细胞生物学之笔记--第4章细胞膜与物质的穿膜运输.
- 格式:doc
- 大小:330.03 KB
- 文档页数:10
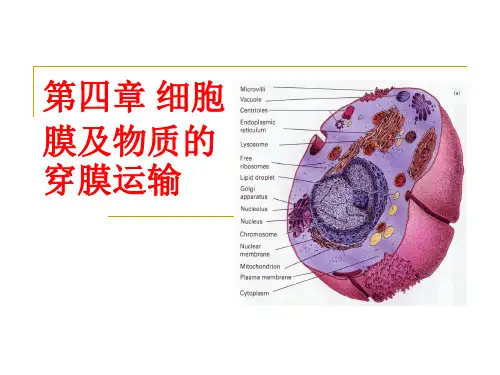


第四章细胞膜与物质的穿膜运输细胞膜:是包围在细胞质表面的一层薄膜,又称质膜。
内膜系统:除质膜外,细胞内还有丰富的膜结构,它们形成了细胞内各种膜性细胞器,如内质网、高尔基复合体、溶酶体、各种膜泡等,称为细胞的内膜系统。
生物膜:质膜和细胞内膜系统的总称。
单位膜:生物膜因在电子显微镜下呈“两暗夹一明”的形态结构,又称为生物膜。
脂质体:脂质分子在水环境中排列呈双层,两层分子的疏水尾部被亲水头部夹在中间,为了避免双分子层两端疏水尾部与水接触,其游离端往往能自动闭合形成充满液体的球状小泡。
孔蛋白:有些穿膜蛋白以β-折叠片层构象穿膜,在脂双层中围成筒状结构,称β筒,有些β筒在质膜上起运输蛋白的作用,称为孔蛋白,主要存在于线粒体、叶绿体和一些细菌的外膜。
膜内在蛋白(穿膜蛋白、整合蛋白):占膜蛋白总量70-80%,两亲性分子,分为单次穿膜、多次穿膜和多亚基穿膜蛋白三种类型。
膜外在蛋白(周边蛋白):占膜蛋白总量20-30%,是一类与细胞膜结合比较松散的不插入脂双层的蛋白质,分布在质膜的胞质侧或胞外侧。
如红细胞的血影蛋白和锚蛋白。
脂锚定蛋白(脂连接蛋白):可位于膜两侧,以共价键与脂双层内的脂分子结合。
糖基磷脂酰肌醇锚定蛋白(GPI):位于质膜外的表面的一些蛋白质,通过与脂双层外层中磷脂酰肌醇分子相连的寡糖链共价键结合而锚定到质膜上,这些蛋白称为GPI细胞外被(糖萼):大多数真核细胞表面富含糖类的周缘区,现一般用来指与质膜相连接的糖类物质,即质膜中糖蛋白和糖脂向外表面延伸出的寡糖链部分,所以细胞外被实质上是质膜结构的一部分,基本功能是保护细胞抵御各种物理、化学性损伤。
(不与质膜相连的细胞外覆盖物称为细胞外物质或胞外结构)膜的不对称性:细胞膜中各种成分如膜脂,膜蛋白,膜糖,分布是不均匀的,包括种类和数量上都有很大差异。
(如红细胞外层鞘磷脂SM最多,内层磷脂酰乙醇胺PE即脑磷脂最多)脂双层的液晶态:脂双层作为生物膜的主体,它的组分既有固体分子排列的有序性,又有液体的流动性,这一两种特性兼有的居于晶态和液态之间的状态即液晶态。




第四章细胞膜与物质的穿膜运输第一节细胞膜的化学组成与生物特性一、细胞膜的化学组成(一)膜脂构成细胞膜的结构骨架1.磷脂是膜脂的主要成分2.胆固醇能够稳定细胞膜和调节膜的流动性3.糖脂主要位于质膜的非胞质面(二)膜蛋白以多种方式与脂双分子层结合1.膜内在蛋白2.膜外在蛋白3.脂锚定蛋白(三)膜糖类覆盖细胞膜表面二、细胞膜的生物学特性(一)膜的不对称性决定膜功能的方向性1.膜脂的不对称性2.膜蛋白的不对称性3.膜糖的不对称性(二)膜的流动性是膜功能活动的保证1.脂双层为液晶态二维流体2.膜脂分子的运动方式①侧向扩散lateral diffusion②翻转运动flip-flop③旋转运动rotation④弯曲运动flexion3.影响膜脂流动性的因素①脂肪酸链的饱和程度②脂肪酸链的长短③胆固醇的双重调节作用④卵磷脂与鞘磷脂的比值⑤膜蛋白的影响4.膜蛋白的运动性①侧向扩散②旋转运动三、细胞膜的分子结构模型(一)片层结构模型具有三层夹板式结构特点(二)单位膜模型体现膜形态结构的共同特点(三)流动镶嵌模型是被普遍接受的模型(四)脂筏模型深化了对膜结构和功能的认识第二节小分子物质和离子的穿膜运输一、膜的选择性通透和简单扩散二、膜运输蛋白介导的穿膜运输(一)易化扩散是载体蛋白介导的被动运输(二)主动运输时载体蛋白逆浓度梯度的耗能运输1.ATP驱动泵2.协同运输:共运输+对向运输(三)离子通道高效转运各种离子1.离子通道的特点2.离子通道的类型①配体门控通道②电压门控通道③应力门控通道(四)水通道介导水的快速转运1.水通道的分类2.水通道蛋白的结构3.水通道对水分子的筛选机制第三节大分子和颗粒物质的穿膜运输一、胞吞作用(一)吞噬作用是吞噬细胞摄入颗粒物质的过程(二)胞饮作用是细胞吞入液体和可溶性物质的过程(三)受体介导的胞吞提高摄入特定物质的效率1.有被小窝和有被小泡的形成2.无被小泡形成并与内体融合3.受体介导的LDL胞吞作用二、胞吐作用(一)连续性分泌是不受调节持续不断的细胞分泌(二)受调分泌是细胞外信号调控的选择性分泌第四节细胞膜异常与疾病一、载体蛋白异常与疾病1.胱氨酸尿症是载体蛋白异常性疾病2.肾性糖尿是葡萄糖载体蛋白异常性遗传病二、离子通道蛋白异常与疾病1.囊性纤维病(cystic fibrosis,CF)细胞膜上一个受cAMP调节的氯离子通道异常。


高一生物必修一第四章物质跨膜运输的方式知识点物质跨膜运输的方式是指一些脂溶性的物质由膜的高浓度一侧向低浓度一侧移动的过程。
以下是第四章物质跨膜运输的方式知识点,希望对大家有帮助。
被动运输自由扩散(简单扩散)定义:物质通过简单扩散作用(simple transport)进出细胞,叫做自由扩散.其特点是:①沿浓度梯度(或电化学梯度)扩散;②不需要提供能量;③没有膜蛋白的协助。
某种物质对膜的通透性(P)可以根据它在油和水中的分配系数(K)及其扩散系数(D)来计算:P=KD/t,t 为膜的厚度。
脂溶性越高通透性越大,水溶性越高通透性越小;非极性分子比极性容易透过,小分子比大分子容易透过。
具有极性的水分子容易透过是因水分子小,可通过由膜脂运动而产生的间隙。
非极性的小分子如O2、CO2、N2 可以很快透过脂双层,不带电荷的极性小分子,如水、尿素、甘油等也可以透过人工脂双层,尽管速度较慢,分子量略大一点的葡萄糖、蔗糖则很难透过,而膜对带电荷的物质如:H+、Na+、K+、Cl—、HCO3—是高度不通透的事实上细胞的物质转运过程中,透过脂双层的简单扩散现象很少,绝大多数情况下,物质是通过载体或者通道来转运的。
离子、葡萄糖、核苷酸等物质有的是通过质膜上的运输蛋白的协助,按浓度梯度扩散进入质膜的,有的则是通过主动运输的方式进行转运。
举例:氧气,二氧化碳,水,甘油,乙醇,苯,脂肪酸,尿素,胆固醇,脂溶性维生素,气体小分子等协助扩散也称促进扩散、易化扩散(faciliatied diffusion),其运输特点是:①比自由扩散转运速率高; ②存在最大转运速率; 在一定限度内运输速率同物质浓度成正比。
如超过一定限度,浓度不再增加,运输也不再增加。
因膜上载体蛋白的结合位点已达饱和; ③有特异性,即与特定溶质结合。
这类特殊的载体蛋白主要有离子载体和通道蛋白两种类型。
④不需要提供能量。
举例:红细胞摄取葡萄糖载体蛋白:能够与溶剂结合,通过对自身构象的改变而介导该溶质跨膜运输。

第四章细胞膜与物质的穿膜运输第一节细胞膜的化学组成与生物特性一、细胞膜的化学组成细胞膜上的脂类=膜脂〔membrane lipid〕,约占膜成分的50%,主要有磷脂〔phospholipid〕、胆固醇〔cholesterol〕、和糖脂〔glycolipid〕(一)膜脂构成细胞膜的结构骨架1.磷脂是膜脂的主要成分➢甘油磷酸的共同特征:以甘油为骨架,甘油分子的1、2位羟基分别于脂肪酸形成酯键,3位羟基与磷酸基团形成酯键.磷酸基团结合胆碱/乙醇胺/丝氨酸/肌醇.脂肪酸链长短不一,通常14~24个碳原子,一条脂肪酸链不含双键,另一条含有一个或几个双键,形成30°弯曲.➢鞘磷脂以鞘氨醇代替甘油,鞘氨醇的氨基结合长链的不饱和脂肪酸,分子末端的一个羟基与胆碱磷酸结合,另一个游离羟基可与相邻分子的极性头部、水分子或膜蛋白形成氢键.鞘磷脂与其代谢产物神经酰胺、鞘氨醇、1-磷酸鞘氨醇参与各种细胞活动.神经酰胺是第二信使;1-磷酸鞘氨醇在细胞外通过G蛋白偶联受体起作用,在细胞内与靶蛋白作用2.胆固醇能够稳定细胞膜和调节膜的流动性✧胆固醇为两性极性分子.✧极性头部为连接于固醇环〔甾环〕上的羟基,靠近相邻的磷脂分子.✧固醇环疏水,富有刚性,固定在磷脂分子临近头部的烃链上,对林芝的脂肪酸尾部的运动具有干扰作用.✧尾部为疏水性烃链.埋在磷脂的疏水尾部中.✧胆固醇分子调节膜的流动性和加强膜的稳定性.没有胆固醇,细胞膜会解体.PS.不同生物膜有各自特殊的脂类组成.哺乳动物细胞膜上富含胆固醇和糖脂,线粒体膜内富含心磷脂;大肠杆菌质膜则不含胆固醇.3.糖脂主要位于质膜的非胞质面糖脂含量占膜脂总量5%以下,遍布原核、真核细胞表面细菌和植物的糖脂均是甘油磷脂衍生物,一般是磷脂酰胆碱PC 衍生来动物糖脂都是鞘氨醇衍生物,称为鞘糖脂,糖基取代磷脂酰胆碱,成为极性头部已发现40多种糖脂,区别在于极性头部不同,由1至几个糖残基构成✧最简单的糖脂是脑苷脂,极性头部只是一个半乳糖/葡萄糖残基✧最复杂的糖脂是神经节苷脂,极性头部有七个糖残基;在神经细胞膜中最丰富,占总膜脂5%~10%✧脂质体〔lipidsome〕可以作运载体(二)膜蛋白以多种方式与脂双分子层结合又称含量作用力特点膜内在蛋白穿膜蛋白70%~80% X德华力α-螺旋构象/β-筒孔蛋白1.内在膜蛋白✧又称跨膜蛋白,占膜蛋白总量70%~80%;分单次跨膜、多次跨膜、多亚基跨膜三种类型✧跨膜区域20~30个疏水氨基酸残基,通常N端在细胞外侧✧内在膜蛋白跨膜结构域与膜脂结合区域,作用方式:①疏水氨基酸形成α-螺旋,跨膜并与脂双层脂肪酸链通过X德华力相互作用②某些α-螺旋外侧非极性,内侧是极性链,形成特异性畸形分子的跨膜通道✧多数跨膜区域是α-螺旋,也有以β-折叠片多次穿膜形成筒状结构,称β-筒,如孔蛋白<porin>2.外在膜蛋白➢又称外周蛋白,占膜蛋白总量20%~30%;完全在脂双层之外,胞质侧或胞外侧,通过非共价键附着膜脂或膜蛋白➢胞质侧的外周蛋白形成纤维网络,为膜提供机械支持,也连接整合蛋白,如红细胞的血影蛋白和锚蛋白➢外周蛋白为水溶性蛋白,与膜结合较弱,改变溶液离子浓度或pH,可分离它们而不破坏膜结构3.脂锚定蛋白①一种位于膜的两侧,蛋白质直接以共价键结合于脂类分子;此种锚定方式与细胞恶变有关②还有糖基磷脂酰肌醇锚定蛋白<GPI>,通过蛋白质C端与磷脂酰肌醇连接的糖链共价结合脂锚定蛋白在膜上运动性增大〔侧向运动〕,有利于结合更多蛋白,有利于更快地与胞外蛋白结合、反应GPI-锚定蛋白分布极广,100种以上,如多种水解酶、免疫球蛋白、细胞黏附分子、膜受体等4.去垢剂〔detergent〕离子型去垢剂:SDS十二烷基磺酸钠引起蛋白质变性非离子型去垢剂:Triton X-100 对蛋白质比较温和(三)膜糖类覆盖细胞膜表面细胞膜的糖类,占质膜重量2%~10%;①大多以低聚糖或多聚糖共价结合膜蛋白,形成糖蛋白〔糖蛋白中的糖基化主要发生在天冬酰胺〔N-连接〕,其次是丝氨酸和苏氨酸〔O-连接〕残基上〕;②或以低聚糖共价结合膜脂,形成糖脂,所有糖链朝向细胞外表面形成低聚糖的单糖类型:甘露糖、岩藻糖、半乳糖、半乳糖胺、葡萄糖、葡萄糖胺、唾液酸等A.唾液酸残基在糖链末端,形成细胞外表面净负电荷B.寡糖链中的单糖的数量、种类、排列顺序、有无支链等不同,可以出现千变万化的组合形式.Eg.人类ABO血腥抗原的差别就是血型糖蛋白在红细胞质膜外表面寡糖链的组成结构决定.△细胞外被cell coat=糖萼glycocalyx=与质膜相连的糖类物质功能:①保护细胞抵御各种物理、化学性损伤②建立起水盐平衡③帮助蛋白质膜上定位、固定,防止翻转④参与细胞与外环境的作用,eg识别、粘附、迁移二、细胞膜的生物学特性(一)膜的不对称性决定膜功能的方向性膜结构上的不对称性保证了膜功能的方向性和生命活动的高度有序性1.膜脂的不对称性SM、PC在细胞外侧叫多,PE在细胞内侧较多.2.膜蛋白的不对称性➢各种膜蛋白在质膜中有特定位置,分布绝对不对称:酶和受体多分布于质膜的外侧面,而腺苷酸环化酶定位内侧面➢跨膜蛋白有一定方向性:多数N外C内,两端肽链长度、氨基酸种类、活性位点不同3.膜糖的不对称性都向着非胞质面(二)膜的流动性是膜功能活动的保证流动性fluidity主要是指膜脂的流动性+膜蛋白的运动性1.脂双层为液晶态二维流体✓液晶态〔lipid-crystal state〕脂双分子层已有固体分子排列的有序性,又有液体的流动性.细胞内外的水环境,使膜脂分子不能从脂双层逸出,只能在二维平面交互位置.✓相变〔phase transition〕正常体温下,膜呈液晶态;当温度下降到临界温度<膜的相变温度>,膜脂转为晶态✓膜的流动性是膜功能活动的保证.2.膜脂分子的运动方式①侧向扩散lateral diffusion =脂双层的单分子层内,脂分子沿膜平面侧向与相邻分子快速交换位置,每秒约107次.侧向扩散运动时膜脂分子主要的运动方式..②翻转运动flip-flop 从脂双层一层翻转到另一层,需要翻转酶,在内质网发生③旋转运动rotation 膜脂分子围绕与膜平面向垂直的轴的自旋运动④弯曲运动flexion 膜脂分子的烃链是有韧性、可弯曲的,分子尾部端弯曲、摆动幅度大,而靠近头部弯曲摆动幅度小.⑤此外,还有伸缩、震荡3.影响膜脂流动性的因素①脂肪酸链的饱和程度磷脂分子长的饱和脂肪酸链呈直线型,具有最大的聚集倾向而排列紧密成凝胶状态;不饱和脂肪酸链在双键出形成折曲而呈弯曲状,感染了脂分子间X德华力的相互作用,故排列疏松,从而增加了膜的流动性.∴脂双分子层中含有的不饱和脂肪酸越多,膜的相变温度越低,流动性越大.环境温度降低时,A.细胞通过去饱和酶〔desaturases〕催化将胆碱去饱和形成双键.B.通过磷脂酶&脂酰转移酶在不同的磷脂分子之间重组脂肪酸链以产生含两个不饱和脂肪酸链的磷脂分子.②脂肪酸链的长短脂肪酸链短的相变温度低,流动性大.短→尾端不易发生相互作用;长→不仅可以在同一分子称内相互作用,而且可以与另一分子层中的长链尾端相互作用③胆固醇的双重调节作用A.当温度在相变温度以上时,由于胆固醇分子的固醇环与磷脂分子靠近极性头部的烃链部分结合,限制了这几个CH2的运动,起到稳定质膜的作用.B.当温度在相变温度以下时,由于胆固醇位于磷脂分子之间隔开磷脂分子,可有效地防止脂肪酸链相互凝聚,干扰晶态的形成.④卵磷脂与鞘磷脂的比值哺乳动物细胞中,卵磷脂和鞘磷脂的含量约占膜脂的50%,卵磷脂的脂肪酸链不饱和程度高,相变温度较低;鞘磷脂则相反.在细胞衰老过程中,卵磷脂和鞘磷脂的比值下降,流动性也下降.⑤膜蛋白的影响膜蛋白嵌入膜脂疏水区后,是周围的脂类分子不能单独活动而形成界面脂;在含较多内在蛋白的膜中,存在有内在蛋白分割包围的富脂区〔lipid-rich region〕磷脂分子智能在一个富脂区内自有扩散,而不能扩散到邻近的富脂区此外,膜脂的极性基团、环境温度、pH值、离子强度等都对膜脂流动性产生一定影响.环境温度高,膜脂流动性大;相变温度内,每下降10℃,膜的粘性增加3倍,膜流动性降低4.膜蛋白的运动性①侧向扩散膜蛋白在膜脂中可以自有漂浮&在膜表面扩散.人鼠杂交细胞表面抗原分布变化可证明.目前测定膜蛋白的侧向扩散常采用光致漂白荧光恢复法〔fluorescence recovery after photobleaching,FRAP〕②旋转运动膜蛋白能围绕与膜平面相垂直的轴进行旋转运动.速度比侧向扩散慢;不同膜蛋白速度不同,有些膜蛋白无法运动;膜蛋白周围脂质的流动性影响膜蛋白的流动性膜蛋白的运动不需要消耗能量膜的流动性意义重大:物质运输、细胞识别、信息传导等;生物膜的各种功能都是在膜的流动状态下进行的,膜的流动过低,代谢终止三、细胞膜的分子结构模型(一)片层结构模型具有三层夹板式结构特点1935年,James Danielli 和Hugh Davson发现细胞膜的表面X力显著低于油-水界面表面X力,推测质膜中有蛋白质;提出"片层结构模型〞<蛋白-磷脂-蛋白三层夹板式结构> (二)单位膜模型体现膜形态结构的共同特点1959年,J.D.Robertson 电镜观察细胞膜"两暗夹一明〞——单位膜单位膜模型:膜蛋白是单层肽链以β折叠通过静电作用与磷脂极性端结合;能对膜的某些属性进行解释,被普遍采用,但是把膜作为静止的单一结构(三)流动镶嵌模型是被普遍接受的模型1972年,"流动镶嵌模型〞Fluid mosaic model:磷脂双层构成膜的连续主体,具有晶体的有序性和液体的流动性;球形蛋白质分子以不同形式结合脂双层分子;膜是一种动态的、不对称的具有流动性结构1975年,"晶格镶嵌模型〞:膜脂可逆地进行"有序<液态>〞和"无序<晶态>〞相变,膜蛋白对膜脂的运动具有限制作用,流动性是局部的1977年,"板块镶嵌模型〞:流动的脂双层中存在能独立移动脂类板块(四)脂筏模型深化了对膜结构和功能的认识✓脂双层中由特殊脂质和蛋白质组成的微区,富含胆固醇和鞘脂类,聚集特定种类膜蛋白;此膜区较厚〔鞘脂类脂肪酸链较长〕,称"脂筏〞Lipid rafts,其周围富含不饱和磷脂,流动性较高✓脂筏的两个特点:许多蛋白聚集在脂筏内,便于相互作用;脂筏提供有利于蛋白质变构的环境,形成有效构象✓脂筏功能:参与信号转导、受体介导内吞作用、胆固醇代谢运输等第二节小分子物质和离子的穿膜运输一、膜的选择性通透和简单扩散简单扩散<simple diffusion>: 小分子的热运动使分子以自由扩散的方式由膜一侧扩散到另一侧,条件:溶质在膜两侧有一定浓度差,溶质必须能透过膜脂溶性物质如醇、苯、甾类激素、O2、CO2、NO、H2O 通过简单扩散跨膜简单扩散不需要运输蛋白协助,顺浓度梯度由高浓度向低浓度方向扩散,不消耗能量;也称"被动扩散〞passive diffusion二、膜运输蛋白介导的穿膜运输除了水和非极性小分子,绝大多数溶质如各种离子、葡萄糖、氨基酸、核苷酸等都不能简单扩散穿膜转运特定膜蛋白——膜运输蛋白<跨膜蛋白,每种只转运一种特定类型溶质>膜运输蛋白分两类:①载体蛋白carrier protein:与特定溶质结合,改变构象使溶质穿越细胞膜②通道蛋白channel protein:形成水溶性通道,贯穿脂双层,通道开放时,特定溶质<无机离子>可穿越脂双层➢"被动运输" passive transport所有通道蛋白和许多载体蛋白,转运溶质分子不消耗能量,消耗顺电化学浓度梯度的势能➢"主动运输〞active transport逆电化学浓度梯度转运溶质,需要载体蛋白参与,还需要消耗能量ATP;这种利用代谢产生能量的进行逆浓度梯度的转运称为主动运输.能量来源:ATP水解、光吸收、电子传递、顺浓度梯度的离子运动etc(一)易化扩散是载体蛋白介导的被动运输∆"易化扩散" facilitated diffusion=帮助扩散=非脂溶性或亲水性小分子,不能简单扩散通过细胞膜,需载体蛋白介导不消耗代谢能量,顺物质浓度梯度或电化学梯度进行转运∆特点:特定易化转运蛋白介导特定物质在两个方向的穿膜运输,取决于该物质在膜两侧的相对浓度——转运特异性强,速率快∆作用机制:载体蛋白对所转溶质具有高度专一性,其分子上的结合位点与某一溶质进行短暂的可逆的结合,引起载体蛋白构象变化,转运溶质分子从膜一侧到另一侧;载体与溶质亲和力下降,释放溶质,构象恢复∆例子:多数细胞<低浓度葡萄糖>从血流和组织液中<高浓度葡萄糖>,通过易化扩散获取葡萄糖.人类基因组编码14种葡萄糖转运载体蛋白glucose transporter, GLUT,构成GLUT家族.它们具有高度同源氨基酸序列,均含有12次跨膜的α-螺旋,α-螺旋含有丝氨酸Ser、苏氨酸Thr、天冬氨酸Asp和谷氨酸残基Glu,其侧链与葡萄糖羟基形成氢键,是葡萄糖结合位点.GLUT的异常或缺陷是2型糖尿病的病因之一.红细胞膜上存在5万个葡萄糖载体蛋白,占膜总蛋白5% 最大转运速率每秒180个葡萄糖分子.(二)主动运输时载体蛋白逆浓度梯度的耗能运输1.ATP驱动泵在胞质侧有一个或多个ATP结合位点,水解A TP从低浓度向高浓度转运协同运输〔co-transport〕是一类由Na+-K+泵〔或H+泵〕与载体蛋白协同作用,间接消耗ATP 所完成的主动运输方式.物质穿膜运动的直接动力来自膜两侧离子的电化学梯度中的能量,而维持这种离子电化学梯度是通过Na+-K+泵〔或H+泵〕消耗ATP来实现的.动物利用Na+-K+泵,植物利用H+泵①共运输:两种溶质分子同一方向穿膜运输.Eg肠腔细胞膜的"Na+/葡萄糖协同运输蛋白〞Na+/glucose cotransporter在质膜外表面结合2个Na+和一个葡萄糖.进入细胞的Na+之后再被Na+-K+泵排出,维持Na+膜内外浓度差.葡萄糖一旦进入小肠细胞,再以易化扩散方式进入血流.主动运输特点:①逆浓度或电化学梯度跨膜转运②消耗能量,直接水解A TP或离子电化学梯度提供能量③膜上特异性载体蛋白介导,载体特异结合转运溶质,载体构象可变(三)离子通道高效转运各种离子1.离子通道的特点〔顺梯度,高选择,高效率,受调控.〕①只介导被动运输,溶质从膜的高浓度一侧自由扩散到低浓度一侧②离子通道对被转运离子的大小所带电荷有高度选择性③转运效率高,通道允许106~108个特定离子/秒通过,比最快效率的载体蛋白高1000倍④离子通道不是持续开放,有开和关两种构象,受信号调控2.离子通道的类型①配体门控通道ligand-gated channel.实际为离子通道型受体,它们与细胞外的特定配体ligand结合后,发生构象改变,结果吧"门〞打开,允许某种离子快速穿膜扩散.Eg.烟碱型乙酰胆碱受体nAChR是典型的配体门控阳离子通道,大量存在与骨骼肌神经接头处.4种不同亚基单组成的五聚体穿膜蛋白〔α2βγδ〕→梅花状通道与结构.①神经冲动→神经末梢→细胞去极化→电压门控Ca2+通道开放→细胞外Ca2+涌入细胞→胞内突触小泡释放乙酰胆碱至突触间隙②释放的乙酰胆碱→结合突触后膜的乙酰胆碱受体→通道开放,Na+流入肌细胞→肌细胞膜局部去极化③肌细胞去极化→诱发膜上Na+通道开放→大量Na+涌入肌细胞,使整个肌细胞膜进一步去极化④肌细胞膜的去极化→使肌浆网上Ca2+通道开放→Ca2+大量释放如胞质→肌原纤维收缩②电压门控通道voltage-gated channel膜电位的改变是控制电压门通道开放与关闭的直接因素.反应快.主要存在与神经元、肌细胞与腺上皮细胞等兴奋细胞,包括钾通道、钙通道、氯通道③应力门控通道stress-activated channel应力激活通道是通道蛋白感受应力而改变构象,通道开放,离子跨膜,膜电位变化Eg.A.内耳听觉毛细胞顶部的听毛具有应力激活通道,受到声波振动而弯曲,应力门控通道开放,离子跨膜进入毛细胞改变膜电位,将声波信号传递给听觉神经元B.细菌与古细菌的应力激活通道均为跨膜蛋白五聚体,通透阳离子(四)水通道介导水的快速转运1.水通道的分类哺乳类水通道蛋白家族已有11个,根据功能特性的差异,分为两个家族:AQP1、2、4、5、6和AQP0 基因结构类似,氨基酸序列同源30%~50%,只能通透水,经典的选择性水通道;AQP3、7、9、10 除通透水,对甘油、尿素等中性小分子也具有通透性,第二家族——水-甘油通道;AQP8位于水选择型与甘油渗透型之间2.水通道蛋白的结构AQP1由4个对称排列的圆筒状亚基围成的四聚体,每个亚基中心的中央孔直径0.28nm,只允许水分子通过.每个AQP1亚基有6个长α螺旋构成基本骨架,两个短嵌入式α螺旋顶对顶排列,顶端均有保守的Asn-Pro-Ala基序,使得顶对顶稳定每个亚基的α螺旋朝向脂双层的一面是非极性氨基酸残基,朝向中央孔的一面是极性氨基酸残基3.水通道对水分子的筛选机制水孔蛋白对水分子高度特异性选择,因为:每个亚基中央孔的直径0.28nm,只比水分子大一点点,限制其它分子通过;每个亚基中央孔道内有特异溶质结合位点,每个水分子通过时,孔道内的3极性氨基酸残基的羰基氧与水分子形成氢键;离子与水分子的复合物比孔道大得多,不能通过水通道持续开放,每秒通过3×109个水分子,不耗能,水分子移动方向由膜两侧渗透压决定,低→高第三节大分子和颗粒物质的穿膜运输大分子物质不能通过膜转运蛋白进入细胞,由膜包围形成膜泡,然后通过膜泡形成和融合来完成转运——小泡运输细胞摄入大分子或颗粒物质的过程,称胞吞作用<endocytosis>细胞排出大分子或颗粒物质的过程,称胞吐作用<exocytosis>胞吞胞吐涉与膜泡的融合与断裂,需要消耗能量,属于主动运输以上膜泡运输转运量较大,也称批量运输;膜泡运输也发生于胞内各种膜性细胞器一、胞吞作用(一)吞噬作用是吞噬细胞摄入颗粒物质的过程免疫系统具有吞噬功能的中性粒细胞、单核细胞、巨噬细胞在摄取大固体颗粒或分子复合物<直径>250nm>时进行细胞膜凹陷或形成伪足,将大颗粒包裹摄入细胞,形成膜泡"吞噬体〞这些免疫细胞通过此方式吞噬入侵微生物、清除损伤和死亡细胞(二)胞饮作用是细胞吞入液体和可溶性物质的过程细胞非特异摄取细胞外液的过程;胞饮发生在质膜的特殊区域,质膜内陷形成小窝,包围液体物质,形成"胞饮体〞,直径小于150nm胞饮作用分为两种类型:①液相内吞:非特异固有内吞作用,摄入细胞外液与可溶性物质;②吸附内吞:细胞外大分子/小颗粒物质以某种方式吸附在细胞表面,具有一定特异性在能形成伪足和转运功能活跃的细胞中多见,如巨噬细胞、白细胞、毛细血管细胞、肾小管上皮细胞、小肠上皮细胞等(三)受体介导的胞吞提高摄入特定物质的效率受体介导的内吞作用receptor mediated endocytosis 是细胞通过受体的介导选择性高效摄取细胞外特定大分子物质的过程可特异性摄入胞外含量很低的成分,比胞饮作用内化效率高1000多倍1.有被小窝和有被小泡的形成◆细胞膜上有多种受体蛋白,往往同类受体蛋白集中在膜特定区域,称"有被小窝〞coatedpit;小窝内受体浓度是质膜其它处的10~20倍◆各种有被小窝约占质膜表面积2%,此处质膜向内凹陷,直径50~100 nm,此处质膜内表面覆盖网格蛋白和衔接蛋白◆网格蛋白,又称"笼蛋白〞,由3条重链和3条轻链组成;3个重链轻链的二聚体,形成三腿蛋白复合物→自我装配,自动形成篮网状结构网格蛋白作用:牵拉质膜向内凹陷,参与捕获特定膜受体使其汇聚有被小窝◆衔接蛋白参与有被小泡组成,处于网格蛋白与配体-受体复合物间◆不同类型的衔接蛋白结合不同类型膜受体,使细胞捕获不同配体网格蛋白没有特异性2.无被小泡形成并与内体融合✧配体结合膜上受体,通过衔接蛋白,网格蛋白聚集在膜的胞质侧,网格由6边形转变成5边形,促进网格蛋白外被弯曲变成笼形,牵动质膜凹陷✧发动蛋白<dynamin> ——GTP结合蛋白,自动组装成一个螺旋状领圈结构,水解GTP,构象改变,将有被小泡从质膜上切离下来,形成网格蛋白有被小泡✧有被小泡很快脱去包被<笼蛋白重新利用> →无被小泡→与早期内体融合✧内体:动物细胞中经胞吞作用形成的膜包围的细胞器,作用是运输由胞吞作用新摄入的物质到溶酶体被降解.内体膜上有A TP驱动的质子泵,将H+泵入内体腔,降低腔内pH 〔pH5~6〕✧低pH使受体与配体分离,内体出芽形成运载受体的小囊泡,返回质膜;受体重新利用,含配体的内体与溶酶体融合3.受体介导的LDL胞吞作用❖胆固醇是构成膜的成分,也是类固醇激素的前体;动物细胞通过受体介导的胞吞作用摄入所需大部分胆固醇.❖胆固醇在肝脏合成并包装成低密度脂蛋白<low density lipoprotein, LDL>,在血液中运输❖LDL为球状颗粒,分子量3106,直径22nm;中心是1500个酯化的胆固醇分子,外面包围800个磷脂分子和500个游离胆固醇分子❖载脂蛋白ApoB100是细胞膜上LDL受体的配体, 组装LDL成颗粒二、胞吐作用(一)连续性分泌是不受调节持续不断的细胞分泌分泌蛋白在粗面内质网合成后,转运到高尔基体进行修饰、浓缩、分选,形成分泌泡,被转运到细胞膜,与膜融合,外排蛋白的过程分泌蛋白:驻留蛋白、膜蛋白、细胞外基质组分等(二)受调分泌是细胞外信号调控的选择性分泌分泌蛋白合成后,包裹于分泌囊泡,储存于胞质中,受到细胞外信号刺激,引起细胞内Ca2+浓度瞬时升高,才启动胞吐作用此种分泌途径只存在于特化细胞,如分泌激素、酶、神经递质的细胞第四节细胞膜异常与疾病一、载体蛋白异常与疾病1.胱氨酸尿症是载体蛋白异常性疾病2.肾性糖尿是葡萄糖载体蛋白异常性遗传病二、离子通道蛋白异常与疾病1.囊性纤维病〔cystic fibrosis,CF〕细胞膜上一个受cAMP调节的氯离子通道异常.三、膜受体异常与疾病1.家族性高胆固醇血症〔familial hypercholesterolemia〕常染色体显性遗传病,患者编码LDL 受体的基因发生突变复习题1.构成细胞膜的脂类有哪三种?2.磷脂分为哪两种?3.哪一种磷脂在神经细胞含量多,其他细胞含量少?4.胆固醇分子对膜的流动性有何影响?5.动物细胞膜的糖脂由何磷脂衍生而来?6.膜功能的活跃与否跟什么成分的含量密切相关?7.根据与脂双层结合方式,膜蛋白可分为哪三类?8.内在膜蛋白的跨膜区,通常是哪类氨基酸残基构成的什么结构?9.外在膜蛋白通过什么键附着膜脂或膜蛋白?10.脂锚定蛋白在膜两侧以什么键结合于什么分子?11.膜糖链的唾液酸残基,在细胞外表面形成什么电荷?12.膜的不对称性主要体现在哪三点?13.膜脂分子能进行哪些运动?14.影响膜脂的流动性的因素有哪些?15.流动镶嵌模型主要内容是什么?16.脂筏模型的主要内容和特点各是什么?17.膜转运蛋白分为哪两类?18.哪些溶质能简单扩散到膜另一侧?19.被动扩散和主动运输主要区别是什么?20.离子通道的四个特点是什么?。

......
第三节物质跨膜运输的方式
一、被动运输
物质进出细胞,顺浓度梯度的扩散,称为被动运输。
(1)自由扩散:物质通过简单的扩散作用进出细胞
(2)协助扩散:进出细胞的物质借助载体蛋白的扩散
二、主动运输
从低浓度一侧运输到高浓度一侧,需要载体蛋白的协助,同时还需要消耗细胞内化学反应所释放的能量,这种方式叫做主动运输。
逆浓度梯度的运输
保证了活细胞能够按照生命活动的需要,主动选择吸收所需要的营养物质,排除代谢废物和有害物质。
方向载体能量举例
自由扩散高→低. 不需要不需要水、CO2、O2、N2、乙醇、甘油、苯、脂肪酸、维生素(水,气体小分子,脂溶性有机小分子,脂肪酸,胆固醇,性激素,维D)协助扩散高→低需要不需要葡萄糖进入红细胞
主动运输低→高需要需要氨基酸、K+、Na+、Ca+等离子、葡萄糖进入小肠上皮细胞三、大分子物质进出细胞的方式
胞吞、胞吐(如蛋白质,体现膜的流动性,需要消耗能量)
......。
特性流动性存在状态液晶态——既具有固态的有序性,又有液态的流动性形式★ 胆固醇的含量:虽可稳定相变温度,但多↓ ★ 脂肪酸链的长短和饱和程度:长↓,短↑★ 卵磷脂、鞘磷脂的比值:卵、鞘占膜脂的50% △卵磷脂:含不饱和脂肪酸程度高 ↑ △鞘磷脂:含 饱和 脂肪酸程度高 ↓ ★ 膜蛋白的含量(内在蛋白):类似胆固醇 影响意义★使膜具有缓冲作用,不易破裂 ★有利于内在蛋白作用发挥★有利于膜的正常分裂及吞噬、吞饮作用发挥不对称性◆ 外层:胆固醇、磷脂酰胆碱(PC)、鞘磷脂(SM)含量多。
①由于碳氢链长互相凝集,伸至全膜; ②三种成分亲合力强,影响流动。
◆ 内层:磷脂酰乙醇胺(PE)、磷脂酰丝氨酸(PS)、磷脂酰肌醇(PI)含量多。
上述三种成份头部基团带较强的负电荷,所以细胞内侧负电荷大于细胞外侧。
膜脂的不对称性膜蛋白不对称性◆糖蛋白、糖脂都分布在细胞膜外表面。
◆细胞内膜系统上的糖蛋白都位于膜腔内侧面。
膜糖类不对称性45%膜糖类2-5% 识别 稳定 保护成分膜 55%胆固醇:占膜脂1/3磷脂:占膜脂2/3糖脂:占2%左右磷脂酰胆碱 (卵磷脂PC ) 磷脂酰乙醇胺 (脑磷脂PE ) 磷脂酰丝氨酸 (PS ) 磷脂酰肌醇 (PI ) 鞘磷脂 (SM )糖蛋白:占膜糖类90%。
糖 脂:量少。
膜内在蛋白(整合、镶嵌、跨膜)脂锚定蛋白(脂连接蛋白) 占膜蛋白的70-80% 镶嵌于脂质双层中间 主要是跨膜蛋白占膜蛋白的20-30% 主要位于胞质面 细胞外表面很少 位于膜的两侧,与子分子结合 在细胞膜外表面共同构成―细胞外被‖ 或称―糖萼‖◆ 侧向扩散 ◆ 翻转运动◆ 旋转运动 ◆ 弯曲运动 ◆ 伸缩振荡细胞膜概念:包围在细胞质表面的一层薄膜。
又称质膜。
将细胞中生命物质与外界环境分隔开,维持细胞特有内环境。
功能膜 脂膜蛋白细胞膜的功能● “界膜”,对细胞起保护作用,为细胞提供生命活动的内环境 ● 内外物质交换和能量传递 ● 细胞识别与信息传递 ● 催化和调节生命代谢活动 ● 形成细胞表面特化结构 ◆ 极性亲水头部:磷酸、磷脂酰碱基(胆碱)非极性疏水尾部:两条非极性的、疏水的脂肪酸烃链◆ 双层排列:称―脂质双层‖(lipid bilayer )◆ 磷脂分子亲水头部都向膜的内外表面,疏水尾部向膜的中央 通常脂质双分子层又称为―双亲分子‖● 结 构 (以磷脂分子为例)◆ 构成生物膜的骨架◆ 膜的流动为膜的运动、分裂、物质交换提供了保证和便利 ◆ 膜脂的双亲性对进出细胞的物质起选择和屏障作用 ● 功 能◆ 特 点● 埋在脂质双层内的氨基酸都是疏水的。
生物细胞膜与物质运输知识点在我们的身体里,每一个细胞都像是一个小小的王国,而细胞膜就是这个王国的城墙和城门。
它可神奇了,不仅能把细胞内的世界保护得好好的,还能控制着各种物质的进出,就像一个超级严格但又聪明的海关管理员。
细胞膜主要是由磷脂双分子层构成的,这就像是两层紧密排列的小卫兵。
磷脂分子有一个亲水的脑袋和两条疏水的尾巴,所以它们会自动排列成两层,亲水的脑袋朝外,疏水的尾巴朝里,这样就形成了一个稳定的结构。
在这两层磷脂分子中间,还镶嵌着各种各样的蛋白质分子,有的像是站岗的哨兵,有的像是负责运输的货车司机。
细胞膜控制物质进出的方式有好几种,咱先说自由扩散。
这就好比是一个没有门卫的大门,一些小分子物质,比如氧气、二氧化碳,它们自己就能轻轻松松地进出细胞,完全不受阻拦,想进就进,想出就出,自由得很呐!再来说说协助扩散,这有点像有门卫稍微看一眼的门。
一些稍微大一点的分子,比如葡萄糖,它们自己过不去细胞膜这道关卡,得靠细胞膜上的一些特殊通道蛋白帮忙,才能顺顺利利地进出细胞。
这些通道蛋白就像是专门为它们开的绿色通道,只要它们符合条件,就能快速通过。
最厉害的要数主动运输啦!这就像是有一群力大无穷的搬运工在努力工作。
细胞需要从外界摄取一些对自己很重要,但浓度又比较低的物质,比如钠离子、钾离子,还有各种氨基酸、葡萄糖等。
这时候,细胞膜上的载体蛋白就得大显身手了。
它们会消耗细胞内的能量,也就是 ATP,然后像大力士一样,把这些物质从低浓度的一侧“搬”到高浓度的一侧。
这可不容易,就像是逆水行舟,需要付出很大的努力。
但细胞为了生存和正常工作,必须要这么做。
我还记得有一次上生物课,老师为了让我们更清楚地了解细胞膜的物质运输,给我们做了一个特别有趣的实验。
老师拿来了几个鸡蛋,我们都特别好奇,这鸡蛋和细胞膜有啥关系啊?只见老师小心翼翼地把鸡蛋的外壳去掉,露出了里面白白嫩嫩的那层膜。
老师说,这层膜就类似于细胞膜。
然后,老师把一个装着蓝色墨水的小杯子放在鸡蛋膜下面,过了一会儿,我们惊奇地发现,蓝色的墨水慢慢渗透进了鸡蛋膜里。
第四章细胞膜与物质的穿膜运输第一节细胞膜的化学组成与生物特性一、细胞膜的化学组成细胞膜上的脂类=膜脂(membrane lipid),约占膜成分的50%,主要有磷脂(phospholipid)、胆固醇(cholesterol)、和糖脂(glycolipid)(一)膜脂构成细胞膜的结构骨架1.磷脂是膜脂的主要成分甘油磷酸的共同特征:以甘油为骨架,甘油分子的1、2位羟基分别于脂肪酸形成酯键,3位羟基与磷酸基团形成酯键。
磷酸基团结合胆碱/乙醇胺/丝氨酸/肌醇。
脂肪酸链长短不一,通常14~24个碳原子,一条脂肪酸链不含双键,另一条含有一个或几个双键,形成30°弯曲。
鞘磷脂以鞘氨醇代替甘油,鞘氨醇的氨基结合长链的不饱和脂肪酸,分子末端的一个羟基与胆碱磷酸结合,另一个游离羟基可与相邻分子的极性头部、水分子或膜蛋白形成氢键。
鞘磷脂及其代谢产物神经酰胺、鞘氨醇、1-磷酸鞘氨醇参与各种细胞活动。
神经酰胺是第二信使;1-磷酸鞘氨醇在细胞外通过G蛋白偶联受体起作用,在细胞内与靶蛋白作用2.胆固醇能够稳定细胞膜和调节膜的流动性✧胆固醇为两性极性分子。
✧极性头部为连接于固醇环(甾环)上的羟基,靠近相邻的磷脂分子。
✧固醇环疏水,富有刚性,固定在磷脂分子临近头部的烃链上,对林芝的脂肪酸尾部的运动具有干扰作用。
✧尾部为疏水性烃链。
埋在磷脂的疏水尾部中。
✧胆固醇分子调节膜的流动性和加强膜的稳定性。
没有胆固醇,细胞膜会解体。
PS.不同生物膜有各自特殊的脂类组成。
哺乳动物细胞膜上富含胆固醇和糖脂,线粒体膜内富含心磷脂;大肠杆菌质膜则不含胆固醇。
3.糖脂主要位于质膜的非胞质面糖脂含量占膜脂总量5%以下,遍布原核、真核细胞表面细菌和植物的糖脂均是甘油磷脂衍生物,一般是磷脂酰胆碱PC 衍生来动物糖脂都是鞘氨醇衍生物,称为鞘糖脂,糖基取代磷脂酰胆碱,成为极性头部已发现40多种糖脂,区别在于极性头部不同,由1至几个糖残基构成✧最简单的糖脂是脑苷脂,极性头部只是一个半乳糖/葡萄糖残基✧最复杂的糖脂是神经节苷脂,极性头部有七个糖残基;在神经细胞膜中最丰富,占总膜脂5%~10%✧脂质体(lipidsome)可以作运载体(二)膜蛋白以多种方式与脂双分子层结合又称含量作用力特点膜内在蛋白穿膜蛋白70%~80% 范德华力α-螺旋构象/β-筒孔蛋白膜外在蛋白外周蛋白20%~30% 非共价键水溶性脂锚定蛋白脂连接的蛋白共价键运动性增大1.内在膜蛋白✧又称跨膜蛋白,占膜蛋白总量70%~80%;分单次跨膜、多次跨膜、多亚基跨膜三种类型✧跨膜区域20~30个疏水氨基酸残基,通常N端在细胞外侧✧内在膜蛋白跨膜结构域与膜脂结合区域,作用方式:①疏水氨基酸形成α-螺旋,跨膜并与脂双层脂肪酸链通过范德华力相互作用②某些α-螺旋外侧非极性,内侧是极性链,形成特异性畸形分子的跨膜通道✧多数跨膜区域是α-螺旋,也有以β-折叠片多次穿膜形成筒状结构,称β-筒,如孔蛋白(porin)2.外在膜蛋白又称外周蛋白,占膜蛋白总量20%~30%;完全在脂双层之外,胞质侧或胞外侧,通过非共价键附着膜脂或膜蛋白胞质侧的外周蛋白形成纤维网络,为膜提供机械支持,也连接整合蛋白,如红细胞的血影蛋白和锚蛋白外周蛋白为水溶性蛋白,与膜结合较弱,改变溶液离子浓度或pH,可分离它们而不破坏膜结构3.脂锚定蛋白①一种位于膜的两侧,蛋白质直接以共价键结合于脂类分子;此种锚定方式与细胞恶变有关②还有糖基磷脂酰肌醇锚定蛋白(GPI),通过蛋白质C端与磷脂酰肌醇连接的糖链共价结合脂锚定蛋白在膜上运动性增大(侧向运动),有利于结合更多蛋白,有利于更快地与胞外蛋白结合、反应GPI-锚定蛋白分布极广,100种以上,如多种水解酶、免疫球蛋白、细胞黏附分子、膜受体等4.去垢剂(detergent)离子型去垢剂:SDS十二烷基磺酸钠引起蛋白质变性非离子型去垢剂:Triton X-100 对蛋白质比较温和(三)膜糖类覆盖细胞膜表面细胞膜的糖类,占质膜重量2%~10%;①大多以低聚糖或多聚糖共价结合膜蛋白,形成糖蛋白(糖蛋白中的糖基化主要发生在天冬酰胺(N-连接),其次是丝氨酸和苏氨酸(O-连接)残基上);②或以低聚糖共价结合膜脂,形成糖脂,所有糖链朝向细胞外表面形成低聚糖的单糖类型:甘露糖、岩藻糖、半乳糖、半乳糖胺、葡萄糖、葡萄糖胺、唾液酸等A.唾液酸残基在糖链末端,形成细胞外表面净负电荷B.寡糖链中的单糖的数量、种类、排列顺序、有无支链等不同,可以出现千变万化的组合形式。
Eg.人类ABO血腥抗原的差别就是血型糖蛋白在红细胞质膜外表面寡糖链的组成结构决定。
△细胞外被cell coat=糖萼glycocalyx=与质膜相连的糖类物质功能:①保护细胞抵御各种物理、化学性损伤②建立起水盐平衡③帮助蛋白质膜上定位、固定,防止翻转④参与细胞与外环境的作用,eg识别、粘附、迁移二、细胞膜的生物学特性(一)膜的不对称性决定膜功能的方向性膜结构上的不对称性保证了膜功能的方向性和生命活动的高度有序性1.膜脂的不对称性SM、PC在细胞外侧叫多,PE在细胞内侧较多。
2.膜蛋白的不对称性各种膜蛋白在质膜中有特定位置,分布绝对不对称:酶和受体多分布于质膜的外侧面,而腺苷酸环化酶定位内侧面跨膜蛋白有一定方向性:多数N外C内,两端肽链长度、氨基酸种类、活性位点不同3.膜糖的不对称性都向着非胞质面(二)膜的流动性是膜功能活动的保证流动性fluidity主要是指膜脂的流动性+膜蛋白的运动性1.脂双层为液晶态二维流体✓液晶态(lipid-crystal state)脂双分子层已有固体分子排列的有序性,又有液体的流动性。
细胞内外的水环境,使膜脂分子不能从脂双层逸出,只能在二维平面交互位置。
✓相变(phase transition)正常体温下,膜呈液晶态;当温度下降到临界温度(膜的相变温度),膜脂转为晶态✓膜的流动性是膜功能活动的保证。
2.膜脂分子的运动方式①侧向扩散lateral diffusion =脂双层的单分子层内,脂分子沿膜平面侧向与相邻分子快速交换位置,每秒约107次。
侧向扩散运动时膜脂分子主要的运动方式。
②翻转运动flip-flop 从脂双层一层翻转到另一层,需要翻转酶,在内质网发生③旋转运动rotation 膜脂分子围绕与膜平面向垂直的轴的自旋运动④弯曲运动flexion 膜脂分子的烃链是有韧性、可弯曲的,分子尾部端弯曲、摆动幅度大,而靠近头部弯曲摆动幅度小。
⑤此外,还有伸缩、震荡3.影响膜脂流动性的因素①脂肪酸链的饱和程度磷脂分子长的饱和脂肪酸链呈直线型,具有最大的聚集倾向而排列紧密成凝胶状态;不饱和脂肪酸链在双键出形成折曲而呈弯曲状,感染了脂分子间范德华力的相互作用,故排列疏松,从而增加了膜的流动性。
∴脂双分子层中含有的不饱和脂肪酸越多,膜的相变温度越低,流动性越大。
环境温度降低时,A.细胞通过去饱和酶(desaturases)催化将胆碱去饱和形成双键。
B.通过磷脂酶&脂酰转移酶在不同的磷脂分子之间重组脂肪酸链以产生含两个不饱和脂肪酸链的磷脂分子。
②脂肪酸链的长短脂肪酸链短的相变温度低,流动性大。
短→尾端不易发生相互作用;长→不仅可以在同一分子称内相互作用,而且可以与另一分子层中的长链尾端相互作用③胆固醇的双重调节作用A.当温度在相变温度以上时,由于胆固醇分子的固醇环与磷脂分子靠近极性头部的烃链部分结合,限制了这几个CH2的运动,起到稳定质膜的作用。
B.当温度在相变温度以下时,由于胆固醇位于磷脂分子之间隔开磷脂分子,可有效地防止脂肪酸链相互凝聚,干扰晶态的形成。
④卵磷脂与鞘磷脂的比值哺乳动物细胞中,卵磷脂和鞘磷脂的含量约占膜脂的50%,卵磷脂的脂肪酸链不饱和程度高,相变温度较低;鞘磷脂则相反。
在细胞衰老过程中,卵磷脂和鞘磷脂的比值下降,流动性也下降。
⑤膜蛋白的影响膜蛋白嵌入膜脂疏水区后,是周围的脂类分子不能单独活动而形成界面脂;在含较多内在蛋白的膜中,存在有内在蛋白分割包围的富脂区(lipid-rich region)磷脂分子智能在一个富脂区内自有扩散,而不能扩散到邻近的富脂区此外,膜脂的极性基团、环境温度、pH值、离子强度等都对膜脂流动性产生一定影响。
环境温度高,膜脂流动性大;相变温度内,每下降10℃,膜的粘性增加3倍,膜流动性降低4.膜蛋白的运动性①侧向扩散膜蛋白在膜脂中可以自有漂浮&在膜表面扩散。
人鼠杂交细胞表面抗原分布变化可证明。
目前测定膜蛋白的侧向扩散常采用光致漂白荧光恢复法(fluorescence recovery after photobleaching,FRAP)②旋转运动膜蛋白能围绕与膜平面相垂直的轴进行旋转运动。
速度比侧向扩散慢;不同膜蛋白速度不同,有些膜蛋白无法运动;膜蛋白周围脂质的流动性影响膜蛋白的流动性膜蛋白的运动不需要消耗能量膜的流动性意义重大:物质运输、细胞识别、信息传导等;生物膜的各种功能都是在膜的流动状态下进行的,膜的流动过低,代谢终止三、细胞膜的分子结构模型(一)片层结构模型具有三层夹板式结构特点1935年,James Danielli 和Hugh Davson发现细胞膜的表面张力显著低于油-水界面表面张力,推测质膜中有蛋白质;提出“片层结构模型”(蛋白-磷脂-蛋白三层夹板式结构) (二)单位膜模型体现膜形态结构的共同特点1959年,J.D.Robertson 电镜观察细胞膜“两暗夹一明”——单位膜单位膜模型:膜蛋白是单层肽链以β折叠通过静电作用与磷脂极性端结合;能对膜的某些属性进行解释,被普遍采用,但是把膜作为静止的单一结构(三)流动镶嵌模型是被普遍接受的模型1972年,“流动镶嵌模型”Fluid mosaic model:磷脂双层构成膜的连续主体,具有晶体的有序性和液体的流动性;球形蛋白质分子以不同形式结合脂双层分子;膜是一种动态的、不对称的具有流动性结构1975年,“晶格镶嵌模型”:膜脂可逆地进行“有序(液态)”和“无序(晶态)”相变,膜蛋白对膜脂的运动具有限制作用,流动性是局部的1977年,“板块镶嵌模型”:流动的脂双层中存在能独立移动脂类板块(四)脂筏模型深化了对膜结构和功能的认识✓脂双层中由特殊脂质和蛋白质组成的微区,富含胆固醇和鞘脂类,聚集特定种类膜蛋白;此膜区较厚(鞘脂类脂肪酸链较长),称“脂筏”Lipid rafts,其周围富含不饱和磷脂,流动性较高✓脂筏的两个特点:许多蛋白聚集在脂筏内,便于相互作用;脂筏提供有利于蛋白质变构的环境,形成有效构象✓脂筏功能:参与信号转导、受体介导内吞作用、胆固醇代谢运输等第二节小分子物质和离子的穿膜运输一、膜的选择性通透和简单扩散简单扩散(simple diffusion): 小分子的热运动使分子以自由扩散的方式由膜一侧扩散到另一侧,条件:溶质在膜两侧有一定浓度差,溶质必须能透过膜脂溶性物质如醇、苯、甾类激素、O2、CO2、NO、H2O 通过简单扩散跨膜简单扩散不需要运输蛋白协助,顺浓度梯度由高浓度向低浓度方向扩散,不消耗能量;也称“被动扩散”passive diffusion二、膜运输蛋白介导的穿膜运输除了水和非极性小分子,绝大多数溶质如各种离子、葡萄糖、氨基酸、核苷酸等都不能简单扩散穿膜转运特定膜蛋白——膜运输蛋白(跨膜蛋白,每种只转运一种特定类型溶质)膜运输蛋白分两类:①载体蛋白carrier protein:与特定溶质结合,改变构象使溶质穿越细胞膜②通道蛋白channel protein:形成水溶性通道,贯穿脂双层,通道开放时,特定溶质(无机离子)可穿越脂双层"被动运输" passive transport所有通道蛋白和许多载体蛋白,转运溶质分子不消耗能量,消耗顺电化学浓度梯度的势能“主动运输”active transport逆电化学浓度梯度转运溶质,需要载体蛋白参与,还需要消耗能量A TP;这种利用代谢产生能量的进行逆浓度梯度的转运称为主动运输。