mol/ L、50mol/ L T ris2Cl, pH 7. 5);缓冲液F( 50 mol/ LTris2Cl、0. 3 mol/ L 甘露醇、0. 2 mol/ L 蔗糖、0. 1% BSA、0. 6% PVP, pH = 7. 5);纱布等。
试验仪器:研钵,研磨棒;离心管(250ml);高速冷冻离心机;移液枪;高速组织捣碎器等三、试验步骤:1、叶绿体的提取与分离(1) 称取新鲜泡桐叶片50 ~ 100 g, , 暗处理12小时,蒸馏水洗干净在液氮中保存3 h以上, 以最快速度磨成粉末, 然后用6层无菌纱布过滤, 收集滤液, 弃渣。
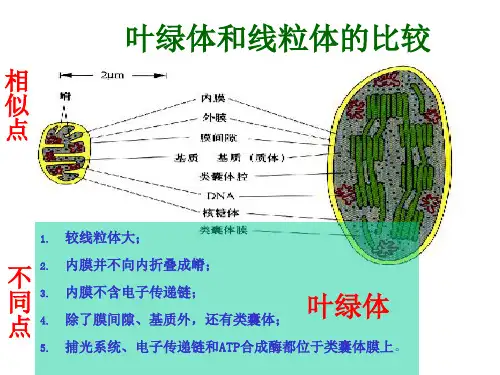
高考专题复习:线粒体和叶绿体的内共生起源学说由于线粒体和叶绿体具有独特的半自主性并与细胞核建立了复杂而协调的互作关系,它们的起源一直以来多被认为有别于其他细胞器。
在人们为这两种细胞器设计的起源假说中,内共生起源学说很好地贴合了线粒体和叶绿体的半自主性和核质关系特征,因而得到了广泛的认可和支持。
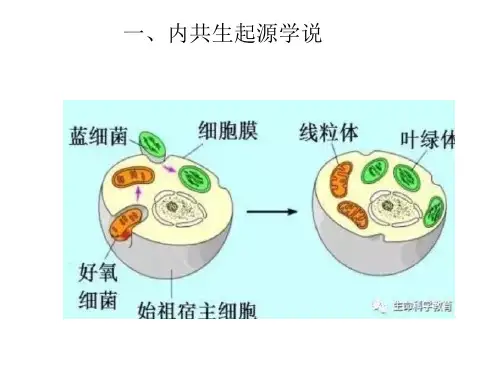
(2019年广州一模第29题第3问)内共生起源学说:认为线粒体和叶绿体分别起源于原始真核cell内共生的细菌和蓝藻。
线粒体来源于细菌,即细菌被真核生物吞噬后,在长期共生过程中,通过演变,形成了线粒体。
叶绿体来源于蓝藻,被原始真核cell摄入胞内,在共生关系中,形成了叶绿体。
(纤毛虫(Nassula ornata)与藻类的共生现象)主要论据:①线粒体和叶绿体的基因组在大小、形态和结构方面与细菌的相似。
②线粒体核叶绿体有自己完整的蛋白质合成系统,能独立合成蛋白质。
③线粒体和叶绿体的两层被膜有不同的进化来源,外膜与内膜的结构和成分差异很大。
④线粒体和叶绿体能以分裂的方式进行繁殖,这与细菌的繁殖方式类似。
⑤线粒体和叶绿体能在异源细胞内长期生存。
⑥线粒体的祖先很可能来自反硝化副球菌或紫色非硫光合细菌。
⑦发现介于包内共生蓝藻与叶绿体之间的结构---蓝小体,其特征在很多方面可作为原始蓝藻向叶绿体演化的佐证。
不足之处:a.从进化角度:如此解释在代谢上明显占优势的共生体反而将大量的遗传信息,转移到宿主cell中,不能解释细胞核是如何进化来的,即原核cell如何演化为真核cell。
b.线粒体和叶绿体的基因组中存在内含子,而真细菌原核生物基因组中不含有内含子,不能解释其内含子从何而来。
香梅说:蓝藻可以进行有氧呼吸,为什么植物还需要线粒体? 欢迎各位参与讨论!例1:生物学家提出了“线粒体是起源于好氧细菌”的假说。
该假说认为,在进化过程中原始真核细胞吞噬了某种好氧细菌形成共生关系,最终被吞噬的好氧细菌演化成线粒体。
下列多个事实中无法支持该假说的是()A.哺乳动物细胞的核DNA由雌雄双亲提供,而线粒体DNA则主要来自雌性亲本B.线粒体外膜的成分与真核细胞的细胞膜相似,而内膜则同现存细菌的细胞膜相似C.高等植物细胞的核DNA与蛋白质结合呈线状,而线粒体DNA裸露且主要呈环状D.真核细胞中有功能不同的多种细胞器,而线粒体中仅存在与细菌中类似的核糖体例2.线粒体起源的内共生学说认为,原始真核细胞吞噬了能进行有氧呼吸的原始细菌,它们之间逐渐形成了互利共生关系,最终原始细菌演变成线粒体。

内共生起源学说是指叶绿体和线粒体起源于原核生物与细胞质共生的假说。
这一学说在生物学领域引起了广泛的关注和讨论,科学家们通过一系列的研究和实验证据,逐渐揭示了叶绿体和线粒体相似点的证据。
1. 细胞生物学研究通过对细胞结构和功能的研究发现,叶绿体和线粒体与细胞质内的原核生物有着明显的相似点。
叶绿体和一些细菌在形态结构上有着相似之处,都具有双层膜结构和一定的自主遗传物质。
线粒体则在形态上与一些真核细胞内的细菌有着相似之处,都具有类似的内膜结构和呼吸链。
2. 生物进化研究通过对叶绿体和线粒体DNA的比较分析发现,它们的遗传物质与一些原核细菌的DNA有着明显的相似性。
这种相似性不仅表现在基因序列上,还表现在某些基因的功能和调控方式上,这为内共生起源学说提供了强有力的证据。
3. 分子生物学研究通过对叶绿体和线粒体的蛋白质结构和功能进行研究发现,它们与一些原核生物的蛋白质具有着相似的结构和功能。
线粒体的氧化磷酸化过程与一些细菌的呼吸链有着相似的机制,叶绿体的光合作用过程与一些细菌的光合作用也有着相似的机制。
4. 生物地理学研究通过对不同物种间叶绿体和线粒体遗传物质的比较发现,它们在物种间的分布和进化历程上有着一定的规律性。
这些规律性与内共生起源学说所预测的叶绿体和线粒体起源于共生原核生物的假说吻合,为该学说提供了地理学上的证据。
通过对细胞生物学、生物进化、分子生物学和生物地理学等多个领域的研究,科学家们发现了大量的证据支持内共生起源学说,揭示了叶绿体和线粒体的相似点。
这些证据不仅深化了我们对生命起源和进化的理解,也为我们认识生物多样性和生态系统的演化历程提供了重要的参考依据。
在未来的研究中,科学家们将继续探索叶绿体和线粒体的起源和进化,推动生物学领域的进一步发展。
在探索内共生起源学说的证据的过程中,科学家们不仅仅局限于叶绿体和线粒体在细胞学、生物进化、分子生物学和生物地理学领域的研究,还通过生化学、遗传学、比较基因组学等多个层面的研究,进一步扩大了对叶绿体和线粒体相似点的认知。

细胞内重要的能量转换器——线粒体和叶绿体真核细胞就像一座复杂的工厂,工厂的内部被分成许多不同的车间,这些车间就是细胞内的各种细胞结构,这些车间各自行使着不同的功能,使得整个细胞有条不紊地进行复杂的生命活动。
这些车间中,有两个重要的能量转换场所,它们就是线粒体和叶绿体。
线粒体是真核生物生命活动所需能量的主要产生场所,被誉为“细胞的动力车间”,没有了线粒体,细胞或生物体的生命就将终结。
叶绿体是大多数植物进行光合作用的场所,被誉为“细胞的养料制造车间”,光能是生物界赖以生存的最根本的能量来源,绿色植物通过光合作用,利用光能将CO2和H2O合成为有机物,这些有机物不仅为植物自身所用,动物和微生物也要直接或间接以之为食,因此叶绿体对整个生物界都有重要作用。
线粒体和叶绿体在外观和构造上都有很多相似的地方,但它们所行使的功能却存在着很大的区别,要弄清线粒体和叶绿体的功能具有很大差别的原因,就必须从它们的亚显微结构入手。
一.叶绿体和线粒体的膜叶绿体和线粒体结构上的相同点之一就是它们都具有双层膜结构,这两层膜和细胞膜一样,都由磷脂双分子层作为基本支架,其上结合各种蛋白质分子,具有一定的流动性,在物质运输方面也都有选择透过性。
叶绿体除了含有表面的两层膜外,其内部的囊状结构也是由一层膜围成的,囊状结构膜也具有上述特点。
但是如果进一步分析这些膜上的各成分的含量,尤其是蛋白质的含量,就不难发现其中的差异。
相关的研究结果如下表:为什么会出现上述结果呢?我们知道蛋白质是生命活动的主要承担者,细胞内的各种膜要行使其功能,也离不开蛋白质。
线粒体内膜上蛋白质含量高,原因是线粒体进行有氧呼吸所需要的各种酶有很多都位于其内膜上,而其外膜并不直接参与有氧呼吸。
叶绿体的双层膜蛋白质含量都很低,也是因为它们并不直接参与光合作用,而囊状结构膜却是光合作用的重要场所,其上含有大量与光合作用有关的酶。
通过上面的分析可以看出,叶绿体和线粒体的各种膜,由于它们的结构组成不同,功能也就不同。
例析线粒体和叶绿体的共生起源学说封开县江口中学勇由于线粒体和叶绿体具有独特的半自主性并与细胞核建立了复杂而协调的互作关系,它们的起源一直以来多被认为有别于其他细胞器。
在人们为这两种细胞器设计的起源假说中,共生起源学说很好地贴合了线粒体和叶绿体的半自主性和核质关系特征,因而得到了广泛的认可和支持。
共生起源学说认为,线粒体和叶绿体分别起源于原始真核细胞共生的行有氧呼吸的细菌和行光能自养的蓝细菌。
该假说的提出远早于mtDNA和cpDNA的发现。
随着人们对真核细胞超微结构、线粒体和叶绿体DNA及其编码机制的认识,共生起源学说的涵得到了进一步充实。
1970年,Margulis在已有的资料基础上提出了一种更为细致的设想。
假设认为,真核细胞的祖先是一种体积较大、不需氧具有吞噬能力的细胞,通过糖酵解获取能量。
而线粒体的祖先则是一种革兰氏阴性菌,具备三羧酸循环所需的酶和电子传递链系统,可利用氧气把糖酵解的产物丙酮酸进一步分解,获得比糖酵解更多的能量。
当这种细菌被原始真核细胞吞噬后,即与宿主细胞间形成互利的共生关系:原始真核细胞利用这种细菌获得更充分的能量;而这种细菌则从宿主细胞获得更适宜的生存环境。
与此类似,叶绿体的祖先可能是原核生物的蓝细菌(cyanbacteria)。
当这种蓝细菌被原始真核细胞摄人后,为宿主细胞进行光合作用;而宿主细胞则为其提供其他的生存条件。
线粒体和叶绿体的共生学说先后得到了大量的生物学研究证据的支持。
特别是近期的分子生物学和生物信息学的研究发现真核细胞的细胞核中存在大量原本可能属于呼吸细菌或蓝细菌的遗传信息,说明最初的呼吸细菌和蓝细菌的大部分基因组在漫长的共进化过程中发生了向细胞核的转移。
这种转移极大的削弱了线粒体和叶绿体的自主性,建立起稳定、协调的核质互作关系。
一、线粒体和叶绿体共生起源学说的主要论据1.基因组与细菌基因组具有明显的相似性线粒体和叶绿体具有细菌基因组的典型特征。
它们均为单条环状双链DNA分子,不含5-甲基胞嘧啶,无组蛋白结合并能进行独立的复制和转录。
为什么说线粒体和叶绿体是半自主遗传细胞器?
因为线粒体和叶绿体中含有质体,它们是一些环状DNA,可以编码一些蛋白质,但是线粒体和叶绿体的功能所需要的大多数蛋白质还是由细胞核表达形成的。
另外,线粒体和叶绿体不能在分裂的细胞中凭空产生,而是依靠线粒体和叶绿体的分裂增殖,然后再分裂的两个细胞中分配,这一点是它们被称为半自主遗传细胞器的主要原因。
细胞核中核仁的作用是什么?
核仁
核仁是真核细胞间期核中最明显的结构。
在光镜下的染过色的细胞内,或者相差显微镜下的活细胞中,或者分离细胞的细胞核内,都容易看到核仁,它通常是单一的或者多个匀质的球形小体。
核仁的大小、形状和数目随生物的种类、细胞类型和细胞代谢状态而变化。
蛋白质合成旺盛、活跃生长的细胞,如分泌细胞、卵母细胞的核仁大,可占总核体积的25%不具蛋白质合成能力的细胞,如肌肉细胞、休眠的植物细胞,其核仁很小。
在细胞周期过程中,核仁是一个高度动态的结构,在有丝分裂期间表现出周期性的消失与重建。
真核细胞的核仁具有重要功能,它是rRNA合成、加工和核糖体亚单位的装配场所。
因此对核仁结构、动态和功能的研究,不仅为早期细胞学家所密切注意,而且在20世纪60年代发现核仁的重要功能以后,也一直受到各相关领域研究者的高度重视。
例析线粒体和叶绿体的内共生起源学说封开县江口中学陶勇由于线粒体和叶绿体具有独特的半自主性并与细胞核建立了复杂而协调的互作关系,它们的起源一直以来多被认为有别于其他细胞器。
在人们为这两种细胞器设计的起源假说中,内共生起源学说很好地贴合了线粒体和叶绿体的半自主性和核质关系特征,因而得到了广泛的认可和支持。
内共生起源学说认为,线粒体和叶绿体分别起源于原始真核细胞内共生的行有氧呼吸的细菌和行光能自养的蓝细菌。
该假说的提出远早于mtDNA和cpDNA的发现。
随着人们对真核细胞超微结构、线粒体和叶绿体DNA及其编码机制的认识,内共生起源学说的内涵得到了进一步充实。
1970年,Margulis在已有的资料基础上提出了一种更为细致的设想。
假设认为,真核细胞的祖先是一种体积较大、不需氧具有吞噬能力的细胞,通过糖酵解获取能量。
而线粒体的祖先则是一种革兰氏阴性菌,具备三羧酸循环所需的酶和电子传递链系统,可利用氧气把糖酵解的产物丙酮酸进一步分解,获得比糖酵解更多的能量。
当这种细菌被原始真核细胞吞噬后,即与宿主细胞间形成互利的共生关系:原始真核细胞利用这种细菌获得更充分的能量;而这种细菌则从宿主细胞获得更适宜的生存环境。
与此类似,叶绿体的祖先可能是原核生物的蓝细菌〔cyanbacteria)。
当这种蓝细菌被原始真核细胞摄人后,为宿主细胞进行光合作用;而宿主细胞则为其提供其他的生存条件。
线粒体和叶绿体的内共生学说先后得到了大量的生物学研究证据的支持。
特别是近期的分子生物学和生物信息学的研究发现真核细胞的细胞核中存在大量原本可能属于呼吸细菌或蓝细菌的遗传信息,说明最初的呼吸细菌和蓝细菌的大部分基因组在漫长的共进化过程中发生了向细胞核的转移。
这种转移极大的削弱了线粒体和叶绿体的自主性,建立起稳定、协调的核质互作关系。
一、线粒体和叶绿体内共生起源学说的主要论据线粒体和叶绿体具有细菌基因组的典型特征。
它们均为单条环状双链DNA分子,不含5-甲基胞嘧啶,无组蛋白结合并能进行独立的复制和转录。
叶绿体与线粒体的半自主性及其起源引言叶绿体和线粒体是细胞内的两种重要的细胞器,它们具有一定的半自主性。
这种半自主性是指叶绿体和线粒体都有自己的DNA和独立的基因组成,并且可以通过自身复制和修复来保持其稳定性和功能。
本文将探讨叶绿体和线粒体的半自主性及其起源。
叶绿体的半自主性叶绿体是植物细胞和某些原生生物细胞中的一种细胞器。
它在光合作用中扮演着重要角色。
除了光合作用,叶绿体还具有一些自主功能,比如自我复制和自我修复。
叶绿体复制是一个复杂的过程,涉及到DNA复制、蛋白质合成和分裂等方面。
研究表明,叶绿体复制主要由细胞核和叶绿体两个部分合作完成。
细胞核主要负责合成叶绿体的DNA和RNA,而叶绿体本身则负责合成叶绿体自身所需要的蛋白质。
叶绿体的自我修复也是一个非常重要的功能。
由于环境压力和各种因素的影响,叶绿体很容易发生损伤和退化。
为了保持其光合作用的正常进行,叶绿体需要进行修复。
研究表明,叶绿体自我修复主要通过巨噬细胞内吞叶绿体碎片来完成。
线粒体的半自主性线粒体是细胞内的一个细胞器,主要负责细胞内的能量生产。
线粒体的半自主性与叶绿体类似,它也有自己的DNA和独立的基因组成,并且可以通过自我复制和修复来保持其稳定性和功能。
线粒体复制是一个复杂的过程,涉及到DNA复制、蛋白质合成和分裂等方面。
线粒体复制主要由细胞核和线粒体两个部分合作完成。
细胞核主要负责合成线粒体的DNA和RNA,而线粒体本身则负责合成线粒体自身所需要的蛋白质。
线粒体的自我修复也是一个非常重要的功能。
线粒体的自我修复主要涉及到细胞核和线粒体之间的相互交流和协作。
当线粒体发生损伤和退化时,细胞核将会向它发送一个信号来进行修复,并且调节其自身的代谢状态以适应新的环境。
叶绿体和线粒体半自主性的起源叶绿体和线粒体的半自主性起源于它们的祖先。
据推测,叶绿体和线粒体的祖先曾经是一种自主生命体,类似于现在的细胞寄生体。
这种自主生命体在进化过程中逐渐演化成了现在的叶绿体和线粒体。